










Services on Demand

article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkRevista de Etologia
Print version ISSN 1517-2805
Rev. etol. vol.9 no.2 São Paulo Dec. 2010
ARTIGO
Interação competitiva de Coragyps atratus (Bechstein, 1793) (Cathartiformes, Cathartidae) no uso de um recurso limitado
Competitive interaction of Coragyps atratus (Bechstein, 1793) (Cathartiformes, Cathartidae) in the use of a limited resource
Leonardo Barros Ribeiro; Daniela Gomes de Lima; Silvia Beatriz Fonseca de Melo
Universidade Federal do Rio Grande do Norte
RESUMO
Descreve-se a interação e o uso do espaço entre espécimes de urubus-pretos (C. atratus) no consumo de um recurso limitado na forma de uma fonte de água temporária, em uma área do nordeste do Brasil, em época de seca. As aves mostraram uma frequência de agressividade elevada pelo aumento do número de indivíduos realizando o comportamento de beber água, e uma distribuição dos indivíduos aparentemente hierárquica no acesso ao recurso.
Palavras-chave: Competição por recursos. Aquisição de água. Aves necrófagas. Urubu- preto. Nordeste do Brasil.
ABSTRACT
The interaction and space use among the specimens of Black Vultures (C. atratus) consuming a limited resource in the form of a temporary water source, during drought period, at one site in northeastern Brazil is described. The birds showed an aggressiveness frequency elevated by the increase of the number of individuals accomplishing behavior of drinking water, and a distribution of the individuals apparently hierarchic in the access to the resource.
Keywords:Competition for resource. Water acquisition. Avian scavenger. Black vultures. Northeastern Brazil.
Quando recursos limitados são encontrados amplamente dispersos, na forma de manchas ricas, efêmeras e imprevisíveis em ocorrência espacial e temporal, a competição entre os consumidores pode envolver exploração diferencial ou de interferência. A exploração diferencial está frequentemente baseada na habilidade diferencial dos indivíduos em localizar o recurso, e a interferência normalmente é alcançada através do estabelecimento de hierarquias de dominância entre os indivíduos que encontram o recurso (Maurer, 1984). Aves necrófagas que se alimentam de carcaças de grandes animais competem por tal recurso limitado, e as diferenças na ordem de chegada à carcaça ou na utilização de qualquer outra fonte de recurso, assim como a hierarquia de dominância quando as aves estão juntas, são estabelecidas entre os membros do grupo (Kruuk, 1967; Houston, 1975).
Coragyps atratus, conhecido como urubu-preto ou urubu-de-cabeça-preta, está incluído no grupo de urubus do Novo Mundo, encontrado na América do Norte e América do Sul em diversos tipos de hábitats e bem adaptado à fragmentação da paisagem causada pela atividade humana (Kirk & Mossman, 1998). Apresenta tamanho médio variando entre 60 e 68 cm de comprimento e massa corpórea em torno de 1,6 a 2,2 quilogramas, sendo uma espécie sexualmente monomórfica. Possui sistema olfatório reduzido e geralmente encontram alimento em áreas abertas através da visualização ou seguindo outros animais (Buckley, 1999). Estas aves não são territoriais e sobrevoam extensas áreas em busca de fontes de recursos, em volta das quais formam grandes agregações (Rabenold, 1987a; Coleman & Fraser, 1989). Formas de agrupamento em aves têm atraído a atenção de pesquisadores por muitas décadas, e um exemplo interessante é encontrado em C. atratus, onde há benefícios individuais no agrupamento como troca de informações (Rabenold, 1987b), facilitação de forrageamento (Buckley, 1996; Stolen, 2000), oportunidades de interação social (Stolen, 1996) e possível proteção de predadores (Buckley, 1996).
Os estudos mais detalhados de interações competitivas entre aves necrófagas aconteceram na África, onde grandes guildas de urubus do Velho Mundo (Accipitridae) alimentam-se de carcaças de ungulados (Petrides, 1959; Attwell, 1963; Kruuk, 1967; Houston, 1974, 1975). Em contraste, existem poucas descrições, muitas destas anedóticas (Koford, 1953; Stuart, 1978), de interações competitivas entre guildas de urubus do Novo Mundo (Cathartidae). Pouco se encontra na literatura sobre competição interespecífica incluindo C. atratus (dois estudos de destaque são Wallace & Temple, 1987 e Hiraldo, Delibes & Donazar, 1991), não sendo encontrado nenhum registro de competição intraespecífica no uso de água como recurso. Neste sentido, este estudo analisa pela primeira vez a interação competitiva apresentada por C. atratus, na utilização de um recurso limitado na forma de uma fonte de água temporária, através do registro dos principais atos comportamentais.
O estudo foi realizado na Floresta Nacional, FLONA-IBAMA (6°5’S, 35°12’W), no município de Nísia Floresta a 45 km da cidade de Natal, Rio Grande do Norte, Brasil. A FLONA possui 170 hectares divididos em fragmentos florestais, sendo 60% correspondente à Mata Atlântica e sistemas associados, 38% de área de experimentação e 2% de área urbanizada (administração, refeitório, garagens, residências). A região é a maior área do Estado com clima subúmido, onde os meses chuvosos são de março a agosto, e os meses secos, de setembro a fevereiro. A temperatura média anual da área é de 28°C.
A fonte de água visitada pelos urubus compreendeu uma área de aproximadamente 15 x 10 m, com profundidade máxima de 8 cm, e apresentando apenas uma margem livre de vegetação herbácea (15 m de extensão) que permitia acesso facilitado dos urubus à água. O estudo foi realizado em um período de plena seca e esta única lâmina de água localizada na FLONA representou um importante recurso.
Os animais foram observados no período de 12 a 14 de dezembro de 2006, durante duas sessões diárias com duração de quatro horas cada, uma pela manhã (das 7h às 11h) e outra à tarde (das 13h às 17h), totalizando 24 horas de observação. As observações foram anotadas sempre por um par de observadores, a olho nu, mantendo-se uma distância de aproximadamente 10 m dos animais, e camuflados pela vegetação local para não influenciarem no seu comportamento. A identificação das aves foi feita por meio de guia de campo segundo Sick (1997) e Souza (1998).
Para observação dos animais foi utilizado o método Scan Sampling (Altmann, 1974) em intervalos regulares de 5 minutos para verificar o número e a distribuição dos indivíduos em torno do recurso, assim como a frequência dos comportamentos exibidos durante sua utilização. Estabeleceram-se quatro categorias comportamentais: parado (animal sem deslocamento, deitado ou em pé), deslocando (andando, correndo, pousando ou alcançando vôo), bebendo (ingerindo água em qualquer local da fonte) e agressividade (perseguindo ou bicando um coespecífico).
Para a análise da distribuição espacial dos urubus na fonte de água foram delimitadas quatro regiões: lâmina de água, área 1 (A1), área 2 (A2) e área 3 (A3), tendo as parcelas de solo (referentes às áreas de 1 a 3) uma largura de 3 m. Estas áreas foram demarcadas no chão com o uso de ramos de vegetação, os quais eram visualmente localizados pelos observados. Ainda no limite final da área 3, uma árvore (imbaúba, Cecropia sp.) foi utilizada como poleiro pelos urubus, e a presença destes nesta planta também foi registrada. É importante ressaltar que no instante das observações cada pesquisador era responsável pelo registro das aves e seus respectivos comportamentos em duas regiões: lâmina de água-A1 e A2-A3 (considerando a árvore).
Os testes t de Student, qui-quadrado e correlações de Spearman (Zar, 1999) foram utilizados para análise das categorias comportamentais. A probabilidade (p) de cometer erro tipo I foi assinalada em cada teste realizado, considerando-se estatisticamente significantes valores de "p" menores que 0,05. As prevalências da amostra total foram apresentadas com intervalos de confiança de 95%.
Durante os três dias de experimento, os espécimes de C. atratus apresentaram frequências de visitas equivalentes a 30% (n = 879), 22% (n = 659) e 48% (n = 1404), respectivamente no primeiro, segundo e terceiro dias. Em relação ao turno do dia, o período da manhã correspondeu ao maior número de registros, 58% de visitas, e 42% à tarde. A ocorrência das aves por turno, apesar de muito próxima, apresentou diferença estatística significativa (X2 = 10,477; GL = 1; p = 0,033). Após o período noturno de descanso, a reposição das reservas corporais de água para o início do forrageio parece suportar esta maior frequência de visitas pela manhã.
Em relação aos comportamentos registrados ao longo das observações, os mais frequentes foram "parado" (81%) e "bebendo" (9%), seguidos de "deslocando" (8%) e "agressividade" (2%). Ao analisarmos os comportamentos por dia de observação (Tab. 1), um maior índice de agressividade foi encontrado para o segundo dia, embora o número de registros de aves neste dia tenha sido o menor (n = 659). Este resultado esteve relacionado com o aumento no comportamento de beber água, o qual foi sensivelmente maior do que nos demais dias e correlacionou-se positiva e significativamente com a "agressividade" (Correlação de Spearman; rs = 0,767; p < 0,0001). A Figura 1 representa os percentuais para as categorias comportamentais em relação aos turnos de observação. Destes dados, destaca-se a categoria "bebendo", que obteve maior valor no turno da manhã do que à tarde (Teste t; t = -4,00; p = 0,0001), e o mesmo padrão de aumento foi observado em relação à "agressividade" (Teste t; t = -2,37; p = 0,024). A agressividade também apresentou correlação positiva com o deslocamento (Correlação de Spearman; rs = 0,663; p < 0,0001), apesar de ter havido diminuição no "deslocar" no segundo dia de observação. Berryman (1999) e Turchin (2003) afirmaram que os mecanismos de competição intraespecífica que incluem agressividade, territorialidade e a competição por espaços livres, aumentam com a densidade populacional crescente, e segundo Berryman (2003), os indivíduos competem entre si para ocupar a quantidade insuficiente de espaço disponível.
Tabela 1. Percentuais dos comportamentos apresentados diariamente por C. atratus observados na Floresta Nacional, FLONA-IBAMA, no município de Nísia Floresta, Rio Grande do Norte.
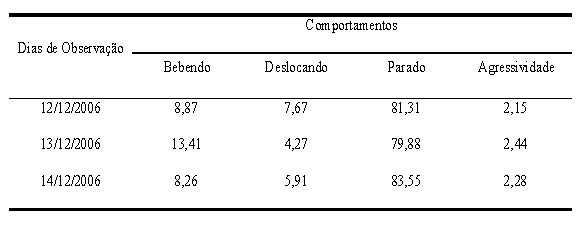
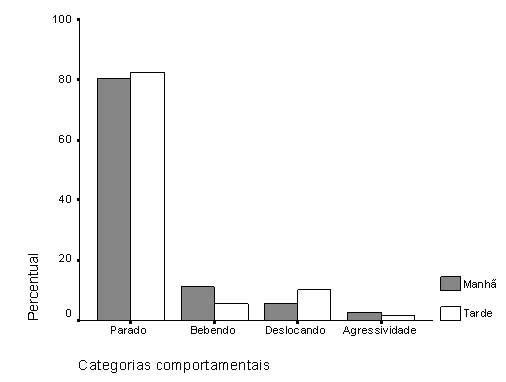
Figura 1. Percentual relativo à frequência das categorias comportamentais analisadas para Coragyps atratus, durante os períodos manhã e tarde na Floresta Nacional, FLONA-IBAMA, no município de Nísia Floresta, Rio Grande do Norte.
Dentre as áreas do solo disponíveis para a visita ao recurso, destacaram-se A1 (34%) e A2 (31%) como as mais utilizadas, muito provavelmente por serem mais próximas à lâmina de água, garantindo assim um acesso mais rápido e facilitado ao recurso. Por outro lado, a árvore destacou-se como o terceiro local de maior utilização (15%), e representou um ponto de observação em potencial para as aves pousarem antes de efetivamente chegarem ao solo. Finalmente, lâmina de água e A3 caracterizaram respectivamente 11% e 9% de uso. O uso das áreas por turnos do dia está representado na Figura 2. De um modo geral, ainda que A1 e A2 tenham apresentado maior percentual de uso, o período de utilização das mesmas apresentou características opostas quanto aos turnos do dia. Houve maior utilização de A1 no turno da manhã (46,29%), enquanto prevaleceu o uso de A2 no turno da tarde (48,06%). Como o comportamento de beber água apresentou o maior índice no turno da manhã (11,33%) do que à tarde (5,67%), esperar-se-ia que os animais se distribuíssem na área de maior proximidade à água (A1), o que de fato ocorreu. No período da tarde, a presença das aves em A2 limitou-se a atos de descanso.
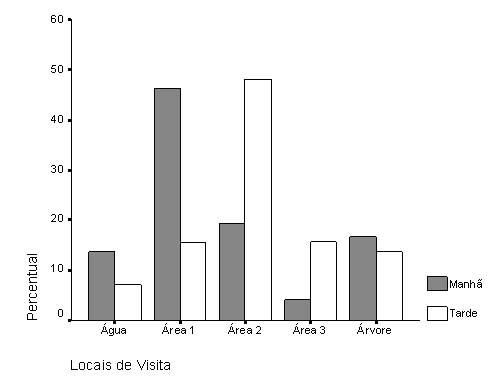
Figura 2. Percentual relativo à presença de espécimes de Coragyps atratus nas regiões delimitadas para observação, durante os períodos manhã e tarde na Floresta Nacional, FLONA-IBAMA, no município de Nísia Floresta, Rio Grande do Norte.
Wallace e Temple (1987) observaram uma hierarquia de dominância em C. atratus para o acesso ao recurso alimentar, de forma que durante os encontros agonísticos, os jovens foram muito mais submissos aos adultos, os quais venceram 93% das interações. Na utilização da fonte de água foi observada a presença de determinados espécimes esperando os coespecíficos saírem da lâmina de água para que também conseguissem beber. Embora não tenha sido possível determinar se estes indivíduos eram jovens, o comportamento de espera, de certa forma, mostra a existência de uma hierarquia de dominância, corroborando os estudos de Kruuk (1967) e Houston (1975), os quais observaram que uma ordem de chegada ou de utilização de recursos é estabelecida entre os membros do grupo de aves necrófagas.
Os resultados permitem concluir que os espécimes de C. atratus mostram um aumento na frequência de agressividade com o respectivo aumento do número de indivíduos realizando o comportamento de beber água, em se tratando de uma fonte escassa e limitada. Podendo-se ainda inferir que nas condições semelhantes às estudadas, estas aves exibem uma hierarquia no acesso e uso de recursos.
Referências
Altmann, J. (1974). Observational study of behaviour: sampling methods. Behaviour, 49,227-267. [ Links ]
Attwell, R. I. G. (1963). Some observations on feeding habits, behaviour and inter-relationships of northern Rhodesian vultures. Ostrich, 34,235-247. [ Links ]
Berryman, A. A. (1999). Principles of population dynamics and their application. Cheltenham, UK. [ Links ]
Berryman, A. A. (2003). On principles, laws and theory in population ecology. Oikos, 103,695-701. [ Links ]
Buckley, N. J. (1996). Food finding and the influence of information, local enhancement, and communal roosting on foraging success of North American vultures. Auk, 113,473-488. [ Links ]
Buckley, N. J. (1999). Black vulture (Coragyps atratus). In A. Poole & F. Gills (Eds.), The Birds of North America. Philadelphia, Pennsylvania. [ Links ]
Coleman, J. S., & Fraser, J. D. (1989). Habitat use and home ranges of Black and vultures. Journal of Wildlife Management, 53,782-792. [ Links ]
Hiraldo, F., Delibes, M., & Donazar, J. A. (1991). Comparison of diets of turkey vultures in three regions of northern Mexico. Journal of Field Ornithology, 62,319-324. [ Links ]
Houston, D. (1974). The role of griffon vultures Gyps africanus and Gyps ruppellii as scavengers. Journal of Zoology of London, 172,35-46. [ Links ]
Houston, D. (1975). Ecological isolation of African scavenging birds. Ardea, 63,55-64. [ Links ]
Kirk, D. A., & Mossman M. J. (1998). Turkey vulture (Cathartes aura). In A. Poole & F. Gill (Eds), The birds of North America. The Academy of Natural Sciences, Philadelphia, Pennsylvania, USA, and The American Ornithologists Union, Washington, D.C., USA. [ Links ]
Koford, C. B. (1953). The California Condor. National Audubon Research Report, 4,1-154. [ Links ]
Kruuk, H. (1967). Competition for food between vultures in East Africa. Ardea, 55,171-193. [ Links ]
Maurer, B. A. (1984). Interference and exploitation in bird communities. Wilson Bulletin, 96,380-395. [ Links ]
Petrides, G. A. (1959). Competition for food between five species of East African vultures. Auk, 76,104-106. [ Links ]
Rabenold, P. P. (1987a). Recruitment to food in black vultures: Evidence for following from communal roosts. Animal Behaviour, 35,1775-1785. [ Links ]
Rabenold, P. P. (1987b). Roost attendance and aggression in Black vultures. Auk, 104,647-653. [ Links ]
Sick, H. (1997). Ornitologia Brasileira. Rio de Janeiro: Nova Fronteira. [ Links ]
Souza, D. G. S. (1998). Todas as aves do Brasil - Guia de campo para identificação. Feira de Santana: Dall. [ Links ]
Stolen, E. D. (1996). Roosting behavior and foraging ecology of Black Vultures in central Florida. M.Sc. Thesis, University of Central Florida, Orlando. [ Links ]
Stolen, E. D. (2000). Foraging behavior of vultures on central Florida. Florida Field Naturalist, 28,173-181. [ Links ]
Stuart, P. (1978). Behavioral interactions and niche separation in Black and Turkey Vultures. Living Bird, 17,79-84. [ Links ]
Turchin, P. (2003). Complex population dynamics: A theoretical/empirical synthesis. USA: Princeton University Press. [ Links ]
Wallace, M. P., & Temple, S. A. (1987). Competitive interactions within and between species in a guild of avian scavengers. Auk, 104,290-295. [ Links ]
Zar, J. H. (1999). Biostatistical analysis 4th ed. New Jersey, USA: Upper Saddle River, Prentice-Hall Inc. [ Links ]
Recebido em: 24/10/2009
Aceito em: 20/01/2012

Silvia Beatriz Fonseca de Melo
Programa de Pós-Graduação em Psicobiologia
Universidade Federal do Rio Grande do Norte
Centro de Biociências
Departamento de Fisiologia
Caixa Postal 1511, Campus Universitário Lagoa Nova, Cep 59078-970, Natal, RN, Brasil
Endereço atual (L.B. Ribeiro)
Universidade Federal do Vale do São Francisco
Campus Ciências Agrárias
Colegiado de Ciências Biológicas
CEP 56300-990, Petrolina, PE, Brasil.
e-mail: leonardo.ribeiro@univasf.edu.br; danielagomesdelima@gmail.com; silviabeatriz28@gmail.com.
Este trabalho é resultado das disciplinas de Ecologia Comportamental e Comportamento Animal. Agradecemos a Professora Dra. Fátima Arruda pela sugestão na coleta de dados, aos revisores anônimos pelas sugestões no texto e ao CNPq pelo auxílio financeiro a L. B. Ribeiro e D. G. Lima.
