










Services on Demand

article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkCurrent Ethology
On-line version ISSN 2594-9985
Curr. Ethol. vol.15 no.2 Ihléus July/dez. 2016
Social behavior adaptations in agroup of black-and-gold howler monkeys (Alouattta caraya; Primates, Atelidae) inside an urban forest in the southeast of Brazil
Adaptações do comportamento social em um grupo de macacos bugios pretos e dourados (Alouatta caraya; Primatas, Atelidae) dentro de uma floresta urbana no sudeste do Brasil.
Ricardo SampaioI, *; Hugo Lopes GomesII; Zelinda Maria Braga-hiranoIII; Juliana Moutinho PedrosaIV; Wagner Ferreira Dos SantosII, IV, V
IPostgraduate Program in Comparative Biology (FFCLRP-USP), Ribeirão Preto, SP, Brazil
IIPostgraduate Program in Psychobiology (FFCLRP-USP), Ribeirão Preto, SP, Brazil
IIIRegional University of Blumenau (FURB), Blumenau, SC, Brazil
IVBiology Department, Collegeof Philosophy, Sciences and Literature of Ribeirão Preto - University of São Paulo (FFCLRP-USP); Barba-Negra Project,Ribeirão Preto, SP, Brazil
VBehavioral Neurosciences Institute (INeC), Ribeirão Preto, São Paulo, Brazil
ABSTRACT
Primates of the Alouatta genus arefolivore - frugivore, resulting in a strategy of reduced energy expenditure, and the spatial relationships between these animals may constitutean important indication of how they establish their social organization. This study aimed to evaluatethe social behaviorof a group of black-and-gold howler monkeys (Alouata caraya) in an urban forest (3 ha) located in Ribeirão Preto (21º10'17" S and 47º48'05" W; São Paulo, Brazil). Thisgroup was organized in a harem systemthat faced instability,although during the study an age-graded system seems to have been stablished in the group. Amatrifocal subgroup was located in the center of the group and a male coalition was located in its periphery. However, this setting changed over the year. Few agonistic episodes were observed (2),and the inter-individual competition seemed reduced due to the high availability of fruit. This study providesimportant information about the plasticity in the social behavior of black-and-gold howler monkeys under the influence of anthropic actionand isolation of habitat.
Keywords: Black howler monkey, urban forest, Alouatta caraya, social interaction.
RESUMO
Os primatas do gênero Alouatta são descritos como folívoros - frugívoros, o que pode levar a uma estratégia de redução do gasto energético e o relacionamento espacial entre indivíduos pode constituir uma importante indicação de como um grupo estabelece sua organização social. Nosso estudo avaliouo comportamento social de um grupo de bugios pretos e dourados (Alouata caraya) em uma floresta urbana (3 ha) em Ribeirão Preto (21º10'17 "S e 47º48'05" W; São Paulo, Brasil). Este grupo se organizou em um sistema de harém, enfrentando instabilidade, porém ao longo do estudo um sistema por ordem de idade parece ter sido estabelecido no grupo. Um subgrupo matrifocal estava localizado no centro do grupo e uma coalizão de machos na periferia, mas essa configuração mudou ao longo do ano. Vimos somente poucos episódios agonísticos (2) e a competição interindividual pareceu diminuída devido à alta disponibilidade de frutos. Este estudo fornece importante informações sobre a plasticidade no comportamento social de bugios vivendo sob influência da ação antrópica e isolamento do hábitat.
Palavras chaves: Bugio preto, floresta urbana, Alouatta caraya, interação social.
INTRODUCTION
Howler monkeys (Alouatta) are considered to have a strategy to reduce energy expenditure (Crokett & Eisenberg, 1987; Milton, 1999, 2000; Strier, 1992).This pattern behavior is probably related to a more folivorousthan frugivorous diet (Garber 1987, Neville et al., 1988, Pavelka & Knopff, 2004), in which amechanism of substance detoxificationto extract energy from leavesleads to an inactive behavior (Glander, 1975).
In the social system ofhowler monkeys,inter-individual aggression israre (Altmann, 1959; Carpenter, 1934; Southwick, 1963),and theyorganize their communication and social behavior by means of indirect signals (approaching and retreating), with unusual episodes of bouts and vocalizations (Jones, 1982; Jones, 1983; Wang& Milton, 2002). Researchers believe that these signals, as well as grooming, are more related to the maintenance of a group-hierarchic social structure than to agonistic behaviors (Jones, 1982). Therefore, the spatial relationship between individuals in a howler monkey group can be an important indication of how they relate and organize their hierarchic social structure (Altmann, 1959; Jones, 1980; Wang & Milton, 2003; Bezanzon et al., 2002 and 2008).
The social groups of howler monkeys are organized in many patterns, from a typical harem - where a dominant male restricts the access to females -to large groups with several males, which shows a hierarchical social structure and differentiated access to females (Eisenberg et al., 1972; Neville et al., 1988), the latter being described for the species A. palliata,A. pigra, A. seniculus,and A. caraya (Crockett & Eisenberg, 1987; Rumiz, 1990; Kowalewski, 2000). However, some individuals can form subgroups as facultative responsesto different social and ecological conditions (Kinzey & Cunningham, 1994; Kappeler & van Schaik, 2002).
Based on indirect signs expressed byinter-individual distance between all individuals, we analyzed, during the course of a year, the social behavior of a black-and-goldhowler monkey group (A. caraya) living in an isolated urban fragmented forest of 25 hectares inside of thecity of Ribeirão Preto(São Paulo, Brazil). We expected this groupto befacing high anthropogenic impact living inside this isolated forest fragment, since the intra-individual competition for resources can be high, and subordinated individuals could not immigrate to other areas,resulting in instability in the social structure of the group.
Considering this information, we have three predictions about their social structure: 1st) The group presentsa multi-male system with social and hierarchic structure;2nd) There are signs of instability and of formation of subgroups; and 3rd) The group shows a weak pacific trend in its social behavior.
In order to corroborate these predictions, we will answer the following questions: 1) Do the adult males have different access to the adult females?; 2) Is there a formation of subgroups, and how arethey built?;and 3) Is there too much aggression in the group, and which individuals are the most involved?
METHODS
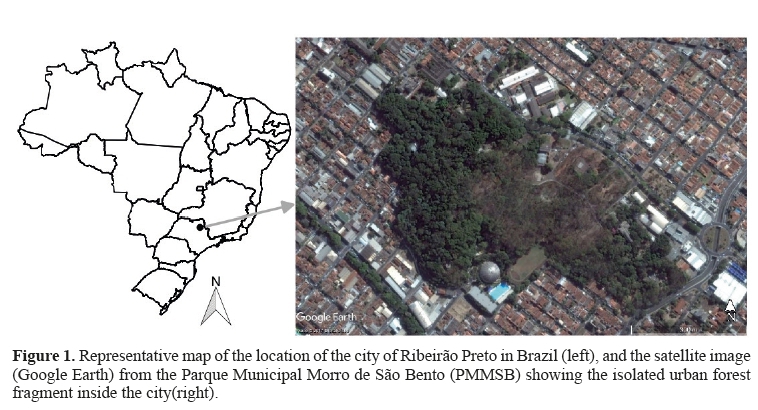
This study was performed at Parque Municipal Morro de São Bento (PMMSB) -21º10´17´´ S, 47º48´05´´ W, which is located in the city of Ribeirão Preto (São Paulo, Brazil). The PMMSB is a park (25 ha, Figure 1) that features the City Zoo, as well as being a semi-deciduous and deciduous mesophyllous forest fragment of approximately 3 ha (Laguna, 1997). This area contains native and exotic plant species and wasformally homologatedas an Environmental Protection Area (Área de Proteção Ambiental; APA).
A four-month period (Jun-Oct, 2000) was necessary for the animals' habituation with the researchers. During the same period, the group was censured and only one group of 17 individuals was confirmed. The animals were grouped using age and gender categories, according to Calegaro-Marques and Bicca-Marques (1993): Adult Males (M); Adult Females (F); SubadultMales (SAM); Juveniles (J); Infants (I). The subadult female category was not defined, for it was difficult to distinguish it category from juveniles.
All membership was separated inthe following classes: M, F, SAM, J, and I;as well as receiving codes (e.g.: M1-2, SAM1-4, F1-5). It was not possible to distinguish individuals within the juvenile category, but they were included in the analysis and were classified in order of appearance in each sampling period (J1, J2, J3...)
The observations were made during four days in each month,from sunrise to sunset(betweendawn and dusk), from Nov/2000 to Oct/2001. We used only two categories of inter-individual distance to interpret the hierarchic social structure and the formation of subgroups: A, two individuals in body contact; and B, 0 to 3 meters between two individuals. Inter-individual distances greater than 3 meters were recorded, but were not used in our analyses.
The ad libitum method (Altman, 1974), was used to analyze our predictions #1 and 3, in which any social interactionsbetween the individuals, like conflicts, agonistic behavior and sexual behavior was sampled during all the study period.The scan sampling method was employed (Altman, 1974)to analyze our prediction #2, in which the spatial relationship between all the members of the group was sampled in a 20-minute intervals.
The study was divided into four social dynamic phases (Table 1), varying in accordance with the composition of the group, which directly affected the social dynamics and interactions between the individuals in our field of vision.
Theinter-individual distance data were analyzed with multivariate analysis (Cluster Analysis). This consists in the formation of groups of individuals according to the similarities of variables displayed bythem. The analyses included two steps: 1) establishment of similarity index (S.I.) matrix, grade of similarity or association between counts inter-individual distances; 2) formation of groups by a linking method, and construction of dendograms based on these indexes (Hair et al., 1995).
In this study, the chosen S.I. was the total number of episodes of distances A or B, which produced similarity matrices for each distance. These were imported to the PRIMER 5 software (Clarke & Warwick, 2001), which grouped the individuals based on the similarities, expressed by percentages of similarity, of all S.I., building dendogramsvia a linking method, in which the average of the groups was used.
Using the dendograms, we built diagrams that represent thelevel of association between all individuals. The intensity of this association is represented by arrows that are proportional to the percentage of similarity showed in the dendograms.
RESULTS
The number of individuals in the group during the study period is shownin Table I. The number of individuals varied from seventeen (Nov/2000) to twenty-two howlers(Oct/2001), andonly two episodes of agonistic behavior throughout the entire study period were recorded, whereas both were performedonly by adult females (Sampaio, 2002).
In the first phase, only one group living in this areawas identified. The second phasewas characterized by the integration1 of three individuals (F5, I3, and J6) - for unknown reasons - in the group, animals which probably lived in the peripheral area and wentunnoticed during the first phase. The third phase was characterized by the birth of a howler (I4). In the fourth phase, another howler (I5) was born.
First Phase
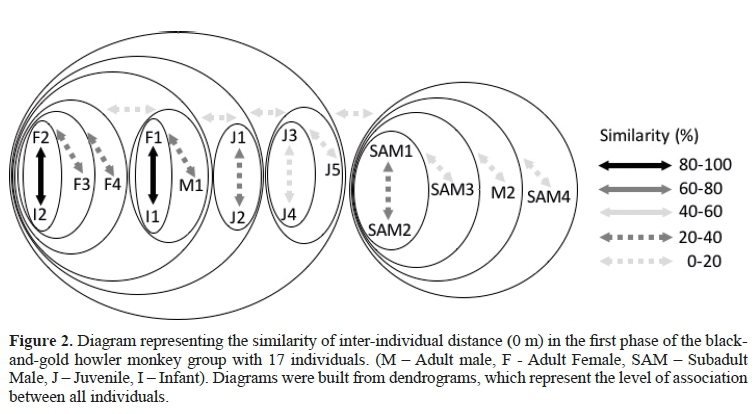
As shown in Figure 2 (A = 0 m), two subgroups of individuals were observed, whereas the first one contained all of the adult females that characterized strong affinities with their infants (matrifocal subgroup). The male adult 1 (M1) was located close to them. The juveniles of the group established an association (0-40%) between themselves, as well aswith the adult females (0-20%). The second subgroup showed associations between subadult males and the M2 (0-40%), with little affinity towardsthe adult females (0-20%).
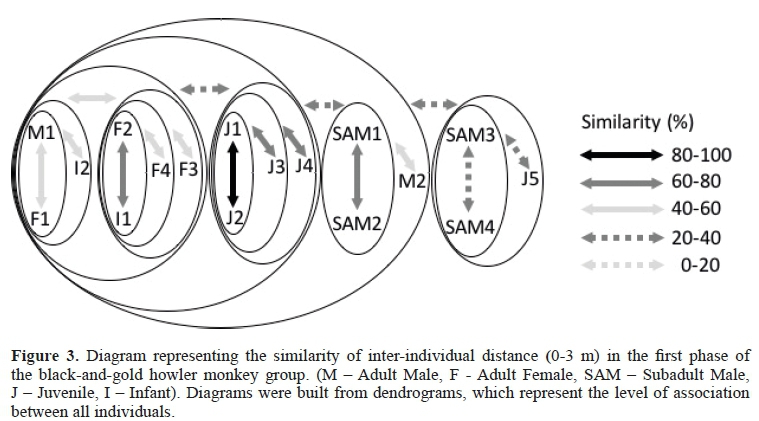
When the distance considered between individuals increased (B= 0 to 3 m), as shown in Figure 3, the adult females still showed high association with their infants (40 - 80%),with the M1 showing proximity to them (40-60%). The juveniles showed greater association between themselves(40-100%)and with the adult females (20-40%). The M2, SAM1, and SAM2 were more associated with subgroup 1, appearing in the periphery of the group. The SAM3 and 4 were associated (20-40%) together with a 5juvenile, and presented little affinity in relation to the rest of the group (0-20%).
Second Phase
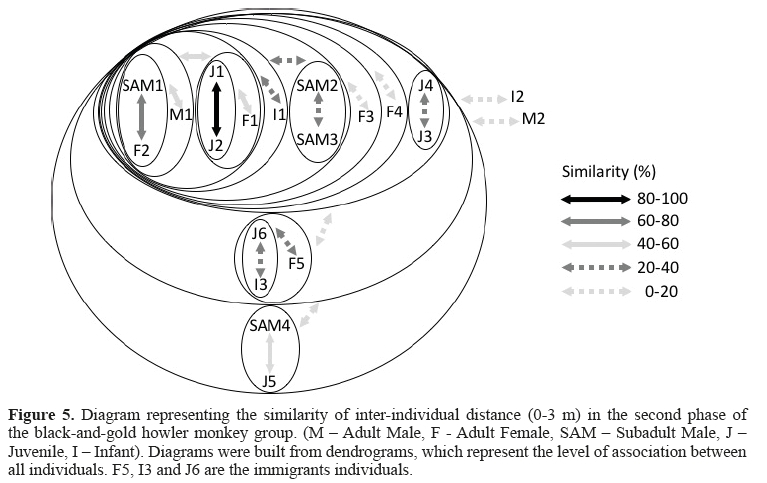
The second phase was characterized by the integration of three new individuals in the group (F5, J6, and I3). These immigrants remained associated (20 - 60%) and isolated from the rest of the group, as shown in Figure 4 (A= 0 m).
In general, the level of closer proximity between the individuals also decreased (Figure 4). The affinity among the adult females (matrifocal subgroup) became less pronounced than in the first phase, and the F4 distanced itself from the matrifocal subgroup, as well as being in association with the J4 (40-60%). The M1 was associated with lower affinitytowards two juveniles (J1 and J2) and with the matrifocal subgroup (0-20%). The SAM1, 2, and 3 were associated (0-40%) and isolated from the main group. The M2 was associated with the J5. Meanwhile, the SAM4 was isolated from the group. The I2, which is presumablyF2's son, showed a smaller proximity to F2in this phase than in the first one.
When analyzing Figure 5 (B= 0 - 3 m),it is not possible toobserve the formation of subgroups. The majority of the individuals interacted with low percentages of affinity (20%). The M1 and SAM1 were in the center of the group, while the SAM2 and 3 were associated andcloser to this centerwhen compared to the previous phase, and the SAM4 and M2 were at the edge of the group. The F1 and 2 were still in the center of the group, and the F3 and 4 were located far from it. The juveniles were still associatedbetween themselves and with other classes.The immigrants still maintained a tight connection, and M2, SAM4, J5, and I2 were positioned in the periphery of the group.
Third Phase
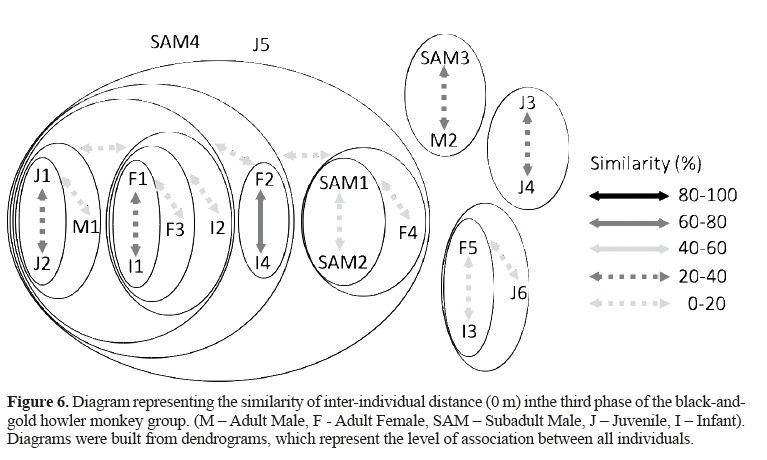
This phase began when the F2 gave birth to the I4 and its previous infant (I2) started to show a juvenile behavior, which could be noticedin thefield and is shown in Figure 6 and 7.However, due to its smaller size,it could be distinguished from the other juveniles andwe still decided to recognize it as an infant.
In Figure 6 (A = 0 m), it is possible to note that the adult females 1, 2, and 3 were in the center of the group, and that twojuveniles and I1 and I2 were still there, too.The M1 was found near them and,with the exception of the interaction between F2-I4, the similarity between them was lower than 40%. The SAM1 and 2 were weakly connected between themselves, with the F4,and with the matrifocal subgroup (0-20%).
The SAM3 and the M2 were associated in the periphery of the group. Two juveniles (J3 and J4)were associated and isolated from the whole group, while another one was found isolated from the group. The immigrants (F5, I3, and J6) were still interacting and separated from the rest of the group.
In Figure 7 (B = 0 - 3 m), it is possible to observe the formation of a single group that displayed a low level of association between the individuals. The adult females were not in the center of this group, whereas this position was occupied by three juveniles (J1, J2, and J3) accompanied by theF1, F2, F3, and F5. The M1 and SAM1 were associated near them. The SAM2 and 3 and the M2 were more peripheral, but associated with of the group.
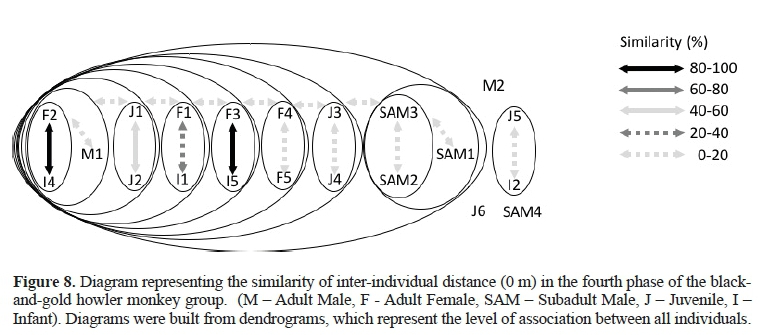
Fourth Phase
This phase began when the F3 gave birth to the I5.Figure 8 (A = 0 m) shows the matrifocal grouping includingall adult females.The M1 and two juveniles (J1 e J2) were found very close to them. Subadult males 1, 2, and 3 were in association between themselves and at the edge of the group, while the SAM4 and M2 were isolated from it. The infant I2 showed affinity with the J5 in the periphery of the group. The strongest affinities (80-100% of similarity) were characterized in the association between the F2 and the F3 with their infants I4 and I5, respectively.
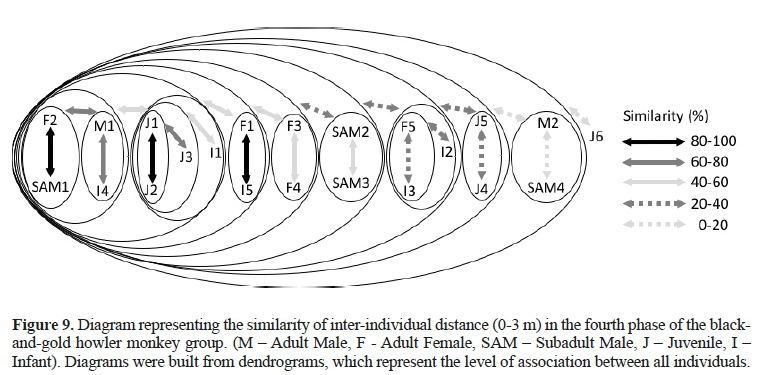
As shownin Figure 9 (B = 0 - 3 m),in the center of the group, it is possible to observe that F2, SAM1, M1, three juveniles,two infants, and the other females (F1, F3, F4, and F5) were displaced from near the individuals - as mentioned above- to interact withSAM2, SAM3, I3, and I4. Therefore, the matrifocal subgroup cannot be seen in Figure 9. The M2, SAM4, and J6were found in the periphery of the group.
DISCUSSION
The number of individuals in the group studied (17-22) was relatively higher than the average for the Alouatta caraya species(Rumiz, 1990; Bicca-Marques, 1991, Kowalewski &Zunino,1999; Dvoskinet al., 2004, Juárez et al., 2005), and for the Alouatta genus, since the average number of individuals usually varies from 3 to 21 animals (Carpenter, 1934; Crockett & Eisenberg, 1987; Miranda & Passos, 2005;Estrada et al., 2002; Flórez& Delgado, 2011, Azkarate et al., 2017). These 22 individuals were confined in a 3-ha semi-deciduous forest fragment (see Laguna, 1997), which comprisesa highpopulation density of 100/km2, similar only in the number found in forests of the Argentina (Brown & Zunino, 1994).
Besides the isolation of this fragmented forest, this large number of individuals living in a small fragmented area may be explained by the high percentage of fruit ingested by the howler black-and-gold monkeys when compared to other groups,as described by Pedrosa (2002) during the same periodstudied. The area contains several exotic botanical species, including Caryota urens (Laguna, 1997 and 2000), a palm tree whose fruit supply lasts the entire year (Gomes, 2004). In the period of the year in which the variety and availability of the fruits are lower, the howler monkeys use the food offered to the animals of the Zoo (Pedrosa, 2002; Gomes, 2004).
The confined system this group faced -in whichthe migration of individuals or subgroups to other associated forest areas is impossibledue to the isolation of the area (see Figure 1)together with the great availability of food resulted in a high population density. This context led us to predict that the group could present: (1) a multi-male system with social and hierarchic structure; (2) signs of instability and formation of subgroups; and (3) weak pacific trends in their social behavior.
During the entire sampling period,the group had two adult males, where the M1 was interpreted as being the first in the hierarchy, since it always located near the adult females, and M2 was always in the periphery of the group, far from the adult females, leading us to interpret that this group was socially organized in a harem system or in an age-graded system, thus not confirming our first hypothesis.
The possible coalition among the M2 and all SAMs took place in the periphery of the group, with few interactions with the adult females. The juveniles were frequently associated between themselves (mainly during playtime), with adult females (looking for protection and social interactions, probably with their mothers or with other relatives), and with SAMs. The only two copulations recorded by us (Sampaio, 2002) were performed by the M1, and we did not observe any sexual interaction between the other males and adult females.
The social organization in the genus Alouatta seems to be variable. Groups of theA. palliataspecies exhibits multi-male organization where the interactions between adult males are meant to assure their access to the adult females, and the inter-individual distances between males and females indicate their reproductive success (Jones, 1982).Neville (1972) showed that theA. seniculusspecies can organize their groups throughanage-graded or harem system. However, the behavior of the alpha male was not well characterized.
The social organization pattern of our study group (harem or age-graded system)is not common for the Alouattacarayaspecies (see Rumiz, 1990 and Kowalewski, 2000), and the harem systems in the Alouattaspecies was organized through a matrifocal nucleus and a dominant adult male that had no need for proximity to the females, but had access to them during the whole time (Neville et al., 1988). Oliveira and Ades (1998) investigated several groups ofA. fuscaspecies (= A. g. clamitans) and observed that adult males were frequently isolated, but that they could sometimes be grouped or associated with adult females.
Concerning our second hypothesis, the formation of subgroups and the occurrence of isolated animals are often considered signs of instability in groups of howler monkeys. Neville (1972) was the first author to report the formation of subgroups in a 16-individual group of A. seniculus.
In our study was observed signs of instability in all sample periods, thus confirming our second hypothesis and suggesting the occurrence of the formation of two separate subgroups - one matrifocal subgroup and a coalition of subordinated males -, and these dynamics changed throughout the sampling, wherein the first subgroup consisted in the matrifocal subgroup with an adult female and young animals. With the exception of the adult male, all the members of another subgroup - formed by peripheral males and juveniles- showed interaction with the matrifocal group.
Bezanson et al. (2002)stated that individuals of different sex-age classes ingroups of Primates may show different patterns of subgroup formation according to the context they belong to. This concurs with what we observed in the adult and subadult males, adult females, and infants. Nevertheless, the interactions and the social organization within this group changed throughout the year.
In the first phase, the matrifocal subgroup was characterized by the association between all adult females and their infants, and the M1associated with them can be understood as a repelling behavior of the other males in order to avoid infanticide and allowaccess to copulation. This male behavior was only observed by Jones (1983) inregardingA. seniculus species.
In the second phase, the immigrant howler monkeys increased the instability within the group, and the affinity of the matrifocal subgroup was weaker than in the first phase. This may have happened because the immigrant adult female (F5) and its infant (I3) may have had a higher interest on the young females of the group, having directed their "new" attention towards an infant, causing a lower association of the F3 and a separation of the F4 from the matrifocal group.The reason behind this may be the possible attempt to form another subgroup.
The M1 remained close to the matrifocal subgroup, but the SAM1 exhibited low association with the other subadult males and with the M2,whereas it showed more association withthe adult females and with the M1. Subadults males 2 and 3 interacted and were inserted in the group, while SAM4 and M2 showed more peripheral positioning within the group.
In the third phase, we believe that the birth of theI4 may have contributed to the return of F3 and F4to the subgroup of adult females (matrifocal subgroup), even though the affinity was lower than that observed in the first phase. This finding was similar to the one observed by Neville et al. (1988), who verified that the birth of an infant of howler monkeys increases the affinities of adult females among themselves, as well as with their youth. We noticed that the F4 had greater interaction with SAM1 and SAM2. However, they were still integrating inthe matrifocal subgroup.
The group structure in this phase was similar to that seen in the first phase. However, the coalition among the subordinated males diminished, and the M2, SAM3, and SAM4 were still out of the center of the group, remaining associated between themselves, even though the SAM1 and SAM2 exhibited greater affinity with the adult females F3, F4, and F5. The immigrants (F5, I3, and J6) seemed to be more integrated in the group in this phase, which could be explained by the birth of the I4.
The behavior of the infants and juveniles in this phase was more distant from the main group, perhaps in search for security, since males had interactions among themselves in the group's center,leading to higher possibilities of conflict. Another possibility is that, with maturity, exploratory environmental behavior andpsychomotor-related activities lead them to be more in the periphery of the group.
In the fourth phase, the birth of the I5 apparentlyinfluenced adult females, and all of them (including the immigrant F5) were thenpresent in the matrifocal subgroup. However, more juveniles and infants were in the center of this association. We could see an increase in the tendency of SAMs interacting with the adult females. Therefore, the coalition of the males became weaker and, in the last month of sampling (Oct/2001), the M2 was no longerpresent in the group. It is not known whether it was banishedfrom the group, if it emigrated or if it became a satellite male, but this fact could be related to the poor association of the M2 with the subadult males and with the M1, or, rather, it could be due to the group's higher instability related to the increase in the number of its individuals.
Interestingly, the M2 was still part of the group and did not stop interacting with the subadult males, even when the SAM1 and SAM2 - apparently the oldest of the subadult males - started to associate with thematrifocal subgroup and, consequently,with the M1. We are not sure if the closer proximity between these two SAMs and the M1 is related to a coalition between them or to competition to get access to females.However,no agonistic behavior betweenthese males was observed during this period, and, for this reason,we assume that this behavior is more likely to be a coalition among these males.
This type of male coalition was also observed by Jones (1980) in a A. palliata group. Most likely, this binding is related to the M2's departure from the group and its migration from the park to the city's surrounding areas. A few days after leaving, it was captured by the Municipal Zoo anddied one week later.
Despite the instability displayed by this group, we did not observe a more aggressive behavior -as we expected -, thus refuting our third hypothesis; we only saw two displaysof agonistic behavior involving two adult females. As mentioned before,the high supply of fruits during year probably reduced competition among the individuals, and the indirect signs were enough to establish and communicate the social hierarchy within this group. However, theaging of subadult males and a consequently increased competition for access to females may increase the aggressiveness between males.
CONCLUSION AND FINAL CONSIDERATIONS
We believe that our study provides an important interpretation on the social behavior of howler monkeys (Alouattacaraya). The patterns of social interaction observed during the samplings were efficiently confirmed by the multivariate methodology used in this research, which helped us in the identification of the social interaction patterns, illustrating the plasticity and tendency that the social structure suffered during the period of study.
This group showed a strong association among adult females (matrifocal subgroup),whereas the dominant male (M1) had greater proximity to them. Signs of instability were common throughoutthe year due to integration of "new" individuals, but the births of infants strengthened the matrifocal subgroup. The aging of subadult males increased their association with the matrifocal subgroup and with the M1, resulting in the departure of the other adult male from the group.
The social organization of this group could be assumed to be a harem or age-graded system, which experienced signs of instability, and the few agonistic episodes and inter-individual competition seemed to be related to the low competition for food, since the availability of fruits was high.
The A. carayaspecies is globally characterized as the "least concern - LC", despite its declining populations (Fernandez-Duque et al.,2008), butit was classified as a "near-threatened species" in Brazil (Ludwig et al., 2015); in the state list ofSão Paulo, the species appears as "vulnerable" (Bressan et al., 2009).
The "near threatened" and "vulnerable" statusesare very evident in our study area, where the remaining forests in the region are highly fragmented, coveringonly 4% of Ribeirão Preto's area (Henriques, 2003). Under these conditions, the geographic isolation leads to high degrees of endogamy, thus resulting in unviable populations in the long term.
The evaluation of the black-and-gold howler monkeys'group dynamics is therefore, of great importance to establish management plans for the conservation of primates that live under this type of environmental pressure. Hence, in this study, the high population density of the group and the resulting migration of an individual to the city area resultedin the relocation of some individuals to other forest fragments (Rossiand Santos, in press). In addition, the risk of extinction that exists for the Primates of the tropical area is considerable. Thus, our work can contribute to the knowledge of a Primate that is under high anthropic action.
ACKNOWLEDGEMENTS
The authors are thankful to PIBIC/USP/CNPq and Mr. Vinicio B. Pecci, from the Environmental Management and Planning Office of Ribeirão Preto City Hall, for the financial support (scholarships); to Mrs. Marisa dos Santos from Ribeirão Preto's City Zoo, who authorized the area of study;as well as to Drs. Gelson Genaro and Mr. Dilmar Oliveira, who provided suggestions and critical comments to improve this article. We would like to thank to Dr. Alexandre A. Oliveira for the help with the statistical analysis, and to Prof. Alexandra Olimpio O. Cunha and Prof. Abilio Borghi for their assistance regarding the English language.
BIBLIOGRAPHY
Altmann, S.A. (1959). Field observations on a howler monkey society. Journal of Mammalogy, 40, 317-330. [ Links ]
Altman, J. (1974). Observational Study of Behavior: Sampling Methods. Behaviour Leiden, 49, 227-267. [ Links ]
Azkarate, J.C., Dunn, J.C.,Balcells, C.D.,&Baró, J.V. (2017).A demographic history of a population of howler monkeys (Alouatta palliata) living in a fragmented landscape in Mexico.PeerJ, 5:e3547; DOI 10.7717/peerj.3547. [ Links ]
Bezanson, M., Garber P.A., Rutherford, J.,& Cleveland, A. (2002). Patterns of subgrouping, social affiliation and social networks in Nicaraguan mantled howler monkeys (Alouatta palliata). American Journal of Physical Anthropology, 34, 44. [ Links ]
Bezanzon, M, Garber, P.A., Murphy, J.T.,&Premo, L.S. (2008).Patterns of Subgrouping and Spatial Affiliation in A Community of Mantled Howling Monkeys (Alouatta palliata).American Journal of Primatology, 70, 282-293. [ Links ]
Bicca-Marques, J.C. (1991). "Ecologia e comportamento de um grupo de bugios pretos Alouatta caraya (Primates, Cebidae) em Alegrete, RS, Brasil". Master Thesis, Universidade de Brasília, Brasília-DF, Brasil. [ Links ]
Bressan, P.M., Kierulff, M.C.M.,& Sugieda, A.M. (2009). Fauna Ameaçada de Extinção do Estado de São Paulo: Vertebrados. Fundação Parque Zoológico de São Paulo, São Paulo. Secretaria de Meio Ambiente do Estado de São Paulo. 645p. [ Links ]
Brown, A. D.,&Zunino, G. E. (1994). Hábitat, densidad y problemas de conservación de los primates de Argentina. Vida Silvestre Neotropical, 3, 131-40. [ Links ]
Carpenter, C. R. (1934). A field study of the behavior and social relations of howling monkeys. Comparative psychology monographs,10, 1-168. [ Links ]
Clarke, K.R.,&Warwick, R.M. (2001). Primer v5: User Manual/Tutorial/Plymouth, U.K.: PRIMER-E Ltd. [ Links ].
Crockett, C.M.,& Eisenberg, J.F. (1987). Howlers: Variations in Group Size and Demography. InBB Smuts, DL Cheney, RM Seyfarth, RW Wrangham, TT Struhsaker (Eds.), Primates Societies(pp. 54-68). The University of Chicago Press, Chicago. [ Links ]
Dvoskin, R., Juárez, C.P.,& Fernandez-Duque, E. (2004).Population Density of Black Howlers (Alouatta caraya) in the Gallery Forests of the Argentinean Chaco: A Preliminary Assessment. Folia Primatologica, 75, 93-96. [ Links ]
Eisenberg, J.F., Muchenhim, N.A.,&Rudran, R. (1972).The relation between ecology and social structure in primates. Science, 4037, 53-79. [ Links ]
Estrada, A., Mendoza, A., Castellanos, L., Pacheco, R., van Belle, S., García,Y.,&Muñoz,D. (2002). Population of the Black Howler Monkey (Alouatta pigra) in a Fragmented Landscape in Palenque, Chiapas, Mexico.American Journal of Primatology, 55, 45-55. [ Links ]
Fernandez-Duque E., Wallace. R.B.,&Rylands, A.B. (2008). Alouatta caraya. The IUCN Red List of Threatened Species, e.T41545A10496784. [ Links ]
Flórez, J.C.M.,& Delgado, J.A.L. (2011). Population Density of the Red Howler Monkey (Alouatta seniculus) in a Tropical Dry Forest Fragment in Northwestern Colombia. Neotropical Primates, 18(2), 64-68. [ Links ]
Garber, P.A. (1987). Foraging strategies among living primates. Annual Review of Anthropology, 16, 339-364. [ Links ]
Glander, K.E. (1975). Habitat description and resource utilization: A preliminary report on mantled howling monkey ecology.In RH Tuttle (ed.),Sociecology and Psychology of Primates(pp. 37-57). The Hague, Mouton. [ Links ]
Gomes, H.L. (2004). "Ecologia alimentar e comportamento geral de Bugios-pretos (Alouatta caraya, Primates, Atelidae) em mata urbana de Ribeirão Preto/SP". Master Thesis, Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Hair, J.F., Anderson, R.E., Tatham, R.L.,& Black, W.C. (1995). Multivariate Data Analysis with Readings. Prentice Hall Inc Ed. New Jersey - USA. [ Links ]
Henriques, O.K. (2003). "Caracterização da vegetação natural em Ribeirão Preto, SP: Bases para conservação". Master Thesis, Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Jones, C.B. (1980). The functions of status in the mantled howler monkeys, Alouatta palliata Gray: intraespecific competition for group membership in a folivorous Neotropical primate. Primates, 21, 389-405. [ Links ]
Jones, C.B. (1982). A field manipulation of spatial relations among male mantled howler monkeys. Primates, 23 (1), 130-134. [ Links ]
Jones C.B. (1983). Social Organization of captive black howler monkeys (Alouatta caraya): "Social competition" an use of non-damaging behavior. Primates, 24, 25-39. [ Links ]
Juárez, C.P., Dvoskin, R., & Fernández-Duque, E. (2005). Structure and Composition of Wild Black Howler Troops (Alouatta caraya) in Gallery Forests of the Argentinean Chaco. Neotropical Primates, 13(1), 19-22. [ Links ]
Kappeler, P.M., &Van Schaik, C.P. (2002). Evolution of Primate Social Systems. International Journal of Primatology,23(4), 707-740. [ Links ]
Kinzey, W.G., & Cunningham, E.P. (1994). Variability in platyrrhine social organization. American Journal of Primatology34(2), 185-198. [ Links ]
Kowalewski, M.M., &Zunino, G.E. (1999). Impact of Deforestation on a Population of Alouatta caraya in Northern Argentina.Folia Primatologica, 70, 163-166. [ Links ]
Kowalewski, M.M. (2000). "Birth seasonality in black howler monkeys (Alouata caraya) in an island system in northern Argentina". Master thesis, State University of New York at Stony Brook, New York, USA. [ Links ]
Laguna, V.G. (1997). "Levantamento Florístico Fitossociológico em remanescente Floresta Mesófila Semidecídua Urbana - Parque Municipal Morro de São Bento, Ribeirão Preto, SP". Monography Thesis, Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Laguna, V.G. (2000). "Estrutura e Diversidade do Remanescente de Floresta Estacional Semidecidual do Parque Municipal Morro de São, Ribeirão Preto, SP. Ribeirão Preto (SP)". Master Thesis, Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Ludwig, G., Bicca-Marques, C., Rímole, J., Cunha, R.G.T., Alves, S.L., Martins, V., Valle, R.R., Miranda, J.M.D., & Messias, M.R. (2015). Avaliação do Risco de Extinção de Alouatta caraya (Humboldt,1812) no Brasil. Processo de avaliação do risco de extinção da fauna brasileira. ICMBio. http://www.icmbio.gov.br/portal/biodiversidade/fauna-brasileira/estado-de-conservacao/7176-mamiferos-alouatta-caraya-bugio-preto.html [ Links ]
Milton, K. (1999). Nutritional characteristics of wild primate foods: do the diets of our closest living relatives have lessons for us? Nutrition,15(6), 488-98. [ Links ]
Milton, K. (2000). Back to basics: why foods of wild primates have relevance for modern human health. Nutrition, (7-8), 480-483. [ Links ]
Miranda, J.M.D.,& PASSOS, F.C. (2005). Composição e dinâmica de grupos de Alouatta guaribaclamitans Cabrera, 1940 (Primates, Atelidae) em Floresta Ombrófila Mista no Estado do Paraná, Brasil. Revista Brasileira de Zoologia, Curitiba, 22(1), 99-106. [ Links ]
Neville M.K. (1972). Social relations within troops of Red Howler Monkeys (Alouatta seniculus). Folia Primatologica, 18, 47-77. [ Links ]
Neville, M.K., Glander, K.E., Braza, F, &Rylands, A.B. (1988). The Howler Monkeys, genus Alouatta. In: R.A. Mittermeier, A.B., Rylands, A.F., Coimba-Filho, &G.A.B. da Fonseca. (eds). Ecology and Behavior of Neotropical Primates (pp. 349-453). World Wild Life Fund. Washington, D.C. [ Links ]
Oliveira, D.A.G.,& Ades, C. (1998). Proximity and grooming interactions as indicators of the social organization of brown howling monkeys (Alouatta fusca clamitans). Neotropical Primates, 6(4), 115-117. [ Links ]
Pavelka, M.S.M.,&Knopff, K. H. (2004). Diet and activity in black howler monkeys (Alouatta pigra) in southern Belize: does degree of frugivory influence activity level? Primates,54, 105-111. [ Links ]
Pedrosa J.M. (2002). "Ecologia Comportamental de um Grupo de Bugios Pretos (Alouatta caraya, Primates, Cebidae) o fragmento de mata Parque Municipal Morro de São Bento, Ribeirão Preto-SP". Monography Thesis. Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Rossi, M.J.,& Santos, W.F. (in press). Births during 7 years after the translocation of a pair of black howler monkeys (Alouatta caraya) to a forest fragment in southeast Brazil. Primates. [ Links ]
Rumiz, D.I. (1990). Alouata caraya: Population density and demography in Northern Argentina. of Primatology, 21, 279-294. [ Links ]
Sampaio,R. (2002). "Estrutura social e hierárquica de um grupo de bugios pretos (Alouatta carayaHumboldt, 1812, Primates, Atelinae) em um fragmento de mata em Ribeirão Preto/SP". Monography Thesis. Universidade de São Paulo, Ribeirão Preto, Brasil. [ Links ]
Southwick, C.H. (1963). Challenging aspects of the behavioral ecology of howling monkeys. In Southwick, C.H. (ed.), Primate Social Behavior (pp. 185-191). D. Van Nostrand, Princeton, NJ. [ Links ]
Strier, K.B. (1992). Atelinae adaptations: Behavioral strategies and ecological constraints. American Journal of Physical Anthropolgy,88(4), 515-524. [ Links ]
Wang, E.,& Milton, K. (2003). Intragroup Social Relationships of Male Alouatta palliata on Barro Colorado Island, Republic of Panama. International Journal of Primatology, 24(6), 1227-1243. [ Links ]
* Corresponding author: Wagner Ferreira Dos Santos. Av. Bandeirantes, 3900. Ribeirão Preto,São Paulo, Brazil. CEP: 14040-901
1 We are not sure if these individuals were living isolated from the focal groups or if they had moved away from the group for a certain period of time.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}