










Services on Demand

article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkPsicologia USP
On-line version ISSN 1678-5177
Psicol. USP vol.4 no.1-2 São Paulo 1993
ARTIGOS ORIGINAIS
Psicobiologia da memória1
The psychobiology of memory
Carlos Tomaz
Laboratório de Psicobiologia - Faculdade de Filosofia, Ciências e Letras - USP - Ribeirão Preto
RESUMO
O objetivo deste artigo é introduzir o leitor na área de pesquisa sobre a psicobiologia da memória e apresentar alguns resultados recentes dessas pesquisas. A pesquisa sobre a psicobiologia da memória está centrada em dois aspectos: (1) identificação de áreas cerebrais envolvidas na regulação do armazenamento da memória; e (2) identificação das alterações celulares e neuroquímicas que ocorrem entre as células nervosas durante a aquisição da informação. Resultados recentes dessas pesquisas indicam que diferentes áreas do cérebro e diferentes neurotransmissores estão relacionados a diferentes tipos de memória.
Descritores: Memória. Psicobiologia. Cérebro. Drogas.
ABSTRACT
The aim of this paper is to introduce the reader to the research field of psychobiology of memory and show some recent findings. Research on the psychobiology of memory is centered on two major issues: (1) identification of brain areas involved in the regulation of memory storage; and (2) identification of the cellular and neurochemical changes mediating experience-induced changes in interactions among neurons. Recent research findings indicate that different brain areas and different neurotransmitters are related to different kinds of memory.
Index terms: Memory. Psychobiology. Brain. Drugs.
Memória é a capacidade de o indivíduo se situar no presente levando em consideração o passado e o futuro. A memória fornece as bases para todos os nossos conhecimentos, habilidades, sonhos, planos e anseios. Memória é, por conseguinte, um fator determinante do nosso comportamento. Assim, o conhecimento sobre a natureza e as bases biológicas da memória é essencial para o entendimento da psique humana.
Vários laboratórios em Universidades e indústrias farmacêuticas possuem programas de pesquisa que investigam as bases neurobiológicas da memória. Existem diversas razões para isso. Primeiro, como citado anteriormente, o fenômeno da memória é um aspecto central da existência humana. Desta forma, o entendimento dos processos cerebrais subjacentes à memória irá se constituir num evento científico da maior importância. Segundo, os avanços científicos e tecnológicos das últimas décadas têm apontado para um intrigante processo de plasticidade cerebral. Terceiro, a compreensão dos mecanismos cerebrais envolvidos na memória é essencial para o desenvolvimento de terapias relacionadas com as doenças cognitivas.
A pesquisa básica que investiga a psicobiologia da memória está centrada em dois aspectos. O primeiro é aquele que diz respeito à identificação das áreas cerebrais envolvidas neste fenômeno bem como ao papel dos diferentes sistemas cerebrais na regulação do armazenamento das lembranças e na mediação dos diferentes tipos de memória. O segundo, está relacionado à identificação das alterações celulares e bioquímicas que ocorrem entre neurônios durante as modificações induzidas pela aquisição da informação.
Resultados recentes dessas pesquisas indicam que a memória pode ser influenciada por alterações nas funções de determinados sistemas cerebrais, assim como por alterações entre interações celulares em áreas cerebrais específicas. Isto é, diferentes áreas do cérebro e diferentes neurotransmissores estão relacionados à memória, e até mesmo, a diferentes tipos de memória. Por exemplo, lesões do hipocampo, uma estrutura cerebral localizada na região medial do lóbulo temporal do cérebro humano (ver Figura 1), impedem seletivamente a memorização de novas informações sem afetar a aprendizagem perceptual e as habilidades motoras (Squire, 1987). Em animais de laboratório, lesões do hipocampo também bloqueiam a retenção de informações relacionadas ao ambiente espacial, e alterações no funcionamento da amígdala (uma estrutura localizada próximo à extremidade do hipocampo; ver Figura 1), prejudicam a memória relacionada a experiências emocionais (LeDoux, 1992 e Parent; Tomaz; McGaugh, 1992). A Figura 1 apresenta um desenho esquemático das principais estruturas cerebrais relacionadas à modulação da memória.
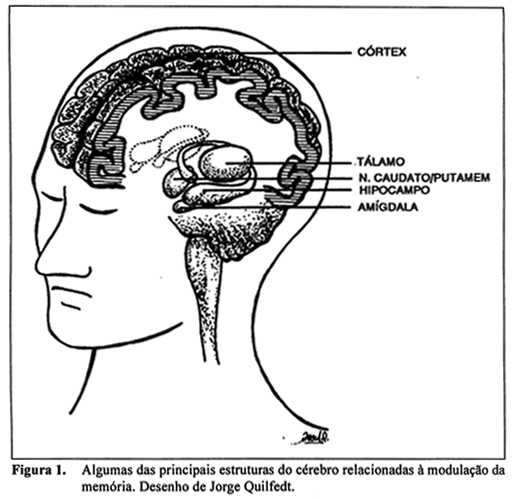
Várias regiões do cérebro influenciam a memória alterando a atividade neuronal em outras regiões do cérebro através da liberação de neurotransmissores e neuromoduladores (moléculas que permitem a comunicação entre células nervosas no sistema nervoso). Por exemplo, células do prosencéfalo basal se projetam para muitas regiões do cérebro, incluindo o hipocampo e o córtex e afetam o funcionamento destas liberando o neurotransmissor acetilcolina e ativando receptores colinérgicos sobre os neurônios nestas regiões. Células do locus coeruleus, um núcleo localizado no tronco cerebral, também se projetam para diversas regiões do cérebro, incluindo a amígdala e o córtex, e influenciam a atividade neuronal dessas regiões pela liberação de norepinefrina e a conseqüente ativação de receptores noradrenérgicos. Sabe-se que em pessoas portadoras da doença de Alzheimer, do qual um dos sintomas é a profunda perda da memória, ocorre um acentuado déficit no funcionamento do sistema colinérgico do prosencéfalo basal e do locus coeruleus. Por outro lado, em animais de laboratório, a lesão induzida experimentalmente dessas regiões cerebrais produz um profundo déficit na memória (Olton;Wenk, 1987).
Existem diversas evidências clínicas e experimentais indicando que os processos de armazenamento de memória são suceptíveis a manipulações que afetam o funcionamento cerebral. Por exemplo, amnésia retrógrada (perda de memória para fatos antecedentes) pode ser induzida pela administração de choque eletroconvulsivo ou pela administração de drogas que alteram os sistemas neurotransmissores no cérebro (McGaugh, 1989).
Os resultados obtidos com as drogas estimulantes, mostrando que os efeitos só são obtidos quando as substâncias são administradas logo após o treino, indicam que esses efeitos estão associados com os processos neurais que participam no armazenamento da aprendizagem imediatamente após a aquisição da informação.
Outros estudos têm demonstrado que hormônios liberados durante situações estressantes são capazes de alterar a memória. Por exemplo, a administração de epinefrina, um hormônio adrenérgico liberado pela medula adrenal diante de situações estressantes, pode facilitar ou bloquear a memória dependendo da dose injetada: a retenção da aprendizagem é facilitada com baixas doses e impedida com altas doses (McGaugh; Gold, 1989). É importante mencionar que esses efeitos têm sido obtidos até mesmo em testes de aprendizagem que utilizam recompensas agradáveis. Isto indica que a situação de aprendizagem não precisa ser altamente emocional para que a epinefrina produza seus efeitos.
Sabe-se, também, que situações emocionalmente estressantes ativam os sistemas opióides peptidérgicos (sistema neuromodulador constituído por pequenas moléculas onde se destacam as encefalinas, as endorfinas e as dinofirnas que causam efeitos semelhantes aos da morfina). Vários estudos, dentre os quais os realizados em nosso laboratório (Tomaz; Aguiar; Nogueira, 1990), têm demonstrado que a administração de drogas bloqueadoras do sistema opióide, tais como naloxona e naltrexona, facilita a memória.
O ácido gama-aminobutírico (GABA), um neurotransmissor inibitório do sistema nervoso central, está associado às emoções e memória. Pesquisas por Graeff e colaboradores (Graeff, 1990) têm demonstrado que o GABA desempenha um importante papel na elaboração e expressão das emoções. Drogas que facilitam a transmissão do GABA (GABA agonistas) produzem um efeito ansiolítico, tranqüilizante, enquanto drogas que bloqueiam a transmissão GABAérgica (GABA antagonistas) induzem ansiedade, aumento da atividade locomotora e, até mesmo, convulsões. Por outro lado, sabe-se que a memória pode ser modulada por drogas que alteram a neurotransmissão do GABA (Castellano et al., 1990). Experimentos conduzidos em vários laboratórios, têm demonstrado que a administração de antagonistas GABAérgicos (picrotoxina e bicuculina), em doses subconvulsivantes, facilitam a memória em testes com animais de laboratório e que agonistas GABAérgicos (muscimol e baclofem) prejudicam a memória.
As benzodiazepinas (BZs), uma classe de drogas que facilitam a ação do GABA, também prejudicam a memória (Lister, 1985). Essas drogas são geralmente empregadas como ansiolíticos, anticonvulsivantes e relaxante muscular. Diferentemente das outras drogas aqui citadas, os efeitos amnésicos das benzodiazepinas foram primeiro observados com humanos. Logo após a introdução da benzodiazepina diazepam na prática clínica, surgiram vários relatos indicando a ocorrência de amnésia anterógrada (incapacidade de armazenar informações a longo prazo) em pacientes tratados com esta droga. Esses dados foram posteriormente confirmados em experimentos de laboratório com animais. As BZs, enquanto tranqüilizantes, são as drogas mais prescritas e vendidas no mundo. Dada a sua importância, diversos grupos de pesquisa têm estudado os mecanismos neurais através dos quais as BZs produzem amnésia.
Várias pesquisas têm demonstrado que a amígdala desempenha um papel importante no controle das emoções e dos processos associados à memória (Tomaz, 1992). Resultados indicam que as propriedades ansiolíticas das BZs são devidas, em parte, a uma facilitação da neurotransmissão GABAérgica na amígdala. Estudos têm também demonstrado que a amígdala possui altas concentrações de receptores para moléculas do tipo GABA/BZ. Microinjeção de diazepam dentro da amígdala produz efeitos ansiolíticos, tranqüilizantes e anticonvulsivantes comparáveis àqueles obtidos com injeção periférica (Nagy; Zámbó; Decsi, 1979).
Outros estudos têm demonstrado que os efeitos mnemônicos de drogas GABAérgicas são mediados, pelo menos em parte, na amígdala. Microinjeções de drogas GABAérgicas na amígdala produzem efeitos mnemônicos semelhantes àqueles obtidos com administração periférica (Brioni; Nagahara; McGaugh, 1989). Lesões da amígdala bloqueiam os efeitos da administração periférica dessas drogas (Ammassari-Teule; Pavone; Castellano; McGaugh, 1991).
Recentemente, trabalhando em colaboração com J. McGaugh e H. Dickinson-Anson na Universidade da Califórnia, EUA, nós verificamos que lesões da amígdala aboliam os efeitos amnésicos produzidos pela administração periférica da BZ diazepam (Tomaz; Dickinson-Anson; McGaugh, 1991). Como a amígdala é uma estrutura complexa, composta por vários grupos celulares, num estudo subseqüente, nós investigamos a possibilidade de haver uma especificidade desses diferentes núcleos na modulação dos efeitos amnésicos produzidos pelo diazepam. Para este mapeamento funcional da amígdala foram feitas lesões circunscritas dos diferentes núcleos da amígdala. Os resultados demonstraram que o núcleo basolateral da amígdala é a principal região mediadora dos efeitos amnésicos produzidos pelo diazepam (Tomaz; Dickinson-Anson; McGaugh, 1992). Isto não significa, porém, que esta é a única região do cérebro que participa na modulação dos efeitos mnemônicos produzidos pelas benzodiazepinas. Trabalhos recentes realizados por Izquierdo e colaboradores (por exemplo, Izquierdo; Medina, 1991) sugerem uma participação das BZs endógenas no septo e hipocampo, além da amígdala, na modulação de diferentes tipos de memória. Interessante, entretanto, é o fato de que o núcleo basolateral da amígdala é o principal sítio de ação ansiolítica das BZs. Assim, os efeitos ansiolíticos e amnésicos das BZs parecem envolver os mesmos mecanismos neurais ou mecanismos sobrepostos na amígdala (ver Tomaz; Dickinson-Anson; McGaugh, 1992).
Estudos anatômicos têm demonstrado que a amígdala, e em particular o núcleo basolateral, recebe informações de todas as modalidades sensoriais, e envia informações para diferentes áreas do cérebro como o tálamo, neocortex, hipocampo e septo, que estão intimamente relacionadas na elaboração e expressão de funções cognitivas. Este padrão de conexões sugere que a amígdala exerce um papel de interface entre estímulos ambientais e as respostas à eles elaboradas. Ou, de outro modo, a amígdala fornece o "colorido" emocional aos aspectos sensoriais dos estímulos processados pelas estruturas hierarquicamente superiores do cérebro, como o cortex. Deste modo a influência da amígdala sobre a aprendizagem e memória é indissociável do seu papel na esfera emocional.
A elucidação dos mecanismos neurais subjacentes a estes fenômenos poderá resultar em aplicações a curto e médio prazo, que podem modificar substancialmente o tratamento dos distúrbios afetivos e de memória.
Muitos estudos em animais de laboratório têm documentado déficits de memória relacionados ao envelhecimento. Apesar de se observar o mesmo para humanos, ainda não se sabe se existe uma fundamentação neurobiológica comum subjacente a este declínio na memória observado em humanos e animais. Os resultados dessas pesquisas têm estimulado experimentos que investigam a possibilidade de que este déficit possa ser atenuado por tratamentos que usualmente melhoram a capacidade cognitiva em animais de laboratório. Epinefrina, por exemplo, quando administrada imediatamente após o treino aumenta a capacidade de memorização em ratos com idade avançada (Sternberg; Isaacs; Gold; McGaugh, 1985). Déficits de memória em ratos velhos podem também ser atenuados através de injeções de norepinefrina dentro dos ventrículos cerebrais (Collier et al., 1985). A administração da droga clonidina, que estimula os receptores adrenérgicos, antes do treinamento atenua os déficits de retenção observados em macacos idosos (Arnsten; Goldman-Rakic, 1985).
Em contraste com as evidências indicando que drogas podem melhorar a aprendizagem e memória em animais de laboratório, estudos dos efeitos de drogas sobre a memória em humanos têm apenas apresentado modestas evidências de resultados positivos. Existem muitas razões para isso. Primeiro, muitas das drogas usadas em experimentos com animais são, por vários motivos, inapropriadas para uso com humanos e por isso, não têm sido investigadas em estudos usando seres humanos. Segundo, estudos com humanos geralmente não usam os mesmos paradigmas experimentais comumente empregados nos experimentos com animais (por exemplo, a administração da droga concomitante à aquisição de uma informação específica). Terceiro, os tipos de memória testados em estudos com animais podem ser diferentes daqueles que são considerados como de grande importância no funcionamento cognitivo em humanos; isto é, os modelos animais podem ser inapropriados. Quarto, algumas drogas usadas em experimentos com humanos apresentam apenas efeitos modestos mesmo em experimentos com animais. Quinto, algumas drogas testadas em humanos produzem efeitos colaterais indesejáveis em doses menores do que aquelas necessárias para se observar uma melhora da memória.
Uma das escassas evidências positivas de substâncias que produzem melhora da memória em humanos advém dos estudos sobre drogas que afetam o sistema neurotransmissor colinérgico (Davies, 1985). Os resultados desses estudos são similares àqueles obtidos em animais: a capacidade cognitiva é melhorada por drogas que aumentam o funcionamento do sistema colinérgico. Experimentos recentes conduzidos por Gold e colaboradores (Gold, 1988) sugerem que, em ratos, injeções de glicose produzem uma melhora na memória semelhante àquela produzida pela epinefrina. Isto é interessante porque se sabe que a epinefrina ativa a liberação de glicose. Ademais, Gold e colegas (Hall; Gonder-Frederick; Chewning; Silveira; Gold, 1989) demonstraram que a glicose melhora a memória em um teste com sujeitos humanos saudáveis. Por certo esses resultados promissores irão estimular outras pesquisas sobre os efeitos da glicose no desempenho cognitivo em humanos.
É importante ressaltar que, a despeito das inúmeras pesquisas que vem sendo desenvolvidas em todo o mundo, ainda não se encontrou nenhuma droga (ou qualquer outro tratamento) cuja eficácia pudesse levar a um amplo uso no tratamento dos distúrbios de memória. Entretanto, esta conclusão pode mudar em breve como conseqüência dos resultados das pesquisas que estão sendo desenvolvidas. Progressos na neurobiologia molecular por certo irão provocar um grande impacto nesta área de pesquisa. A identificação de genes que regulam sistemas neurobiológicos específicos poderá levar à geneterapia ou técnicas para ativar ou desativar genes que estejam relacionados aos distúrbios de memória. A década de 90 foi considerada pelo Congresso Norte-Americano como a década do cérebro. Grandes investimentos estão sendo feitos nas neurociências. É possível que nos encontremos no limiar de uma nova fronteira do conhecimento científico e que o conhecimento das bases biológicas da memória se encontre a ponto de dar um salto qualitativo na compreensão dos fenômenos da psique humana.
REFERÊNCIAS BIBLIOGRÁFICAS
AMMASSARI-TEULE, M.; PAVONE, F.; CASTELLANO, C.; McGAUGH, J.L. Amygdala and dorsal hippocampus lesions block the effects of GABAergic drugs on memory storage. Brain Research, v.551, n.1/2, p.104-9, 1991. [ Links ]
ARNSTEN, A.F.T.; GOLDMAN-RAKIC, RS. Alpha2-adrenergic mechanisms in pre-frontal cortex associated with cognitive decline in aged nonhuman-primates. Science, v.230, n.4731, p. 1273-6, 1985. [ Links ]
BRIONI, J.D.; NAGAHARA, A.H.; McGAUGH, J.L. Involvement of the amygdala GABAergic system in the modulation of memory storage. Brain Research, v.487, n.1,p.105-12, 1989. [ Links ]
CASTELLANO, C. et al. GABAergic modulation of memory. In: SQUIRE, L.; LINDENLAUB, E., eds. Biology of memory. s.l, Schattauer Verlag, 1990. p.361-78. [ Links ]
COLLIER, G. et al. Impaired regulation of arousal in old age and the consequences for learning and memory: replacement of brain norepinephrine via neuron transplants improves memory performance in aged F344 rats. In: DAVIS, B.B.; WOOD, W.G., eds. Homeostatic function and aging. New York, Raven Press, 1985. p.99-110. [ Links ]
DAVIES, A. A critical review of the role of the cholinergic system in human memory and cognition. In: OLTON, D.S. et al., eds. Memory disfunctions: an integration of animal and human research from preclinical and clinical perspectives. New York, Academy of Sciences, 1985. p.212-7. [ Links ]
GOLD, P. Plasma glucose regulation of memory storage processes. In: WOODY, D.L. et al., eds. Cellular mechanisms of conditioning and behavioral plasticity. New York, Plenum Press, 1988. p.329-42. [ Links ]
GRAEFF, F.G. Brain defense systems and anxiety. In: BURROWS, G.D. et al., eds. Handbook of anxiety. Amsterdam, Elsevier, 1990. v.3, p.307-54. [ Links ]
HALL, J.L.; GONDER-FREDERICK, L.A.; CHEWNING, W.W.; SILVEIRA, J.; GOLD, P. Glucose enhancement of performance on memory tests in young and aged humans. Neuropsychologia, v.27, n.9, p. 1129-38, 1989. [ Links ]
IZQUIERDO, I.; MEDINA, J.H. Brain benzodiazepine/GABA A modulation of memory: the role of endogenous benzodjazepines. Trends in Pharmachological Sciences, v.12,p.260-5, 1991. [ Links ]
LeDOUX, J.E. Emotion and the amygdala. In: AGGLETON, J.P., ed. The amygdala. New York, Wiley-Liss, 1992. p.339-51. [ Links ]
LISTER, R. The amnestic action of benzodiazepines in man. Neuroscience and Biobehavioral Reviews, v.9, p.87-94, 1985. [ Links ]
McGAUGH, J.L. Involvement of hormonal and neuromodulatory systems in the regulation of memory storage. Annual Review of Neuroscience, v.12, p.255-87, 1989. [ Links ]
McGAUGH, J.L.; GOLD, P. Hormonal modulation of memory. In: BRUSH, R.B.; LEVINE, S., eds. Psychoneuroendocrinology. s.l., Academic Press, 1989. p.305-39. [ Links ]
NAGY, J.; ZÁMBÓ, K.; DECSI, L. Anti-anxiety action of diazepam after intra-amygdaloid application in the rat. Neuropharmacology, v.18, n.6, p.573-6, 1979. [ Links ]
OLTON, D.S.; WENK, P. Dementia: animal models of cognitive impairments produced by degeneration of the basal forebrain cholinergic system. In: MELTZER, H.Y., ed. Psychopharmacology: the third generation of progress. New York, Raven Press, 1987.p.941-53. [ Links ]
PARENT, M.B.; TOMAZ, C.; McGAUGH, J.L. Increased training in an aversively motivated task attenuates the memory impairing effects of posttraining N-methyl-D-aspartic acid-induced amygdala lesions. Behavioral Neuroscience, v.106, n.5, p.789-97, 1992. [ Links ]
SQUIRE, L.R. Memory and brain. Oxford, Oxford University Press, 1987. [ Links ]
STERNBERG, D.B.; ISAACS, K.R.; GOLD, P.E.; McGAUGH, J.L. Epinephrine facilitation of appetitive learning: attenuation with adrenergic receptor antagonists. Behavioral and Neural Biology, v.44, n.3, p.447-53, 1985. [ Links ]
TOMAZ, C. The role of the amygdala in learning and memory. Biotemas, v.5, n.2, p. 1-40, 1992. [ Links ]
TOMAZ, C.; AGUIAR, M.S.; NOGUEIRA, P.J. Facilitation of memory by peripheral administration of substance P and naloxone using avoidance and habituation learning tasks. Neuroscience and Biobehavioral Reviews, v.14, n.4, p.447-53, 1990. [ Links ]
TOMAZ, C.; DICKINSON-ANSON, H.; McGAUGH, J.L. Amygdala lesions block the amnestic effects of diazepam. Brain Research, v.568, n.1/2, p.85-91, 1991. [ Links ]
TOMAZ, C.; DICKINSON-ANSON, H.; McGAUGH, J.L. Basolateral amygdala lesions block diazepam-induced anterograde amnesia in an inhibitory avoidance task. Proceedings of the National Academy of Sciences of the United States of America, v.89, n.8, p.3615-9, 1992. [ Links ]
1 A preparação deste artigo foi financiada em parte pela FAPESP (90/3474-0) e pelo CNPq (300364/86-5).
