










Services on Demand

article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.20 no.2 Guadalajara 2012
ARTIGOS
Estudo exploratório sobre os efeitos do fechamento de suturas cranianas na aprendizagem discriminativa em ratos
Exploratory study on the effects of closuring cranial sutures on discriminative learning in rats
Nathalí Di Martino Sabino; Ana Beatriz Albino de Almeida; Vera Lúcia Adami Raposo do Amaral; Alexandre Shiobara; Gerson Yukio Tomanari1
Universidade de São Paulo (Brasil)
Sociedade Brasileira de Pesquisa e Assistência para Reabilitação Crânio-Facial (Brasil)
RESUMO
Intervenções no cérebro, tais como as provocadas por lesões mecânicas, podem afetar o comportamento dos organismos e sua capacidade de aprendizagem. O presente estudo analisou os efeitos do fechamento das suturas cranianas de ratos sobre a aprendizagem discriminativa. Serviram como sujeitos quatro ratos cirurgicamente intactos e nove submetidos a diferentes procedimentos cirúrgicos para o fechamento das suturas cranianas. Os sujeitos, sob privação controlada de água, foram submetidos a quatro fases experimentais. Na primeira fase, respostas de pressão à barra foram modeladas e reforçadas continuamente por três sessões consecutivas. Na segunda fase, empregou-se esquema de intervalo fixo (FI 4 s, FI 8 s e FI 11 s). Na terceira fase, iniciou-se um esquema múltiplo com componentes de reforçamento (FI 11 s) e Extinção, na presença e ausência de luz, respectivamente. Atingido ao menos 80% das resposta nos componentes de FI, as funções dos estímulos foram revertidas (quarta fase). De modo geral, os resultados revelaram que os procedimentos cirúrgicos não produziram efeitos evidentes sobre o responder dos animais nos diferentes esquemas, tampouco sobre o responder sob controle discriminativo (e sua reversão). A partir da metodologia inovadora apresentada nesse trabalho, investigações futuras poderão fazer uso de medidas adicionais (por exemplo, técnicas de neuroimagem) para avaliar, com mais precisão e controle, a forma e a extensão com que os procedimentos cirúrgicos afetaram (ou não) o desenvolvimento cerebral dos animais.
Palavras-chave: Aprendizagem discriminativa, esquema múltiplo, fechamento de suturas cranianas, resposta de pressão à barra, ratos.
ABSTRACT
Brain interventions, such as those produced by mechanical lesions, may affect organisms' behavior as well as their learning behavior. This study analyzed the effects of rat's cranial suture closing upon discriminative learning. Four surgical intact rats and nine rats that had different surgical procedures for the closing of cranial sutures were used as subjects. All subjects, under water-controlled deprivation, went through four experimental phases. In the first phase, lever responding was shaped and reinforced continuously for three consecutive sessions. In the second phase, fixed interval schedules were used (FI 4 s, FI 8 s, and FI 11 s). In the third phase, a multiple schedule composed of reinforcement (FI 11 s) and extinction, in the presence and absence of light respectively, was introduced. After 80% or more responses were emitted during FI components, stimulus functions were reverted (fourth phase). In general, results showed that surgical procedures did not have clear effects upon the animals' responding under the different schedules or under discriminative control (and its reversion). Based on the innovative methodology showed in this paper, future investigations will be able to use additional measures (e.g., neuroimage techniques) to evaluate, with more precision and control, the extend to which surgical procedures affected (or not) the development of the animals' brain.
Keywords: Discriminative learning, multiple schedule, cranial suture closure, bar press, rats.
Identificar relações entre determinados comportamentos e suas estruturas cerebrais correspondentes tem sido objeto de grande interesse científico. Inúmeras pesquisas ilustram historicamente esse interesse: a demonstração de que o neocórtex é essencial para o condicionamento de respostas reflexas de cães (Pavlov, 1984) é uma delas. Um outro exemplo são as evidências de que o processo de aprendizagem operante mantém-se intacto ainda que estejam lesionadas as estruturas telencefálicas de ratos (Huston & Tomás, 1986; Huston, Tomás, & Fix, 1985) e aves (Cerutti, Cintra, Diáz-Cintra, & Ferrari, 1997; Cerutti & Ferrari, 1995; de Souza, Brito & Ferrari, 1990).
Procedimentos de discriminação, nos quais comportamentos são emitidos sob controle de estímulos (por exemplo, o responder em esquema múltiplo), têm se mostrado especialmente úteis para a análise da aprendizagem discriminativa em sujeitos não humanos submetidos a lesões do sistema nervoso central. Esses estudos propiciam tanto um maior conhecimento fisiológico do órgão afetado por alguma patologia ou lesão quanto propiciam a compreensão de seus efeitos sobre o comportamento do organismo (Bitterman, 1965; Cerutti et al., 1997; Cerutti & Ferrari, 1995; Lam, Sethi, & Low, 2005; Lashley, 1929; Mazarakis et al., 2005; Mitchell, Brabano, Losken, Siegel, & Mooney, 2003; Stettner & Schultz, 1967).
Na análise do comportamento operante, Lashley (1929) realizou um estudo pioneiro ao verificar os efeitos da destruição mecânica de partes do cérebro sobre o responder discriminado de ratos adultos. Para isso, utilizou como equipamento um labirinto composto de dois corredores que podiam ser iluminados alternadamente. Em um primeiro experimento, quando o sujeito seguia pelo corredor iluminado, alcançava uma porta que o levava a um compartimento em que havia comida. Quando seguia pelo corredor não iluminado, era exposto a um choque elétrico. Os corredores eram iluminados randomicamente nas 100 tentativas de cada sessão. O experimento era encerrado quando o sujeito percorria o corredor iluminado por 30 tentativas consecutivas.
Após a aquisição da resposta discriminada, os sujeitos passaram por uma cirurgia destinada à destruição de porções variáveis dos lobos occipitais dos dois hemisférios cerebrais. Após a recuperação, os sujeitos eram novamente expostos à tarefa experimental. Nessa condição pós-cirúrgica, todos os sujeitos deixaram de emitir a resposta discriminada. Entretanto, no re-treino, os sujeitos voltaram a se comportar sob controle discriminativo e, curiosamente, o número de repetições necessário para atingir o critério de encerramento mostrou-se proporcional à extensão da parte inutilizada do lobo occipital a despeito da localização exata da lesão.
Em um segundo experimento, com um novo grupo de ratos, Lashley (1929) realizou o procedimento cirúrgico para a destruição de um terço do lobo occipital antes do início da atividade experimental. Apesar da lesão, não foi observada diferença na aquisição da discriminação desses sujeitos em relação aos sujeitos intactos, o que levou o autor a sugerir que a discriminação da luminosidade não dependeria de uma região específica do lobo occiptal.
Em um terceiro experimento, Lashley (1929) inutilizou cirurgicamente áreas cerebrais parietais em um grupo de ratos que já havia aprendido a responder discriminadamente. O resultado indicou que esses sujeitos conseguiam responder sob controle dos estímulos luminosos, o que indicava que a região parietal não seria fundamental para o desempenho bem-sucedido em uma tarefa discriminativa.
Para ampliar a análise da aprendizagem discriminativa, um importante aspecto a ser considerado são as alterações comportamentais que acompanham a reversão das funções dos estímulos. Nesta, por exemplo, o estímulo discriminativo de alta probabilidade de reforçamento passa a atuar como estímulo discriminativo de baixa ou nula probabilidade de reforçamento, e vice-versa. Com isso, em sujeitos intactos, o responder diferencial que resultava da discriminação previamente estabelecida é perdida e, na sequência, passa a se constituir um responder diferencial compatível com os novos estímulos discriminativos. Em estudos que investigam o processo de aprendizagem discriminativa (ou variáveis que a afetam, como é o caso do presente trabalho), a adaptação do responder diante da reversão fortalece a demonstração do controle exercido pelos estímulos (pré- e pós-reversão), bem como pelas contingências responsáveis pela nova discriminação (Matos, 1981).
Sob essa perspectiva, Bitterman (1965) analisou não apenas a aquisição de uma discriminação, mas também as suas reversões, em ratos submetidos à destruição cirúrgica da região cortical do cérebro. A tarefa consistia em uma discriminação visual sucessiva de dois painéis luminosos, na qual apenas respostas de pressão a um deles produzia o reforçador. Em cada sessão, o sujeito era exposto a 20 tentativas. Caso ocorressem pelo menos 17 acertos em 20 tentativas, procedia-se a reversão da discriminação. Os resultados mostraram que, considerando-se as sessões iniciais, a aquisição da discriminação ocorreu de forma mais lenta para os animais submetidos à cirurgia do que para os animais intactos. Entretanto, consideradas as últimas sessões, diferenças marcantes entre os grupos de sujeitos deixaram de existir. Ou seja, apesar de terem a região cortical cirurgicamente lesionada, os ratos cirurgicamente lesionados respondiam discriminadamente a estímulos visuais e, sobretudo, na medida em que passavam por uma série de reversões, deficiências observadas inicialmente na aprendizagem discriminativa tendiam a desaparecer.
Em síntese, modelos animais têm contribuído para a análise de comportamentos que parecem se relacionar funcionalmente com determinadas regiões identificadas no cérebro por meio de manipulações cirúrgicas. Modelos como esses são de grande importância para os estudos de patologias que também acarretam alterações no cérebro. Por exemplo, a cranioestenose, uma dessas patologias, caracteriza-se pelo fechamento precoce das suturas cranianas que leva a alterações anatômicas no cérebro e, potencialmente, pode produzir efeitos comportamentais (Mitchell et al., 2003).
Lam et al. (2005) estudaram os efeitos decorrências do fechamento precoce das suturas cranianas sobre respostas motoras de ratos. Nesse estudo, foram utilizados 56 ratos que tiveram suas suturas fechadas por meio do uso do methyl 2-Cyanocrylate quando tinham zero, três e sete dias de vida. Para controle experimental, outros 56 sujeitos com mesma idade não sofreram intervenção cirúrgica. Os testes comportamentais empregaram medidas em campo aberto, isto é, uma caixa de plástico translúcida de 40 cm² de base. Os sujeitos eram colocados na caixa, quando completavam 30 dias de vida, por três minutos. Como medida de atividade locomotora, foi mensurado a distância linear total que cada sujeito percorria na caixa. Os resultados mostraram que os sujeitos cirurgicamente manipulados, diferentemente dos demais, percorriam distâncias maiores, o que levou os autores a sugerirem que o grau maior de atividade locomotora seria um possível efeito do fechamento precoce das suturas do cérebro.
No estudo de Lam et al. (2005), os efeitos do fechamento precoce das suturas cranianas foram analisados sobre a resposta de percorrer um campo aberto. Sob o ponto de vista da análise do comportamento, trata-se de um comportamento cuja natureza operante encontra-se, muito provavelmente, mesclada a comportamentos típicos da espécie - por exemplo, comportamentos exploratórios. Por essa razão, avanços importantes nessa linha de pesquisa seriam: analisar os efeitos do fechamento precoce das suturas cranianas sobre a aprendizagem de uma resposta com características (1) fundamentalmente operante e (2) sob controle discriminativo. Considerando-se uma resposta operante largamente empregada em estudos experimentais com ratos, ou seja, a pressão à barra, estariam as discriminações e suas reversões preservadas em ratos cujas suturas do crânio fossem precocemente fechadas cirurgicamente?
Visando fornecer subsídios empíricos que contribuam para responder a essa pergunta, o presente estudo investigou, em ratos expostos ao fechamento cirúrgico precoce das suturas cerebrais, a aquisição da resposta de pressão à barra sob controle discriminativo, seguida pela reversão da discriminação.
MÉTODO
Sujeitos
Foram utilizados 13 ratos Wistar, 9 machos e 4 fêmeas, experimentalmente ingênuos, provenientes do Centro Multidisciplinar para Investigação Biológica da Universidade de Campinas (CEMIB/UNICAMP).
Dos 13 sujeitos, nove (E, F, G, H, I, J, K, L e M) foram submetidos a uma cirurgia de indução ao fechamento das suturas cranianas (Condição Cirúrgica) e quatro (A, B, C e D) não foram submetidos a nenhum procedimento cirúrgico (Condição Não-Cirúrgica).
Equipamentos
Foram utilizadas duas caixas de condicionamento operante modulares, modelo ENV-008VP, fabricadas por Med Associates Inc. As caixas são equipadas com um bebedouro localizado no centro da parede direita. À direita do bebedouro encontra-se uma barra de respostas. Em repouso, o bebedouro é mantido mergulhado numa cuba com água localizada na parte externa da caixa. A cada acionamento do bebedouro, por meio de uma resposta de pressão à barra, uma concha com cerca de 0,06 ml de líquido emerge da cuba d'água e fica acessível no interior da caixa operante por 4 s. Na parede direita da caixa, na parte superior, há uma lâmpada de 40 Watts.
O registro das respostas de pressão à barra e a programação das contingências experimentais foram controladas pelo aplicativo MedPC – Schedule Manager.
Procedimento
Os 13 sujeitos foram submetidos a quatro fases experimentais: 1) modelagem de pressão à barra e reforçamento contínuo (CRF); 2) treino do responder sob esquema de Intervalo Fixo (FI); 3) treino discriminativo em esquema múltiplo; e 4) reversão da discriminação. As quatro fases encontram-se detalhadas a seguir e sintetizadas na Tabela 1.

Fase Pré-Experimental - Procedimento Cirúrgico
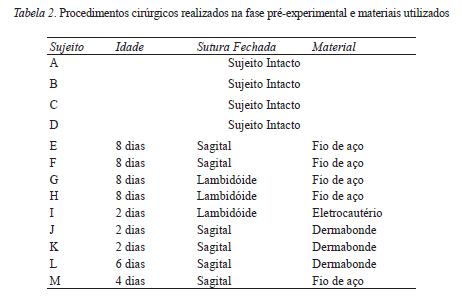
Os nove sujeitos da Condição Cirúrgica passaram por uma fase pré-experimental. Nesta, a equipe cirúrgica do Hospital SOBRAPAR (Sociedade Brasileira de Pesquisa e Assistência para Reabilitação Craniofacial), por meio de diferentes procedimentos e materiais exploratórios2 (ver descrição a seguir), buscou produzir o fechamento das suturas cranianas que levaria à constrição no crescimento cerebral desses animais.
Os sujeitos E, F, G e H foram submetidos ao procedimento cirúrgico com 8 dias de vida. Na cirurgia, os sujeitos E e F tiveram a sutura sagital fechada com um fio de aço. O sujeito G teve as placas parietais unidas com a placa occipital por meio de fio de aço. O sujeito H teve a placa occipital unida com a parietal esquerda com a utilização de fio de aço.
O sujeito I foi submetido ao procedimento cirúrgico com 2 dias de vida e teve as placas parietais unidas com a occipital por meio de eletrocautério. Com 2 dias de vida, os sujeitos J e K tiveram unidas suas placas parietais com aplicação de dermabonde após uma incisão na sutura sagital e raspagem do periósteo. O sujeito L, com 6 dias de vida, teve uma incisão na sutura sagital, cauterização das margens da placa parietal e aplicação de um adesivo de colagem instantânea. Com 4 dias de vida, o sujeito M teve uma incisão na sutura sagital e as placas parietais foram unidas com fio de aço. A Tabela 2 sumariza os procedimentos cirúrgicos realizados nesta fase pré-experimental e os materiais utilizados nesses procedimentos.
Todos os sujeitos experimentais foram anestesiados com Xilasina3 (dose intramuscular) e Quetamina4 (dose intramuscular). Após a cirurgia, os animais permaneceram por 20 min sob luz incandescente para restabelecer a temperatura corpórea, pois, durante o procedimento cirúrgico, os mesmos permaneceram afastados do calor materno e seus corpos resfriaram-se. Na sequência, foram recolocados junto à genitora na caixa viveiro. Foram misturados à maravalha (material utilizado para forrar a gaiola) pré-existente no interior da caixa para retirar o odor da manipulação e do procedimento cirúrgico. Alguns animais foram submetidos, no momento do ato cirúrgico, à dermabonde ou adesivo de colagem instantânea. Para que as mães se adaptassem ao odor dessas substâncias e, com isso, não rejeitassem os filhotes após a cirurgia, essas substâncias foram gotejadas nos cantos das gaiolas antes do retorno dos mesmos à caixa.
Transcorridos cinco dias após a cirurgia, os animais passaram a ser pesados, diariamente, até o final do experimento. Vinte e cinco dias após o início da pesagem, os animais foram alojados em gaiolas individuais e tiveram, até o oitavo mês, acesso livre à alimentação e à água, com vistas à completa recuperação do procedimento cirúrgico.
Com oito meses de vida, ocasião em que já se encontravam plenamente recuperados da cirurgia, todos os animais, de ambos os grupos, foram mantidos sob condição de acesso restrito à água de modo a que seus pesos permanecessem em cerca de 85% do peso ad libitum. Esses valores foram calculados a partir da média de consumo em ad libitum durante os últimos 5 dias de pesagem (cf. Tomanari et al., 2007; Tomanari, Pine, & Silva, 2003).
Fase 1 – Modelagem de Pressão à Barra e Reforçamento Contínuo (CRF)
Os 13 sujeitos passaram por um treino preliminar que consistiu de uma sessão de modelagem na qual a resposta de pressionar a barra foi reforçada diferencialmente por aproximações sucessivas. Instalada a resposta, os sujeitos foram expostos a três sessões de CRF, com 60 reforçadores cada, objetivando fortalecer a resposta anteriormente modelada. A luz ambiente da caixa experimental permaneceu acesa no decorrer de todas as sessões dessa fase.
Fase 2 – Treino do Responder sob Esquema de Intervalo Fixo (FI)
Nessa fase, foi realizada uma sessão de 60 min em esquema FI 4 s, uma sessão de 60 min de FI 8 s e, finalmente, duas sessões de 60 min cada de FI 11 s. A luz ambiente da caixa experimental permaneceu acesa no decorrer de todas as sessões dessa fase.
Fase 3 – Treino Discriminativo em Esquema Múltiplo
Nessa fase, para colocar as respostas de pressão à barra sob controle discriminativo, os animais foram expostos a um esquema múltiplo FI 11 s EXT com componentes de 1 min. Durante o componente de FI 11 s, a luz da caixa era mantida acesa (S+) e, durante o componente de extinção, a luz permanecia apagada (S-). Cada um dos componentes foi apresentado 30 vezes por sessão, constituindo sessões de 60 min de duração. A ordem de apresentação dos componentes era semi-randômica, com repetição de cada componente em, no máximo, duas vezes consecutivas (ou seja, 2 min no mesmo componente). Esta fase foi encerrada quando o sujeito atingia três sessões consecutivas em que o Índice Discriminativo (ID = razão entre a taxa de respostas em S+ e a taxa total de respostas) era maior ou igual a 80% em cada sessão.
Fase 4 – Reversão da Discriminação
Após atingirem o critério de encerramento da Fase 3 (três sessões consecutivas com, no mínimo, 80% das respostas emitidas diante do S+), os animais foram expostos à reversão das funções dos estímulos discriminativos, sem alterações adicionais nos outros parâmetros do treino e no critério de encerramento da fase. Nesse caso, o componente extinção passou a ser acompanhado pela presença da luz, e o componente FI 11 s, pela ausência da luz.
RESULTADOS
A avaliação do desempenho dos sujeitos nas Condições Não-Cirúrgica (sujeitos A, B, C e D) e Cirúrgica (sujeitos E, F, G, H, I, J, K, L e M) foi realizada por meio da análise da taxa de respostas por minuto (resp/min) nas Fases 1 e 2, assim como pelo Índice Discriminativo (ID) nas Fases 3 e 4. Além disso, foi analisado o número de sessões nas Fases 3 e 4 para todos os sujeitos, em ambas as condições, Cirúrgica e Não-Cirúrgica.
As Figuras 1 e 2 fornecem uma análise geral dos dados ao mostrar as taxas de pressão à barra na Condição Não-Cirúrgica (Figura 1) e na Condição Cirúrgica (Figura 2), Fases 1 e 2.
Comparando as duas figuras, é possível notar que, de modo geral, os diferentes esquemas de reforço empregados ao longo das duas fases geraram efeitos sobre as taxas de respostas que, aparentemente, independeram da exposição prévia à cirurgia. De modo geral, verifica-se uma relativa variação de desempenho entre os sujeitos. Por exemplo, o responder sob reforçamento contínuo produziu taxas de respostas relativamente maiores do que as obtidas, na sequência, sob o responder em FI 4 s (sujeitos A, C, F, G, H e J). Todavia, nem sempre a mudança de reforçamento contínuo para FI 4 s gerou diminuição na taxa de respostas. Por vezes, houve aumento (sujeitos B, D, E); por vezes, houve manutenção (sujeitos I e K, L e M).
Com o aumento do intervalo do FI de 4 a 8 segundos, o efeito mais preponderante foi o aumento na taxa de respostas (sujeitos A, C, D, G, H, I, J, L, M). Pontualmente, as taxas permaneceram praticamente constantes para o sujeito B e diminuíram para os sujeitos E, F, e K.
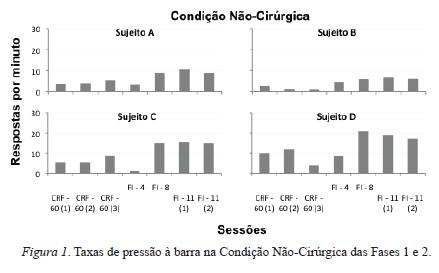
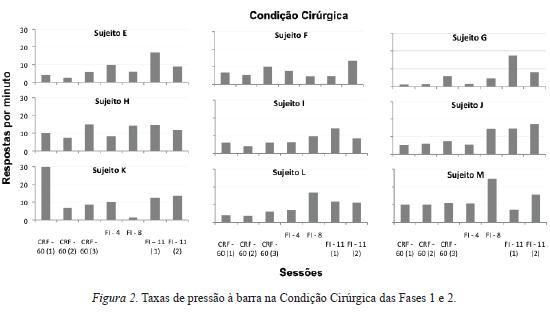
Com a mudança do FI 8 s para FI 11 s, praticamente não houve alterações nas taxas de respostas (sujeitos A, B, C, F, H, J). Porém, houve também uma leve redução para alguns animais (sujeitos D, L, M) e aumento para outros (sujeitos E, G, I, K).
As Figuras 3 e 4 apresentam o ID nas sessões que compuseram as Condições Não-Cirúrgica (Figura 3) e Cirúrgica (Figura 4) das Fases 3 e 4. A linha vertical tracejada sinaliza a mudança da Fase 3 para a Fase 4.

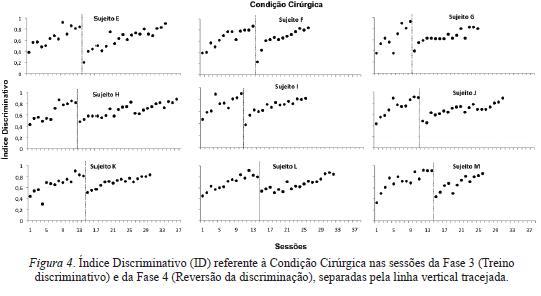
O ID é uma medida da discriminação entre os componentes do esquema múltiplo. De modo geral, para todos os sujeitos, em ambas as fases e nas duas condições experimentais, houve uma aquisição gradual da discriminação. Inicialmente, o ID era próximo de 0,5, o que representa frequências não diferenciadas entre os componentes de FI e extinção. No decorrer das sessões, entretanto, os IDs tenderam a aumentar, até atingirem valores próximo a 1,0, o que indica que praticamente todas as pressões à barra eram emitidas durante os componentes de FI.
A Figura 5 mostra o número de sessões realizadas até que os sujeitos concentrassem ao menos 80% das respostas nos componentes de FI (isto é, atingissem ID igual ou superior a 0,8) no treino discriminativo (Fase 3) e na reversão da discriminação (Fase 4), para ambas as condições experimentais, Cirúrgica e Não- Cirúrgica. Nessa fi gura, cada barra vertical refere-se a um sujeito. Os sujeitos estão ordenados em ordem crescente de acordo com o número total de sessões.
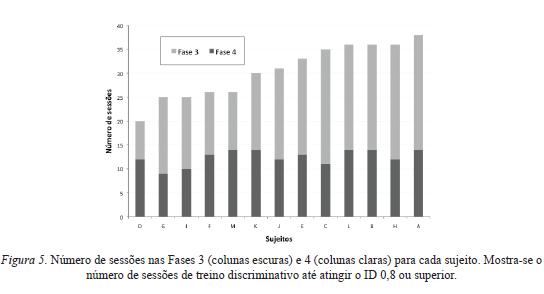
No que diz respeito à Fase 3 (porções escuras das barras verticais), os sujeitos atingiram IDs de 0,8 em torno de 9 a 14 sessões, sem qualquer aparente distinção entre os sujeitos das Condições Cirúrgica e Não-Cirúrgica.
No que diz respeito à Fase 4, na qual ocorreu a reversão da discriminação (porções claras das barras verticais), houve uma maior variação entre os sujeitos com relação ao número de sessões realizadas até que o ID fosse igual ou superior a 0,8. Por exemplo, o sujeito D passou por 8 sessões (o menor número registrado entre todos animais), enquanto que o sujeito C passou por 24 sessões (o maior número registrado entre todos animais).
Ao que parece, os dados não mostram uma relação entre o número de sessões realizadas no treino discriminativo e na reversão. Isto é, não indicam possibilidades preditivas da duração da reversão a partir da duração do treino discriminativo imediatamente anterior.
Finalmente, deve-se destacar o fato de que o número total de sessões por parte de três dos quatro sujeitos que não passaram por cirurgia encontra-se entre os maiores entre todos os animais (sujeitos A, B e C). Por outro lado, o sujeito que passou pelo menor número de sessões foi, também, um sujeito que não passou por cirurgia (sujeito D).
DISCUSSÃO
Os resultados do presente estudo mostraram padrões de respostas operantes muito semelhantes entre os animais expostos à cirurgia e os animais intactos. Pelo que se verifi cou, não houve diferenças comportamentais nos ratos que tiveram o fechamento das suturas cranianas em relação aos animais intactos, ao menos nas seguintes condições: 1) quando expostos a esquemas de reforçamento contínuo e a esquemas de reforçamento intermitente (FI) da resposta de pressão à barra mantida pela produção de água; 2) quando expostos a esquema múltiplo constituído por componentes de reforçamento e extinção sob controle de presença e ausência de luz; e 3) quando expostos à reversão da discriminação previamente treinada.
Embora estudos da literatura apresentem variações nas metodologias utilizadas e digam respeito a lesões de partes cerebrais diversas, estes costumam descrever uma bem-sucedida instalação de respostas operantes sob controle discriminativo em diferentes sujeitos (Bechara, Damasio, Damasio, & Anderson, 1994; Bitterman, 1965; Jackson, Jackson, Harrison, Hederson, & Kennard, 1995; Knopman & Nissen, 1991; Lam et al., 2005; Lashley, 1929; Schulund, 2002; Schulund & Pace, 2000; Stettner & Schultz, 1967; Swenson & Butters, 1996). Uma exceção é o trabalho de Mazarakis et al. (2005). Em camundongos usados como modelo animal da doença de Huntington, doença neurodegenerativa que afeta a coordenação dos músculos e leva à demência, esses autores constataram dificuldades em tarefas comportamentais que envolviam aprendizagem discriminativa.
No que diz respeito particularmente à aquisição de repertórios discriminativos em ratos submetidos à fusão precoce das suturas cranianas, a literatura é escassa. Conforme relatado anteriormente, Lam et al., (2005) abordaram a questão, porém limitaram-se a avaliar somente o comportamento de caminhar, ou seja, apenas o comportamento motor dos ratos. Portanto, uma das contribuições do presente estudo é sua proposta metodológica, na medida em que oferece um contexto experimental que permite analisar os possíveis efeitos do fechamento precoce das suturas cranianas sobre um comportamento operante, controlado por suas conseqüências, bem como por estímulos discriminativos. Nessa direção, abrem-se valiosas oportunidades para que sejam estudados parâmetros da relação resposta-consequência (e.g., por meio dos esquemas de razão e intervalo) e o controle da resposta por estímulos antecedentes (e.g., por meio de discriminações simples e condicionais).
Com relação aos resultados, o presente estudo mostrou que o desempenho dos ratos que passaram pela cirurgia, nos esquemas simples, foi muito semelhante aos que não passaram. Essa similaridade sugere que a fusão das suturas cranianas não tenha tido impacto sobre as pressões à barra, seja em razão de comprometimento motor ou da idade do organismo. Ao iniciar a tarefa na caixa operante, os animais estavam com 8 meses de idade, ou seja, eram ratos adultos. Conforme afirmado anteriormente, não há estudos que tenham investigado os efeitos do fechamento precoce de suturas cranianas sobre o comportamento operante de ratos. Portanto, sem poder contar com dados da literatura, o papel investigatório da presente pesquisa nos levou a considerar razoável supor que, aos 8 meses de idade, os animais, recuperados da cirurgia, demonstrassem eventuais efeitos cirurgia em razão do crescimento total do cérebro. É possível que os efeitos surgissem em momentos anteriores ou posteriores do desenvolvimento físico do animal. No entanto, para responder conclusivamente a essa questão, pesquisas futuras precisarão manipular essa variável diretamente.
Os resultados mostraram ainda que a fusão das suturas cranianas não impediu a aquisição do responder discriminado, ao menos nas situações contrastantes de luz acesa e luz apagada. Possivelmente, estímulos de outra natureza (e.g., auditiva ou tátil), ou mesmo estímulos luminosos menos discrepantes entre si (e.g., luz forte vs. luz fraca no lugar de luz acesa vs. luz apagada), poderiam provocar efeitos distintos na aquisição da discriminação em função do fechamento (ou não) das suturas cranianas. De qualquer modo, tal como foi conduzido o presente experimento, os animais submetidos à cirurgia não precisaram de maior ou menor número de sessões de treino que os demais para adquirir a discriminação. Ao contrário, um treino mais longo foi necessário para a instalação do responder discriminado em pombos destelencefalados nos estudos de Cerutti et al. (1997) e Cerutti e Ferrari (1995).
Diferentemente do que foi observado por Lam et al., (2005), em ratos, e Mitchell et al. (2003), em coelhos, os dados do presente estudo mostraram variações mais marcantes entre os sujeitos, ao menos no que diz respeito ao desempenho nos esquemas simples (CRF e FI) e no número de sessões conduzidas na reversão da discriminação. Embora Lam et al. (2005) não tenham apresentado a informação de qual foi a sutura fechada em cada um dos ratos, é possível, em termos ainda hipotéticos, que a diferença no fechamento precoce de diferentes suturas possa explicar as variações comportamentais que obtivemos. Entretanto, conforme afirmado anteriormente, estudos a esse respeito são escassos, de modo que um nível de controle experimental que permita investigar os efeitos de cada sutura em particular ainda não está disponível. A partir da continuação de trabalhos como o presente, um modelo animal poderá se constituir e, assim, permitir que os efeitos de cada sutura, em particular, possam ser identificados com alguma precisão.
Do que parece possível aventar a partir do presente estudo, é bem possível que o fechamento das suturas cranianas nos ratos não tenha afetado a região frontal do cérebro ou o estriado. Somos levados a essa suposição com base na comparação com os estudos de Bechara et al. (1994), Jackson et al. (1995) Knopman e Nissen (1991), Schulund (2002), Schulund e Pace (2000) e Swenson e Butters (1996), nos quais as lesões encontradas na região frontal ou o estriado prejudicariam a aprendizagem discriminativa, fato não constatado no desempenho dos nossos sujeitos. Em vista disso, é importante que estudos futuros verifiquem in loco os reais efeitos do fechamento das suturas cranianas sobre o cérebro de cada animal. Com esses dados, possíveis relações entre as áreas afetadas e as medidas comportamentais seriam estabelecidas com maior precisão. Além disso, para aproximar ainda mais o presente procedimento a um possível modelo animal de cranioestenose, técnicas que permitissem o fechamento da sutura craniana antes do nascimento dos animais seriam bem-vindas (Lam, et al., 2005), ainda que se possam reproduzir efeitos análogos quando se realiza a cirurgia logo após o nascimento (Shen et al., 2009).
REFERÊNCIAS
Bechara, A., Damasio, A. R., Damasio, H., & Anderson, S. W. (1994). Insensitivity to future consequences following damage to human prefrontal córtex. Cognition, 50, 7-15. [ Links ]
Bitterman, M. E. (1965). Philetic differences in learning. American Psychologist, 20, 396-410. [ Links ]
Cerutti, S. M., Cintra, L., Diáz-Cintra, S., & Ferrari, E. A. M. (1997). Behavioural and neuroanatomical correlates of long-term detelencephalation in pigeons. Journal of Physiology and Biochemistry, 53, 104. [ Links ]
Cerutti, S. M., & Ferrari, E. A. M. (1995). Funções da neuroglia no sistema nervoso central: contribuições para a plasticidade neural. LECTA: Revista de Farmácia e Biologia, 13, 79-100. [ Links ]
de Souza, C. M. E., Brito, L. R. E., & Ferrari, E. A. M. (1990). Key pecking operant conditioning in detelencephaled pigeons. Behavioral Brain Research, 38, 223-231. [ Links ]
Huston, J. P., & Tomás, C. (1986). Subtelencephalic location of reinforcement and learning: Looking for the minimal necessary structures. Behavioral Brain Research, 22, 153-161. [ Links ]
Huston, J. P., Tomás, C., & Fix, T. (1985). Avoidance learning in rats devoid of telencephalon plus thalamus. Behavioral Brain Research, 17, 87- 99. [ Links ]
Jackson, G. M., Jackson, S. R., Harrison, J., Henderson, L., & Kennard, C. (1995). Serial reactions time learning and Parkisnson's disease: Evidence for a procedural learning deficit. Neuropsychologia, 33, 577-593. [ Links ]
Knopman, D., & Nissen, M. J. (1991). Procedural learning is impaired in Huntington's disease: Evidence from the serial reaction time task. Neuropsychologia, 29, 245-254. [ Links ]
Lam, C. H., Sethi, K. A., & Low, W. C. (2005). A morphometric, neuroanatomical, and behavioral study on the effects of geometric constraint on the growing brain: The methyl 2-cyanocrylate craniosynostosis model. Journal of Neurosurgery Pediatrics, 102, 396-402. [ Links ]
Lashley, K. S. (1929). Brain mechanisms and intelligence. Chicago: University of Chicago Press. [ Links ]
Matos, M. A. (1981). O controle de estímulo sobre o comportamento. Psicologia, 7, 1-15. [ Links ]
Mazarakis, N., Cybulska-Klosowicz, A., Grote, H., Pang, T., Van Dellen, A., Kossut, M., Blakemore, C., & Hannan, A. J. (2005). Deficits in experience-dependent cortical plasticity and sensory-discrimination learning in presymptomatic Huntington's disease mice. The Journal of Neuroscience, 25, 3059-3066. [ Links ]
Mitchell, R. L., Barbano, T. E., Losken, H. W., Siegel, M. I., & Mooney, M. P. (2003). Early neuromotor behavior in craniosynostotic rabbits. Cleft Palate-Craniofacial Journal, 40, 486-492. [ Links ]
Pavlov, I. P. (1984). O Trabalho dos Grandes Hemisférios Cerebrais (R. Moreno, Trad.). Em Os pensadores (pp. 85-98). São Paulo: Abril. (Trabalho original publicado em 1926) [ Links ]
Schlund, M. W. (2002). Effects of acquired brain injury on adaptative choice and the role of reduced sensitivity to contingencies. Brain Injury, 16, 527-535. [ Links ]
Schlund, M. W., & Pace, G. (2000). The effects of traumatic brain injury on reporting and responding to causal relations: An investigation of sensitivity to reinforcement contingencies. Brain Injury, 14, 573-583. [ Links ]
Shen, K., Krakora, S. M., Cunningham, M., Singh, M., Wang, X., Hu, F. Z., Post, J. C., & Ehrlich, G. D. (2009). Medical treatment of craniosynostosis: Recombinant Noggin inhibits coronal suture closure in the rat craniosynostosis model. Orthodontics & Craniofacial Research, 12, 254–262.
Stettner, L. J., & Schultz, W. J. (1967). Brain lesions in birds: effects on discrimination acquisition and reversal. Science, 155, 3770. [ Links ]
Swenson, M., & Butters, N. (1996). Dissociations within nondeclarative memory in Huntington's disease. Neuropsychology, 10, 538-548. [ Links ]
Tomanari, G. Y., Amaral, V. L. A. R., Ramos, K. P., Sabino, N. M., Geremias, M. C. G., Sanches, L. C., & Albino, A. B. (2007). Parâmetros generalizados de operações de privação por restrições hídrica e alimentar em ratos machos Wistar. Acta Comportamentalia, 15, 131-145. [ Links ]
Tomanari, G. Y., Pine, A. S., & Silva, M. T. A. (2003). Ratos Wistar sob regimes rotineiros de privação hídrica e alimentar. Revista Brasileira de Terapia Comportamental e Cognitiva, 5, 57-71. [ Links ]
Received: February, 25, 2011
Accepted: August, 24, 2011
1) Os autores agradecem ao doutorando Arturo Clavijo e ao Pós-Doutorando Saulo M. Velasco, ambos da Universidade de São Paulo, por suas contribuições à versão final desse artigo. O trabalho recebeu apoio do CNPq na forma de bolsa de mestrado à Nathalí Sabino e bolsa de Produtividade em Pesquisa ao Dr. Gerson Y. Tomanari. Correspondências podem ser enviadas a Gerson Yukio Tomanari, Departamento de Psicologia Experimental, Universidade de São Paulo. Av. Prof. Mello Moraes, 1721, São Paulo, SP 05508-030, Brasil. E-mail: tomanari@usp.br
2) Pelo fato de se tratarem de intervenções cirúrgicas inéditas, ainda em desenvolvimento, os procedimentos empregados na indução do fechamento precoce das suturas cranianas foram realizados de diversas maneiras e com materiais distintos para futuras análises e avaliações no âmbito do interesse médico-cirúrgico. No presente artigo, cujo interesse é analisar variáveis comportamentais, os aspectos técnicos dos procedimentos e métodos cirúrgicos não serão detalhados.
3) Agonista alfa adrenérgico usado como sedativo, analgésico e relaxante muscular.
4) Derivado cicloexanona utilizado para a indução de anestesia.
