










Services on Demand

article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkUniversitas Psychologica
Print version ISSN 1657-9267
Univ. Psychol. vol.7 no.2 Bogotá Aug. 2008
Análisis de la variación en la disponibilidad en la entrega de agua bajo distintas condiciones de señalización de los subciclos en un programa temporal
Analysis of variation in the availability of water delivery under different signaling conditions of subcycles in a time schedule
Carlos de Jesús Torres-Ceja*; Alfredo Mayoral; Emilio Ribes-Iñesta
Universidad de Guadalajara, México
ABSTRACT
This study was conducted to evaluate the effects of the variations of water availability through the manipulation of T values, under different signaling conditions of subcycles, maintaining constant the water delivery probability (P). Data from this study show that in some rats were observed an increase in the frequency response associated with varying T. There were no systematic effects related to signals in subcycles. We analyze the functional difference between P and T and its relationship with the frequency and temporal distribution of response.
Key words authors Temporary distribution of response, Time schedules, Rats.
Key words plus Water-análisis, Laboratory research, Animals, Laboratory.
RESUMEN
Se diseñó un estudio con el objetivo de evaluar los efectos de variar la disponibilidad en la entrega de agua a través de la manipulación de los valores de T, bajo distintas condiciones de señalización de los subciclos en un programa temporal, manteniendo constante el valor la probabilidad (P) de entrega del agua. Los datos de este estudio muestran que en algunas ratas se observó un incremento en la frecuencia de respuesta asociada a la variación de T. No se observaron efectos sistemáticos relacionados con las señales en los subciclos. Se analiza la diferencia funcional entre P y T y su relación con la frecuencia y la distribución temporal de respuesta.
Palabras clave autores Distribución temporal de la respuesta, Programas temporales, Ratas.
Palabras clave descriptores Agua-análisis, Investigación de laboratorio, Animales de laboratorio.
Los programas temporales (Schoenfeld, Cumming & Hearst, 1956; Schoenfeld & Cole, 1972) permiten la manipulación de manera independiente de dos parámetros asociados con la presentación de estímulos: a) la probabilidad de ocurrencia en un ciclo T determinado relativo al número total de ciclos T programados (P) y b) la disponibilidad temporal dentro de cada ciclo T (T). P se define como la probabilidad de entrega o presentación de un estímulo ante la primera respuesta en el subciclo y su valor puede variar en el rango de valores de 1.0 a 0.0. La distribución de P de los subciclos es aleatoria a lo largo de la sesión experimental y, consecuentemente, no se puede anticipar en cuál ciclo esté disponible el estímulo. Por su parte, T se define como la proporción del tiempo discriminativo relativo al valor del ciclo T ó tD / (tD + t). El tiempo discriminativo (tD) hace referencia a la porción del ciclo T en que la disponibilidad para la ocurrencia de un estímulo es mayor que en el resto del ciclo (t). En los programas temporales la disponibilidad para la ocurrencia de un estímulo, se restringe sólo a una respuesta dentro del ciclo T cuando el programa es contingente. Su programación es independiente de P y sólo afecta a cada ciclo en lo particular. Sus valores también varían de 1.0 a 0.0.
Inicialmente la probabilidad de ocurrencia de un estímulo dentro del sistema se podía definir en términos de la duración total del ciclo T y T, al darle un valor a T menor que el tiempo entre respuestas (TER) mínimo por parte del organismo. De esta manera, la probabilidad de reforzamiento para cualquiera de las respuestas se determinaba por el valor de T. A partir de este procedimiento, el parámetro P adquiría valores por debajo de la unidad (Brandauer, 1958; Sidley & Schoenfeld, 1964). Posteriormente, el parámetro P se definió de manera independiente de los parámetros de T y T a través de la integración en los programas temporales del sistema T (Snapper, 1962; Schoenfeld & Cole, 1972) o a través de dispositivos generadores de pulsos aleatorios que controlaban la elección de la disponibilidad del estímulo en cada ciclo (Schoenfeld, Cumming, Snapper & Haas, 1960; Farmer, 1963; Millenson, 1963).
Pese a la relativa independencia de los procedimientos para manipular estos parámetros, Martin (1971) sugirió que podrían estudiarse como parámetros funcionalmente equivalentes, en la medida en que ambos regulan la ocurrencia de estímulos como una razón del tiempo de disponibilidad para la producción del estímulo en relación con el tiempo total de exposición al programa. Sin embargo, los resultados de estudios iniciales con el sistema T sugieren que estos parámetros no son necesariamente equivalentes.
A diferencia de los resultados obtenidos al disminuir el valor de P, en que no se han observado efectos sistemáticos en la frecuencia de respuesta, salvo que la frecuencia más baja de respuesta se encontró con P = 1.0 (Brandauer, 1958; Farmer & Schoenfeld, 1967), los efectos de manipular T son relativamente sistemáticos. En general, los resultados mostraron que la frecuencia de respuesta aumentó conforme disminuía el valor de T y que el aumento en la tasa correlacionó con la disminución en la frecuencia de reforzamiento (Hearst, 1958; Clark, 1959).
En estudios más recientes se evaluó el efecto de reducir el valor de P bajo condiciones de entrega contingente (Martin, 1971; Carpio, González & Ribes, 1986; Carpio, 1989) y bajo condiciones de entrega no contingente (Bruner, 1981). En general, los resultados mostraron que la frecuencia de respuesta incrementó conforme disminuyó el valor de P. Este efecto se observó hasta el valor de P = 0.3, en que la frecuencia de respuesta mostró una función descendente. Estos estudios sugieren que los cambios en la frecuencia de respuesta son un efecto directo de P.
Resultados similares se encontraron en estudios posteriores en que se evaluaron los efectos de la disminución del valor de P en el desarrollo de funciones discriminativas de estímulos. En una serie de estudios (Ribes & Torres, 1996; Ribes & Torres, 1997; Ribes, Torres & Piña, 1999; Ribes, Torres & Mayoral, 2000a, 2000b, 2002a; Torres, Ribes & Mayoral, 2003) se examinaron los efectos de correlacionar estímulos neutros con distintos valores de probabilidad de entrega de agua en tD y en t. Los resultados mostraron un incremento en las tasas locales de respuesta conforme disminuía el valor de P en tD. Otro hallazgo sistemático fue que las frecuencias de respuesta en t fueron mayores que en tD, independientemente de la probabilidad asignada a cada subciclo, excepto cuando se evaluaron los valores de probabilidad local equivalentes. Por último, en ninguno de estos estudios se generaron patrones de respuesta que indicaran un control por parte del estímulo que fuera acorde con lo esperado al utilizar los programas de reforzamiento tradicionales (Ferster & Skinner, 1957; Rilling, 1983).
En un estudio posterior, Ribes, Torres y Mayoral (2002b) manipularon el valor de T en un ciclo de 30 s, manteniendo constante el valor de P bajo distintas condiciones de señalización de los subciclos tD y t. Los resultados de estos estudios no mostraron resultados distintos a los obtenidos en los estudios previos. Sin embargo, se pudo observar que bajo condiciones de señalización del subciclo tD, hubo un aumento de la frecuencia de respuesta como función de la disminución de la disponibilidad temporal en la entrega de agua, hasta que en T = 0.01 la frecuencia de respuesta fue cercana a cero. Este resultado no se obtuvo en las otras condiciones experimentales (con t señalado y con ambos subciclos señalados), ya que no se observaron cambios sistemáticos en la frecuencia de respuestas con las variaciones en T. Además, el porcentaje de entregas de agua perdidas no covarió directamente con la frecuencia de respuestas, en tanto que se perdieron porcentajes crecientes de entregas de agua bajo frecuencias de respuesta equivalente o mayores. Estos resultados sugieren una interacción entre la ubicación temporal de la respuesta, la señalización de diferencial dentro del ciclo y la disponibilidad temporal para la ocurrencia de la entrega de agua. Los parámetros de P y T parecieran no ser equivalentes funcionalmente. Sin embargo, en la medida en que bajo condiciones de señalización del subciclo tD la manipulación de T parece generar patrones similares de ejecución a los obtenidos al manipular P y que estos resultados se obtuvieron utilizando programas cuya duración de T era menor (30 s) a la tradicionalmente estudiada (60 s), es importante explorar paramétricamente las condiciones de señalización y su relación con T y T.
El presente experimento tiene por objetivo evaluar los efectos de variar la disponibilidad en la entrega de agua a través de la manipulación de los valores de T, bajo distintas condiciones de señalización de los subciclos en un programa temporal, manteniendo constante el valor de P.
Método
Sujetos
Se utilizaron cuatro ratas albinas machos (cepa Wistar) de 5 meses de edad, experimentalmente ingenuas, cuyo peso ad libitum osciló entre los 290 y 430 gramos. Las ratas permanecieron alojadas en jaulas plásticas individuales, con un acceso diario de 30` a agua purificada después de cada sesión experimental. Las ratas fueron privadas de agua 23 horas diarias antes del inicio del experimento. Las sesiones experimentales tuvieron lugar diariamente de 10 a.m. a 11 a.m., seis días a la semana durante ocho meses.
Aparatos
Se emplearon dos cámaras de condicionamiento operante de construcción propia (30 cm de largo por 25.5 cm de ancho y 32 cm de altura), colocadas dentro de cubículos sonoamortiguadores con extractores de aire de 115 w. Las cámaras tenían un dispensador de agua (0.01 cc) en la parte central inferior de la pared operativa, así como dos palancas equidistantes de cada lado del dispensador del agua. La palanca derecha era operativa y se ubicaba a 2.6 cm de la pared derecha y a 2.5 cm arriba del piso enrejado. La palanca izquierda no era operativa y se ubicaba a 2.8 cm de la pared izquierda y a 2.5 arriba del piso enrejado. Ambas palancas tenían 3.5 cm de ancho y sobresalían 2 cm. Las palancas requerían de 0.13 N de fuerza para operar. Se utilizaron cuatro luces de 28 v. Un foco proporcionó la iluminación general a la cámara, ubicado en la parte superior de la pared operativa arriba del dispensador. Un segundo foco se ubicó dentro del dispensador de agua. Se colocó una tecla translucida 6.5 cm arriba de la palanca operativa y detrás de ella se montaron dos focos de 5 w: uno proyectaba luz verde y el otro, luz roja. La luz general de la caja estaba encendida todo el tiempo excepto durante los tres segundos del periodo de disponibilidad de agua, en el que se encendía la luz del dispensador.
Procedimiento
Cada rata fue entrenada a presionar la palanca operativa por moldeamiento directo, que concluyó cuando cada animal obtuvo 100 entregas de agua consecutivas en la sesión. Se empleó un programa temporal de estímulos (Schoenfeld & Cole, 1972), consistente en un ciclo T continuo, repetitivo, de 60 s, dividido en dos subciclos, tD y t, cada uno con una duración dependiente del valor de T.
La probabilidad (p) de entrega del agua siempre fue de 1.0 para cada subciclo tD y de 0.0 para cada subciclo t. T tuvo un valor distinto en cada fase experimental. Los valores sucesivos de T fueron de 1.0, 0.5, 0.3, 0.1 y 0.01, con una fase final de redeterminación (ver Tabla 1). Los valores de T se programaron variando la duración del subciclo tD en 60, 30, 20, 6 y 0.6 s respectivamente, con un valor de 60 s en la fase de redeterminación. Cada fase experimental constó de 25 sesiones, excepto la fase final de redeterminación con sólo 10 sesiones. En cada sesión, el ciclo T iniciaba siempre con el subciclo tD y terminaba con el subciclo t, repitiéndose la secuencia hasta concluir 60 ciclos. Durante las fases 1 y 6, el subciclo tD se señalizó con una luz verde en cada uno de los sesenta ciclos que constituían la sesión. En las fases 2 a 5, las sesiones 1 a 10 y 16 a 25, cada uno de los subciclos se correlacionó con una luz diferencial. El subciclo tD se correlacionó con una luz verde, mientras que el subciclo t se correlacionó con una luz roja en los sesenta ciclos de la sesión. Durante las sesiones 11 a 15 (condición interpolada), en los primeros treinta ciclos (1-30) el subciclo tD se correlacionó con una luz verde, mientras que en el subciclo t no se presentó ninguna luz. En los últimos 30 ciclos (31-60), durante el subciclo tD no se presentó con ninguna luz, mientras que el subciclo t se correlacionó con una luz roja. Esta condición se implementó en las fases 1 a 5. Después de cada una de las fases experimentales se incluyeron periodos de sondeo de 5 sesiones cada uno. En cada periodo de sondeo se programaron los mismos valores de P y T y se presentaron los subciclos tD y t correlacionados con las mismas luces que en la fase experimental precedente inmediata. En el periodo de sondeo, el agua se entregaba independientemente de la respuesta al término del subciclo correspondiente. Durante las fases experimentales, la entrega del agua, contingente a la primera respuesta en tD, apagaba la luz verde. En caso de no haber respuesta en el subciclo tD, la luz se apagaba cuando se cumplía la duración del ciclo correspondiente. En los periodos de sondeo, la luz se apagaba al entregarse el agua al cumplirse el valor del subciclo tD. En t la luz era coextensiva con el subciclo.
Resultados
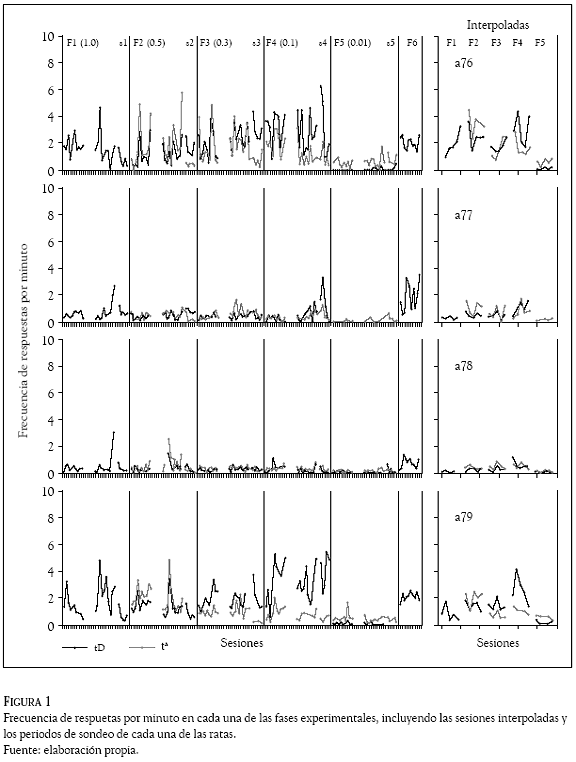
La Figura 1 muestra la frecuencia de respuestas por minuto en cada una de las fases experimentales, incluyendo las sesiones interpoladas y los periodos de sondeo de cada una de las ratas. En las gráficas se observan dos tipos de ejecuciones. Por un lado las ratas a76 y a79 mostraron una ejecución ascendente en el subciclo tD conforme disminuyó el valor de T durante las Fases 1 a 4 con un rango de 1 a 6 respuestas por minuto tanto en las condiciones de presentación de agua contingente como no contingente. Durante la Fase 5 (T = 0.01) la frecuencia de respuesta por minuto descendió hasta niveles cercanos a 0 en tD y de 1 en t. Por su parte, las ratas a77 y a78 no mostraron cambios en la ejecución conforme varió el valor de T. Durante la Fase 6 (redeterminación) todas las ratas mostraron un incremento en la frecuencia de respuesta con valores cercanos a 2 respuestas por minuto. Aunque las frecuencias locales de respuesta de las ratas fueron muy variadas a lo largo de las condiciones experimentales, se observó una frecuencia mayor en t que en tD cuando T tuvo un valor de 0.5. La ejecución durante las sesiones interpoladas fue similar a la obtenida en las sesiones experimentales en cada una de las fases en todas las ratas. En las sesiones con entrega de agua no contingente (sondeo), la frecuencia de respuesta en tD siempre estuvo por encima de la mostrada en t, con excepción de la Fase 5 donde la frecuencia en tD fue menor o igual a t.
La Figura 2 muestra la frecuencia por minuto obtenida cuando sólo se señaló tD y cuando sólo se señaló t durante las sesiones interpoladas. Los datos mostraron que en los ciclos donde se señalaba exclusivamente el subciclo tD, tres de las cuatro ratas (a76, a77 y a79) mostraron un incremento en la frecuencia de respuesta conforme disminuyó T hasta la Fase 5, cuando T tuvo un valor de 0.01y las frecuencias de respuesta cayeron a valores cercanos a cero.
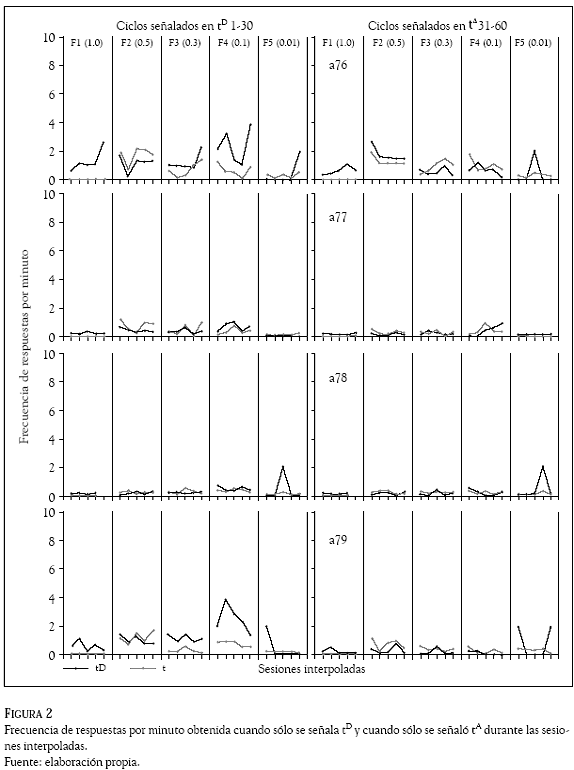
En esta misma condición, las frecuencias de respuesta en el subciclo t fueron menores que en el subciclo tD, con excepción de la fase 5. Por su parte, en los ciclos donde se señaló exclusivamente t, el efecto fue distinto en la medida en que las ratas mostraron una ejecución relativamente estable a lo largo de todas las condiciones experimentales, con excepción de la rata a76 que mostró un pequeño incremento de la Fase 1 a la 2 (T de 1.0 a 0.5) y a partir de esa fase una disminución gradual conforme disminuyó el valor de T. Sólo en las ratas a76 y a79 se observaron frecuencias ligeramente mayores en t que en tD.
La Figura 3 muestra el porcentaje promedio de entregas de agua perdidas en cada una de las fases experimentales y de redeterminación para cada una de las ratas. Se observó una pérdida por encima del 50% de entregas de agua en las ratas a76 y a79 mientras que en las ratas a77 y a78 fue por encima del 80% a lo largo del experimento.
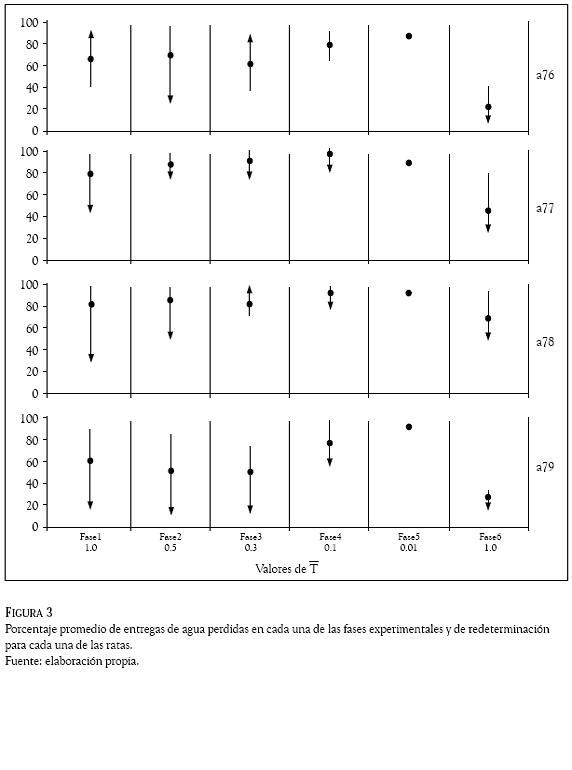
En dos de las ratas (a76 y a79), se observó una función descendente en los valores iniciales de T (1.0, 0.5 y 0.3) y a partir de la Fase 4 se observó un incremento en el porcentaje de pérdidas de agua (T = 0.1 y 0.01). En las otras dos ratas (a77 y a78) se observó una función ascendente conforme disminuyó el valor de T. En la fase de redeterminación se observó una disminución en el porcentaje de aguas perdidas (entre el 20 y el 60%) en todas las ratas.
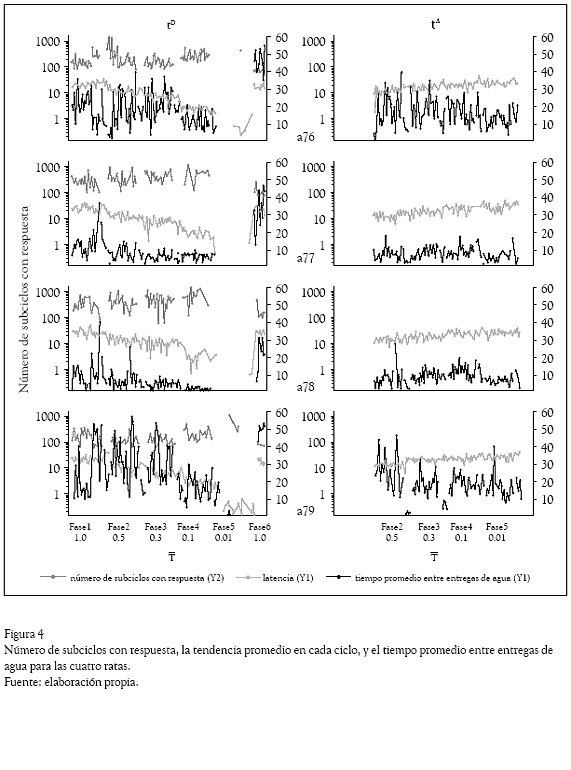
La Figura 4 muestra el número de subciclos con respuesta, la latencia promedio en cada ciclo y el tiempo promedio entre entregas de agua para las cuatro ratas, tanto en tD como en t. En general se observaron efectos diferenciales entre los subciclos. Durante el subciclo tD se observó, en tres de las cuatro ratas, un efecto decreciente en la latencia y el número de subciclos con respuesta conforme disminuyó el valor de T, hasta llegar a valores cercanos a cero durante la Fase 6 con T igual a 0.01. Por su parte, en el subciclo t se observó una tendencia ligeramente ascendente en el valor de la latencia a lo largo de las fases experimentales, mientras que el número de subciclos con respuesta se mantuvo relativamente constante. El tiempo promedio entre entregas de agua en general fue supe rior a 100 segundos, llegando a valores cercanos a 1000 sobre todo en las ratas a77 y a78.
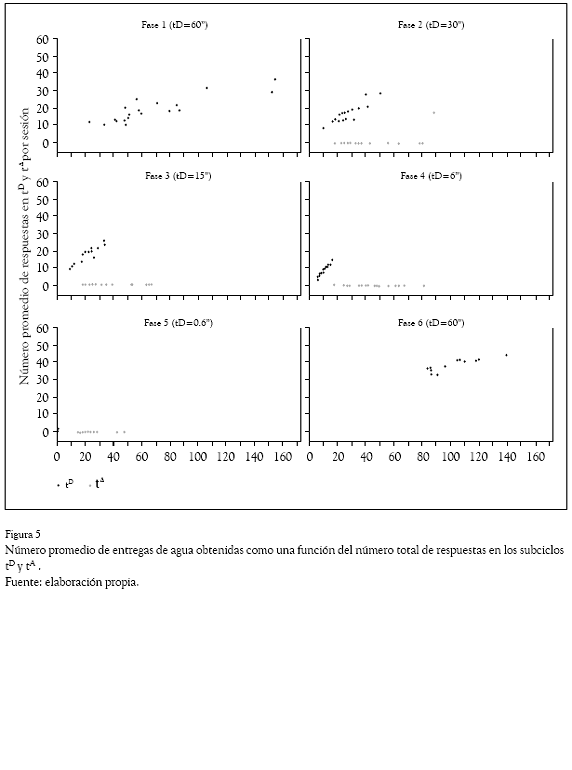
La Figura 5 muestra el número promedio de entregas de agua obtenidas como una función del número total de respuestas en los subciclos tD y t. En general se observó que el número total de entregas de agua por sesión aumentó como función del número total de respuestas y que el número total de respuestas decreció conforme disminuyó el valor de T, aunque esto no afectó directamente el número de entregas obtenidas, salvo en la Fase 5 (T = a 0.01) donde el número de entregas y respuestas en tD fueron cercanas a cero.
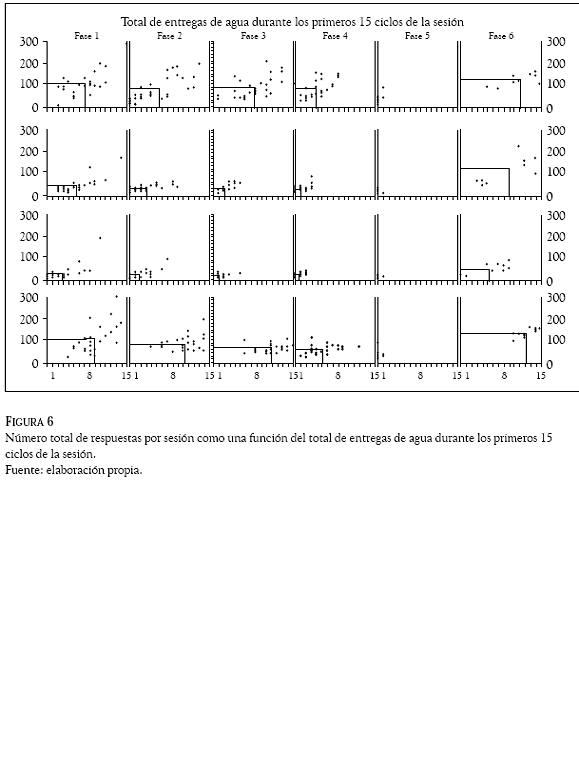
Por último, la figura 6 muestra el número total de respuestas por sesión como una función del total de entregas de agua durante los primeros 15 ciclos de la sesión. Se observó en todas las ratas que el número total de respuesta fue una función del número total de entregas de agua durante los primeros quince ciclos, aunque se observó un decremento gradual del total de respuestas por sesión conforme disminuyó el valor de T. Durante la fase de redeterminación (Fase 6) todas las ratas mostraron un incremento en el número de entregas de agua por sesión, aunque no necesariamente se observó el mismo incremento en el número total de respuestas en tres de las cuatro ratas, que no llegó a los niveles de respuesta obtenidos en las tres primeras fases en las ratas a 76, a78 y a79.
Discusión general
Los resultados de este estudio no son consistentes con resultados obtenidos previamente al manipular los valores de T en programas temporales señalados, en que no se observaron cambios sistemáticos en la frecuencia de respuesta conforme se varió el valor de T (Ribes, Carpio, Pallares & Torres, 1986; Ribes, Torres & Mayoral, 2002b), los resultados en el presente trabajo no son conclusivos. En términos generales, dos de las ratas (a76 y a79) mostraron un incremento en la frecuencia de respuesta en el subciclo tD conforme decrecía el valor de T hasta llegar al valor de 0.01, en que se observaron frecuencias de respuesta cercanas a cero, mientras que las otras dos ratas (a77 y a78) no mostraron ningún cambio sistemático en la frecuencia de respuesta asociada a la cambios en el valor de T.
En el estudio de Ribes, Torres y Mayoral (2002b), la disminución de la disponibilidad de reforzamiento a través de manipular el valor de T no pareció afectar el patrón de ejecución cuando se señalizó el subciclo t de manera exclusiva (Grupo 2) o conjuntamente con tD (Grupo 3). Sin embargo, cuando sólo se señalizó el subciclo tD, se pudo observar una disminución de la frecuencia conforme disminuyó T. Estos resultados sugieren que los parámetros T y P no son funcionalmente equivalentes, en la medida en que la frecuencia de respuesta no se ve afectada directamente por las variaciones en T. Sin embargo, al considerar el porcentaje de entregas de agua perdidas, los resultados mostraron un aumento conforme se manipuló el valor de T sólo en los grupos 2 y 3. En el grupo 1, el porcentaje de entregas de agua perdidas se mantuvo estable. En su conjunto, estos resultados lo que demuestran es que, al parecer, hay una relación entre la señalización, la distribución en la entrega de agua y su disponibilidad temporal. De acuerdo con los autores, bajo la contingencia establecida en los programas T, la distribución temporal de la respuesta (sólo se requiere una respuesta por ciclo) es más importante que la emisión de frecuencias más o menos elevadas.
En el presente estudio se observó un porcentaje elevado de pérdida de entregas de agua (en general del 60% o mayor). En dos de las ratas (a 76 y a79), dicho porcentaje covarió de manera directa con la frecuencia de respuesta. También se observó en estas dos ratas que la frecuencia de respuesta aumentó conforme disminuyó T. Por su parte, las otras dos ratas (a77 y a78) mostraron un porcentaje de entregas de agua perdidas mayor (80% o superior), el cual aumentó conforme disminuyó el valor de T. Sin embargo, en la fase 6 de redeterminación de los valores iníciales, se observó una disminución considerable del porcentaje de entregas de agua perdida en las cuatro ratas (alrededor del 40%), aun cuando la frecuencia de respuesta fue similar a la obtenida en la fase 1. Este dato apoya el hecho de que la ejecución en los programas temporales está determinada más por la distribución temporal de la respuesta que por su frecuencia (Ribes, Torres & Mayoral, 2002). De hecho, la diferencia entre P y T tiene que ver con la distribución temporal versus la disponibilidad temporal en la presentación de los estímulos. El parámetro P hace referencia a la distribución temporal de la entrega de agua durante la sesión y se sintetiza en términos de la razón T/P, mientras que el parámetro de T se relaciona directamente con el valor absoluto del tiempo disponible para responder y que se presente el agua, independientemente de la frecuencia programada.
Otro efecto que se ha observado sistemáticamente en programas temporales con estímulos diferenciales, tiene que ver con las diferencias locales en la frecuencia de respuesta. En estudios previos, donde se ha manipulado el valor de P (Ribes & Torres, 1996; Ribes, Torres & Mayoral, 2000a, 2000b, 2002a; Torres, Ribes & Mayoral, 2003), los datos muestran una frecuencia de respuesta mayor en el subciclo con una menor probabilidad de reforzamiento. Además, se ha observado una alta variabilidad en la frecuencia de respuesta entre sesiones, en ocasiones asociadas con un alto porcentaje de entregas de agua perdidas. En la medida en que los programas temporales (Schoenfeld et al., 1956) prescriben que una disponibilidad temporal limitada de reforzamiento, en contraste con los programas de reforzamiento operante (Ferster & Skinner, 1957), algunas entregas de agua se pierdan si la respuesta no ocurre en los periodos (tD) de disponibilidad del reforzador. Cuando este periodo correlaciona con un estímulo, la ocurrencia de la primera respuesta, que produce la entrega del reforzador, también termina con la presentación del estímulo correlacionado. De esta manera, sólo la respuesta que produce el reforzador está correlacionada con el estímulo, por lo que las respuestas subsecuentes dentro del periodo tD ocurren en ausencia del estímulo, mientras que el responder en t (normalmente asociado a extinción o con una probabilidad baja de reforzamiento) está correlacionado con un estímulo diferente. Es razonable suponer que la variabilidad observada entre sesiones y entre sujetos en la frecuencia de respuesta, se debe a la variabilidad temporal en la entrega del agua, debida a la interacción de la aleatoriedad de la probabilidad de reforzamiento, las condiciones de señalización, la ausencia de respuestas en los ciclos con disponibilidad y el valor de cada uno de los subciclos. De hecho, los programas temporales a diferencia de los programas de reforzamiento tradicional, requieren de tasas o frecuencias diferenciales de respuesta para que ocurra el reforzamiento en una posición temporal determinada (Lee, Sturmey & Fields, 2007).
Sin embargo, en el presente estudio sólo en la condición de T = 0.5 se observó una mayor frecuencia de respuesta durante el subciclo t. En el resto de las condiciones experimentales se observaron frecuencias locales similares o mayores en tD que en t, sobre todo en las ratas (a 76 y a 79) sugiriendo un efecto diferencial del estímulo correlacionado con tD. Los resultados en el presente estudio sugieren la pertinencia de evaluar experimentalmente las interacciones entre los parámetros P y T bajo distintas duraciones del ciclo T.
Bibliografía
Brandauer, C. M. (1958). The effects of uniform probabi lities of reinforcement on the response rate of the pigeon. Tesis Doctoral, Columbia University. [ Links ]
Bruner, C. A. (1981). The effect of cycle length, interstimulus interval and probability of reinforcement in autoshaping. Revista Mexicana de Análisis de la Conducta, 7, 149-157. [ Links ]
Clark, R. (1959). Some time-correlated reinforcement schedules and their effects on behaviour. Journal of the Experimental Analysis of Behavior, 2, 1-22. [ Links ]
Carpio, C. A. (1989). Efectos de la variación de la probabilidad y la disponibilidad temporal relativa de reforzamiento en programas temporales, señalados y no señalados de distinta duración. Tesis de Maestría, UNAM. [ Links ]
Carpio, C. A., González, R. & Ribes, E. (1986). Probabilidad de reforzamiento y su señalización en un programa definido temporalmente. Revista Mexicana de Análisis de la Conducta, 12, 89-104. [ Links ]
Farmer, J. (1963). Properties of behaviour under random interval reinforcement schedules. Psychonomic Science, 7, 173-174. [ Links ]
Farmer, J. & Schoenfeld, W. N. (1967). Response rate under random interval schedules of reinforcement. Journal of the Experimental Analysis of Behavior, 6, 437-443. [ Links ]
Ferster, C. B. & Skinner, B. F. (1957). Schedules of Reinforcement. New York: Appleton Century Crofts. [ Links ]
Hearst, E. (1958). The behavioural effect of some temporally-defined schedules of reinforcement. Journal of the Experimental Analysis of Behavior, 1, 45-55. [ Links ]
Lee, R., Sturmey, P. & Fields, L. (2007). Scheduleinduced and operant mechanisms that influence response variability: A review and implications for future investigations. The Psychological Record, 57, 429-455. [ Links ]
Martin, J. M. (1971). Temporally defined schedules of stimulus correlations. Tesis Doctoral no publicada, City University of New York, USA. [ Links ]
Millenson, J. R. (1963). Random interval schedules of reinforcement. Journal of the Experimental Analysis of Behavior, 6, 437-443. [ Links ]
Ribes, E., Carpio, C. A., Pallares, A. & Torres, J. (1986). Efectos de la reducción en la disponibilidad de reforzamiento en un programa temporal señalado. Revista Mexicana de Análisis de la Conducta, 22, 127-136. [ Links ]
Ribes, E. & Torres, C. (1996). Efectos de la variación de la probabilidad de reforzamiento correlacionada con dos estímulos neutros en un programa definido temporalmente. Revista Mexicana de Análisis de la Conducta, 22, 41-78. [ Links ]
Ribes, E. & Torres, C. (1997). Stimulus functions: Lack of discrimination or escess of generalization Revista Mexicana de Análisis de la Conducta, 23, 249-274. [ Links ]
Ribes, E., Torres, C. & Piña, J. (1999). Comparación de los efectos de la presencia y ausencia de estímulos diferenciales en dos programas temporales con probabilidades variantes igualadas. Acta Comportamentalia, 7, 5-29. [ Links ]
Ribes, E., Torres, C. & Mayoral, A. (2000a). Señalización no diferencial de distintas probabilidades de entrega de agua en dos subciclos de un programa definido temporalmente. Acta Comportamentalia, 8, 5-21. [ Links ]
Ribes, E., Torres, C. & Mayoral, A. (2000b). Efectos de la ausencia y presencia de estímulos correlacionados con distintas probabilidades de reforzamiento y con extinción en programas definidos temporalmente. Revista Mexicana de Análisis de la Conducta, 26, 327-354. [ Links ]
Ribes, E., Torres, C. & Mayoral, A. (2002a). Extended exposure to a discriminated limited-hold temporal schedule does not produce stimulus control. Behavioural Processes, 59, 131-146. [ Links ]
Ribes, E., Torres, C. & Mayoral, A. (2002b). Señalización de la entrega y no entrega de agua en programas temporales con variaciones en la disponibilidad de reforzamiento. Acta Comportamentalia, 10, 129-149. [ Links ]
Rilling, M. (1983). Control de estímulos y procesos inhibitorios. En W. K. Honig & J. E. R. Staddon (Eds.), Manual de Conducta Operante (pp. 432- 480). México: Trillas. [ Links ]
Schoenfeld, W. N. & Cole, B. K. (1972). Stimulus schedules:The t-tau system. New York: Harper & Row Publishers. [ Links ]
Schoenfeld, W. N., Cumming, W. W. & Hearst, E. (1956). On the classification of reinforcement schedules. Proceedings of the National Academy of Science, 42, 536-570. [ Links ]
Schoenfeld, W. N., Cumming, W. W., Snapper, A. G. & Haas, P. (1960). Some electronic control units for operant behaviour studies: II. A random ratio generator. Journal of the Experimental Analysis of Behavior, 3, 107-108. [ Links ]
Sidley, N. A. & Schoenfeld, W. N. (1964). Behavior stability and response rate as functions of reinforcement probability on “random ratio” schedules. Journal of the Experimental Analysis of Behavior, 6, 265-268. [ Links ]
Snapper, A. G. (1962). Properties of behaviour under response independent temporally defined reinforcement schedules. Tesis Doctoral no publicada, Columbia University. [ Links ]
Torres, C., Ribes, E. & Mayoral, A. (2003). Efectos de probabilidades complementarias y contrastadas de reforzamiento con y sin señalización en programas temporales. Acta Comportamentalia, 11, 130-149. [ Links ]
Recibido: marzo 9 de 2008
Revisado: mayo 11 de 2008
Aceptado: mayo 12 de 2008
*Toda correspondencia debe dirigirse al primer autor a: Centro de Estudios e Investigaciones en Comportamiento, Francisco de Quevedo #180 (Arcos Vallarta), Guadalajara, Jal. 44100. Correo electrónico: jtorres@cencar.udg.mx

