








Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores
Compartir

 Permalink
PermalinkNeuropsicologia Latinoamericana
versión On-line ISSN 2075-9479
Neuropsicologia Latinoamericana vol.7 no.2 Calle 2015
https://doi.org/10.5579/rnl.2015.0242
DOI:10.5579/rnl.2015.0242
Mediación del fascículo uncinado en el volumen de la amígdala y la corteza orbitofrontal
Médiation du fascicule unciforme et le volume entre l'amygdale et le cortex orbitofrontal
Mediação do fascículo uncinado no volumen da amígdala e do córtex orbitofrontal
Mediation of the uncinate fascicle between the amygdala and orbitofrontal cortex volume
Julián MarinoI,II; Fernando G. LunaI; Rodrigo JaldoI; Gustavo Foa TorresIII,IV; Damián CenturiónIV; Alberto Acosta MesasII
I Laboratorio de Procesamiento de Neuroimágenes, Facultad de Psicología, Universidad Nacional de Córdoba, Argentina
II Centro de Investigación Mente, Cerebro y Comportamiento, Universidad de Granada, España
III Facultad de Ciencias Médicas, Universidad Nacional de Córdoba, Argentina
IV Centro Privado de Tomografía Computada Córdoba S.A. – Fundación Oulton, Córdoba, Argentina
RESUMEN
La amígdala y la corteza orbitofrontal son estructuras cerebrales en estrecha relación en el procesamiento de emociones. El fascículo uncinado es un tracto de sustancia blanca que une ambas estructuras y que comunica el polo temporal. De esta manera, forman una red importante de regulación de emociones y procesos semánticos, ya que en el polo temporal se computan los procesos de abstracción conceptual. Se utilizaron secuencias de resonancia magnética de pesos de difusión y T1 en 32 jóvenes de sexo femenino (18-30 años de edad) para profundizar la relación entre las estructuras amigdalino- frontales y el tracto uncinado. Durante el post-análisis se realizaron técnicas de imágenes por tensores de difusión y morfometría basada en voxels. Se calculó el volumen en mililitros de las estructuras cerebrales y la conectividad mediante anisotropía fraccional del fascículo uncinado. El análisis de datos se realizó con un modelo de mediación (test de Sobel) con procedimiento Bootsrapping. Nos propusimos indagar si la conectividad del fascículo uncinado mediaba la influencia del tamaño de la amígdala sobre el tamaño de la corteza orbitofrontal. Los resultados principales fueron que la amígdala derecha predice sin mediaciones de conectividad el volumen orbitofrontal derecho, pero que en el hemisferio izquierdo el rol de la mediación por parte del fascículo uncinado es más complejo y cruzado.
Palabras clave: Fascículo uncinado, Amígdala, Corteza orbitofrontal, Resonancia magnética, Test de Sobel
RÉSUMÉ
L'amygdale et le cortex orbitofrontal sont des structures cérébrales qui sont étroitement liés au traitement des émotions. Le faisceau unciforme est une question paquet blanc reliant les deux structures et le pôle temporal. Ces deux structures de la matière grise et ce bundle est conforme un important réseau de réglementation sémantique et émotionnelle. IRM de diffusion pondérée et des séquences T1 ont été utilisés dans 32 des jeunes femmes (18-30 ans) pour approfondir la relation dans le réseau susmentionné. Au cours de la post-analyse ont été effectués techniques d'imagerie basée sur l'imagerie du tenseur de diffusion conjoint avec voxels morphométrie. On a calculé le volume en millilitres de deux structures cérébrales et de la connectivité du faisceau uncinate par anisotropie fractionnée. L'analyse des données a été effectuée en utilisant un modèle de médiation (test de Sobel) avec Bootsrapping technique. L'hypothèse était si la connectivité unciforme était un médiateur de l'influence de la taille de l'amygdale sur la taille du cortex orbitofrontal. Les principales conclusions sont que l'amygdale droite prédit directement (sans intermédiaire) le volume orbitofrontal droit, mais dans l'hémisphère gauche du rôle de la médiation par le faisceau était unciforme plus complexe et hémisphérique croix. De nouvelles données sont nécessaires pour trouver plus de preuves sur ces relations de chemin.
Mots clés: Fascicule unciforme, Amygdale, Cortex orbitofrontal, Imagerie par résonance magnétique, Sobel Test.
RESUMO
A amígdala e o córtex orbitofrontal são estruturas cerebrais que tem uma estreita relação com o processamento das emoções. O fascículo uncinado é um trato de substância branca que comunica o pólo temporal e isto demonstra uma rede importante de regulação das emoções e de processos semânticos. Utilizou-se sequências de ressonância magnética de pesos de difusão em T1 em 32 indivíduos do sexo feminino (18 a 30 anos de idade) para investigar a relação entre as estruturas amígdala-frontal e o trato uncinado. No pós-processamento utilizou-se técnicas de imagens por tensor de difusão e morfometria baseada em voxels. Calculou-se o volume em milímetros das estruturas cerebrais e a conectividade mediante anisotropía fracionada do fascículo uncinado. As análises de dados se realizaram a partir do modelo de mediação (Teste de Sobel) com procedimento Bootstrapping. Os resultados principais indicaram que a amígdala direita pred2iz sem mediações de conectividade o volume orbitofrontal direito, mas o hemisfério esquerdo tem o papel de mediação por parte do fascículo uncinado é mais complexo e cruzado.
Palavras-chave: Fascículo uncinado, Amígdala, Córtex orbitofrontal, Resonância magnética, Test de Sobel.
ABSTRACT
The amygdala and orbitofrontal cortex are brain structures that are closely related in emotional processing. The uncinate fasciculus is a white matter bundle connecting the two structures and the temporal pole. These two gray matter structures and this bundle conforms an important semantic and emotional regulatory network. MRI diffusion weighted and T1 sequences were used in 32 young women (18-30 years old) to deepen the relationship in the aforementioned network. During the post-analysis were performed imaging techniques based on diffusion tensor imaging conjoined with voxels based morphometry. It was calculated the volume in milliliters of the two brain structures and the connectivity of the uncinate fasciculus by fractional anisotropy. Data analysis was performed using a model of mediation (Sobel test) with Bootsrapping technique. The hypothesis was if the uncinate connectivity was a mediator of the influence of the size of the amygdala on the size of the orbitofrontal cortex. The main findings were that the right amygdala directly predicts (unmediated) the right orbitofrontal volume, but in the left hemisphere the role of mediation by the uncinate fasciculus was more complex and cross hemispheric. New data are necessary to find more evidence over these path relationships.
Key words: Uncinate fascicle, Amygdala, Orbitofrontal cortex, Magnetic resonance imaging, Sobel Test.
1. INTRODUCCIÓN
Las medidas de imágenes por tensores de difusión resultaron una de las revelaciones de los últimos tiempos en lo que respecta a la medición precisa del grado de conexión entre estructuras cerebrales a través de los fascículos de sustancia blanca (Bosch et al., 2012). Los gradientes de conectividad cerebral se miden esencialmente mediante la anisotropía fraccional cerebral. Esta es una medida de la difusión de las moléculas de agua dentro de las fibras nerviosas. A mayor anisotropía fraccional, mayor es la conectividad entre dos o más estructuras cerebrales. La anisotropía fraccional está determinada por la integridad de la fibra (más o menos compacta), el número de fibras que la integran, el nivel de mielinización y 'la elegancia' de su forma (Leemans, Jeurissen, Sijbers, & Jones, 2009). Por elegancia se entiende la ausencia de fragmentos de fibras que obstaculizan el paso de las moléculas de agua en la dirección principal de tráfico (Lochner et al., 2012). La dirección principal de una fibra corresponde a un espacio relativo tridimensional trazado sobre la fibra-objetivo.
De este modo quedan determinados tres vectores de dirección con sus respectivos autovectores (λ1, λ2 y λ3). El movimiento isotrópico tiene un grado de anisotropía igual a cero, no se revelan diferencias en la señal de movimiento de la molécula de agua captado en una dirección. Esto ocurre por ejemplo en el líquido cefalorraquídeo, que no le presenta obstáculos. En los tractos de sustancia blanca se encuentran niveles de anisotropía en favor de uno de los tres vectores. Las fórmulas para calcular la anisotropía fraccional son variadas (Smith et al., 2006), pero similares en su concepto: el privilegio del movimiento browniano de las moléculas de agua en dirección del autovector (λa) durante un tiempo específico en comparación con las dos direcciones de vectores (λb y λc) ortogonales.
El fascículo uncinado es un tracto de sustancia blanca cerebral que conecta la amígdala con la zona prefrontal. Es una pequeña fibra en forma de hoz. Se la considera uno de los últimos tractos evolutivos de la especie humana (Catani & Thiebaut de Schotten, 2008). La corteza orbitofrontal está relacionada de forma estrecha con la amígdala a través del fascículo uncinado. También el polo temporal queda unificado con las estructuras mencionadas mediante ese tracto (Highley, Walker, Esiri, Crow, & Harrison, 2002). El fascículo uncinado se divide en dos porciones: una dorsal, que llega a la corteza prefrontal y otra ventral, que desciende al 'pozo filogenético' amigdalino (Damasio, Tranel, Grabowski, Adolphs, & Damasio, 2004; Pessoa & Adolphs, 2010). Un gradiente bajo en anisotropía fraccional en el fascículo uncinado fue relacionado con tener pocos amigos, abundantes problemas interpersonales, escasa capacidad de regulación emocional, y altos niveles de agresividad (Phan et al., 2010). En un estudio de fuerte impacto, (Eluvathingal et al., (2006) encontraron que en niños/as huérfanos, migrantes de la guerra balcánica, el fascículo uncinado no se había desarrollado. Presentaban un comportamiento a simbólico, desorganizado e impulsivo.
La amígdala es una pequeña estructura subcortical relacionada con las emociones de miedo. La sobreactividad de la amígdala está asociada a los trastornos de ansiedad, depresión y fobia (Phan et al., 2010). A mayor tamaño de la amígdala, más probabilidades existen de contraer las mencionadas psicopatologías (Tottenham et al., 2010; van Marle et al., 2011). La corteza orbitofrontal fue hallada responsable del appraisal o etiquetamiento afectivo (Ochsner & Gross, 2005), que se define por la posibilidad de categorizar una emoción. La señal somática emocional proviene de estructuras como la amígdala, el tálamo o el hipotálamo. Entre la amígdala y la corteza orbitofrontal está el foco de la regulación emocional (Kalisch, 2009). El lenguaje es el contexto de las emociones (Barrett, Lindquist, & Gendron, 2007) y la primera parte de la regulación emocional consiste en brindar una etiqueta lingüística para el impacto emocional. Diversos estudios mostraron que una reducción del tamaño de la corteza orbitofrontal está relacionada con la presencia de trastornos de ansiedad y depresión (Feldmann et al., 2008; Vasic, Walter, Höse, & Wolf, 2008). De este modo, resulta de interés conocer la interacción entre el volumen de la amígdala, la corteza orbitofrontal y la posible mediación de la conectividad del fascículo uncinado. La historia de esta relación está presente en la explicación de la toma de decisiones, de la noción de marcador somático y de modelos clásicos de trastornos psicopatológicos como la ansiedad y la depresión (Bechara, Damasio, Tranel, & Damasio, 2005; Eysenck, Derakshan, Santos, & Calvo, 2007). La corteza orbitofrontal actúa de filtro de la información amigdalina. Su función es evitar que las emociones negativas provoquen disrupciones en el espacio de trabajo cognitivo. La cognición fría, a la que responde este último concepto, está relacionada con la corteza dorsolateral prefrontal (Kerr & Zelazo, 2004).
En el presente estudio se utilizaron secuencias de resonancia magnética combinadas (imágenes sin pesos de difusión y por tensores de difusión) para evaluar la conectividad en el fascículo uncinado, el volumen de la amígdala y el volumen de la corteza orbitofrontal. El objetivo fue conocer si el volumen de la corteza orbitofrontal estaba mediado por la anisotropía fraccional del fascículo uncinado en función del tamaño de la amígdala. En otras palabras, si el volumen de la amígdala podía explicar el tamaño de la corteza orbitofrontal y en qué medida esto se encuentra mediado por el nivel de conectividad entre estas estructuras cerebrales.
La hipótesis de trabajo es que el volumen de la amígdala será un predictor del tamaño de la corteza orbitofrontal mediatizado por el gradiente de conectividad del fascículo uncinado. Esto significa que habrá una predicción significativa del volumen en mililitros de la corteza orbitofrontal a partir del volumen de la amígdala en función de cuán fluido sea el tráfico de información entre las dos estructuras. Cuando el monto de información (medido por anisotropía fraccional) sea bajo, la amígdala de mayor tamaño predeciría una corteza orbitofrontal de menor tamaño. De encontrar evidencias en favor de esta hipótesis se apoyaría con estadísticos de mediación (Preacher & Hayes, 2004) el modelo de 'falla del filtro' en trastornos de depresión, fobias y ansiedad generalizada. De forma simplificada, la falla del filtro implicaría que la corteza orbitofrontal tendría dificultades para inhibir la actividad de los circuitos reverberantes amigdalinos (Cardinal, Parkinson, Hall, & Everitt, 2002).
2. MÉTODO
2.1. Participantes
Participaron voluntariamente 32 mujeres estudiantes y egresadas de la Universidad Nacional de Córdoba, Argentina, de entre 18 y 36 años (M=24,34; DS=5,24). Se realizó una entrevista clínica y una evaluación cognitiva previa para detectar antecedentes de patologías psiquiátricas o clínicas, siendo excluyente para la participación en el estudio el contar con diagnósticos psicopatológicos. El estudio fue aprobado por el comité de ética del Instituto de Tomografía Computada Córdoba – Fundación Oulton y cada participante dio su consentimiento informado por escrito.
2.2. Procedimiento
Cada participante realizó la adquisición de imágenes en una sola sesión y asistiendo de manera individual en el Centro de Tomografía Computada Córdoba - Fundación Oulton. Una vez firmado el consentimiento informado, se realizó la entrevista clínica y la evaluación cognitiva. Posteriormente se explicó a cada participante las condiciones para la adquisición de imágenes. Cuando los participantes ingresaron al resonador se les dieron cinco minutos de relajación y adaptación. Luego se iniciaron las secuencias de adquisición de imágenes estructurales (sin pesos de difusión y con pesos de difusión). Una vez finalizada esta etapa se administró una batería de pruebas neuropsicológicas de lenguaje, atención, regulación emocional y funciones ejecutivas, que serán reportadas en otros estudios. El procedimiento total tuvo una duración aproximada de 100 minutos por participante.
2.3. Adquisición de imágenes
Las imágenes fueron obtenidas mediante un scanner Philips Achieva (1.5 T) perteneciente al Centro de Tomografía Computada Córdoba - Fundación Oulton. Para las imágenes estructurales sin peso de difusión aparente se utilizó una matriz de reconstrucción de 232 X 232, con una secuencia T1 volumétrica con los siguientes parámetros: amplitud por 'slice' 1 mm, gap 0 mm; TR/TE 500/50, tamaño de voxels: 1 X 1 X 1. El campo de visión tuvo los siguientes parámetros: FOV RL (mm) 257 FOV AP (mm) 256 FOV FH (mm) 176.
Para las imágenes estructurales sin peso de difusión aparente se utilizó una matriz de reconstrucción de 232 X 232, con una secuencia T1 volumétrica con los siguientes parámetros: amplitud por 'slice' 1 mm, gap 0 mm; TR/TE 500/50, tamaño de voxels: 1 X 1 X 1. El campo de visión tuvo los siguientes parámetros: FOV RL (mm) 257 FOV AP (mm) 256 FOV FH (mm) 176. En las imágenes obtenidas por tensores de difusión se utilizó una matriz de reconstrucción de 112 X 132, con secuencias echo-planares DTI 'single-shot' con los siguientes parámetros: amplitud por 'slice' 1.5 mm, gap 0 mm; TR/TE 21386/76, tamaño de voxels: 1.5 X 1.5 X 1.5; cantidad de direcciones de difusión = 64; valor β en unidades s/mm2 = 1000. El campo de visión tuvo los siguientes parámetros: FOV RL (mm) 192 FOV AP (mm) 226 FOV FH (mm) 105.
2.4. Post procesamiento de imágenes
Imágenes sin pesos de difusión
El post-procesamiento fue realizado offline utilizando Matlab 7.10.0 y Statistical Parametrical Mapping 8 (SPM8). Las imágenes adquiridas de secuencia T1 fueron convertidas a formato 'Niifti', y se generó la reconstrucción de los 160 cortes adquiridos en un único volumen. Los espacios nativos cerebrales fueron reorientados, utilizando como espacio cerebral de referencia la imagen canónica avg152T1 creada en el Instituto Neurológico de Montreal (acrónimo MNI en inglés) (Van Hecke et al., 2011). Se utilizó como punto de referencia estructural la corteza cingulada anterior. Los espacios cerebrales nativos fueron reorientados en sus tres ejes espaciales (X, Y, Z) y rotados hasta mostrar una diferencia menor a 5 cm del punto de referencia del espacio cerebral canónico del MNI.
Posteriormente, se realizó el proceso de normalización y segmentación de tejidos. Se calcularon los mapas de los tres tejidos principales: sustancia gris, sustancia blanca, y líquido cefalorraquídeo. Se descartaron los voxels con tejidos que no eran de interés para el análisis: cráneo, tejidos blandos y aire. Para la normalización de alta dimensionalidad se utilizó el algoritmo DARTEL creado por John Ashburner para deformar sobre la plantilla el espacio nativo. DARTEL tiene grandes ventajas sobre otras formas de normalización, en especial su modularización no lineal (John Ashburner, 2007). Se cargaron 18 iteraciones para el procesamiento de cada imagen, conservadas para conocer los estiramientos y contracciones de la imagen nativa. Para las probabilidades gaussianas a priori por tejido se colocaron las probabilidades apriori de sustancia gris, sustancia blanca, líquido céfalorraquídeo, huesos, tejidos blandos y espacio extra cerebral-aire. Se emplearon Campos de Markov que le indicaron al algoritmo que opere con una probabilidad 'x' de que los 'vecinos más cercanos' pertenezcan a la misma clase de tejidos. Esto fue importante para evitar los artefactos que crea la adquisición de secuencias T1, en especial en las regiones más periféricas del lóbulo temporal (Poldrack, 2007). Luego se realizó un control de calidad de las segmentaciones realizadas. Implicó volúmenes y escalamiento proporcional. En el control de calidad se visualizó en un corte de 1 mm que las imágenes segmentadas correspondieran a cada tejido estructural.
Se controló la homogeneidad de la muestra mediante una matriz de covarianza entre los archivos obtenidos, para analizar qué espacios cerebrales podrían ser excluidos del análisis por altas diferencias estructurales. En este caso, se trató de personas controles con una alta homogeneidad en edad, nivel educativo, del mismo sexo. Por lo tanto, no fue necesario descartar ningún participante. Por último, se realizó el proceso de suavización de las imágenes segmentadas de sustancia gris, utilizando un valor de [8 8 8] mm de anchura entera en la altura media del núcleo Gaussiano de suavización.
Imágenes con pesos de difusión
El post-procesamiento fue realizado offline utilizando Matlab 7.10.0 y ExploreDTI v4.8.3 (Leemans et al., 2009). Las imágenes adquiridas de secuencia T1 y DTI fueron convertidas a formato 'Niifti', y se generó la reconstrucción de todas las imágenes en una única matriz. La estimación de los tensores de difusión se realizó mediante el método 'linear'. Se realizó corrección de movimiento por sujeto sobre el espacio cerebral nativo. Sobre la matriz reconstruida se generó una tractografía de cerebro entero, excluyendo fibras de longitud menor a 10 mm y mayores a 500 mm; se estableció un rango de AF entre .2 y 1, y se utilizó un método de interpolación 'linear'.
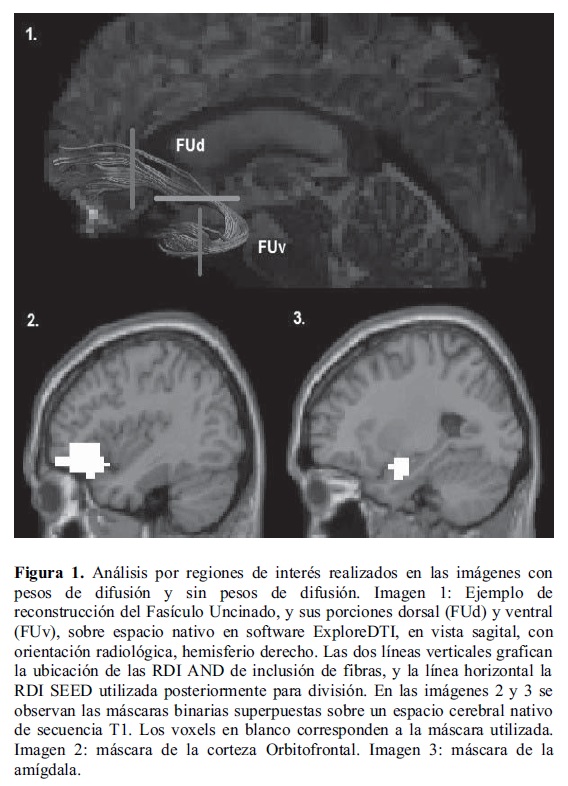
2.5. Análisis por regiones de interés
Imágenes sin pesos de difusión
Para analizar el volumen de las estructuras de sustancia gris de interés, se generaron máscaras binarias en la caja de herramientas XjView (http://www.alivelearn.net/xjview) que incluyeran los voxels correspondientes a la amígdala izquierda y derecha, y a la corteza orbitofrontal izquierda y derecha (una máscara por cada hemisferio de cada estructura, cuatro máscaras independientes), y excluyeran los voxels restantes de la imagen segmentada del análisis. Las máscaras fueron ingresadas en la función 'get_totals' desarrollada por Ridgway (http://www0.cs.ucl.ac.uk/staff/g.ridgway/vbm/) en MATLAB 7.10.0 para calcular el volumen en mililitros de una estructura cerebral.
Imágenes con pesos de difusión
Luego de generar la tractografía de cerebro entero, se realizó un análisis de Regiones de Interés (RDI) con el objetivo de aislar grupos de fibras específicos (fascículos). Se utilizaron RDI con conectores lógicos AND (para incluir fibras en el análisis). Cada análisis de RDI se realizó en los dos hemisferios cerebrales sobre el espacio nativo de cada sujeto. Para ubicar las RDI sobre la matriz reconstruida y segmentar el Fascículo Uncinado (FU), se utilizó un atlas estandarizado de disecciones virtuales in vivo como referencia (Catani & Thiebaut de Schotten, 2008). El FU conecta regiones de la corteza prefrontal ventral y orbital con zonas mediales del sistema límbico y regiones anteriores del lóbulo temporal. Se utilizaron dos RDI AND coronales para su disección: una en la zona orbito-frontal y una en la zona temporal anterior. Posteriormente el FU fue dividido en sus porciones dorsal y ventral (Von Der Heide, Skipper, Klobusicky, & Olson, 2013), mediante la herramienta 'splitter tool' de ExploreDTI, colocando una RDI SEED axial para su división. De cada porción del FU, se extrajo el valor de la anisotropía fraccional con su respectiva desviación estándar.
La Figura 1 muestra las RDI utilizadas para segmentar las estructuras de sustancia gris y sustancia blanca de interés en el presente estudio.
2.6. Análisis de datos
Los valores extraídos de los análisis de regiones de interés de las imágenes obtenidas de las secuencias de resonancia magnética fueron posteriormente relacionados mediantes técnicas estadísticas multivariadas. Se realizaron correlaciones bivariadas de Pearson y modelos de regresión múltiple entre los datos de volúmenes de sustancia gris y anisotropía fraccional. Posteriormente, se realizó un análisis de mediación mediante la macro Sobel (Preacher & Hayes, 2004) para conocer los efectos directos e indirectos de la variable independiente (amígdala total, amígdala izquierda, amígdala derecha) sobre la variable dependiente (orbitofrontal, izquierda y derecha). Se considera mediador al grado de explicación que una variable aporta al efecto de un predictor sobre la variable criterio.
3. RESULTADOS
3.1. Estadísticos descriptivos
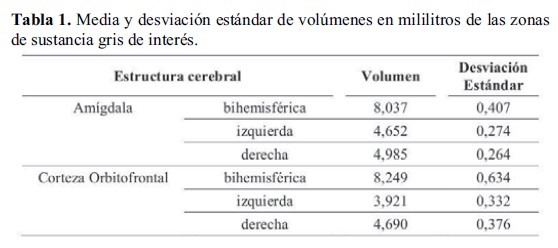
En la Tabla 1 se presenta el volumen de las estructuras cerebrales de interés, expresadas en mililitros. Se destaca que la suma de las dos máscaras bihemisféricas no arroja como resultado exacto el valor de la estructura total unificada, ya que se trata de un nuevo diseño de máscara que implica alguna diferencia mínima en los voxels que abarca y excluye.
En la Tabla 2 los descriptivos de anisotropía fraccional de las dos porciones del fascículo uncinado, dorsal y ventral.

3.2. Matriz de correlaciones
En la Tabla 3 se presentan las correlaciones entre los valores de la amígdala, la corteza orbitofrontal y la conectividad del fascículo uncinado. Entre las correlaciones calculadas se destacan aquellas significativas. Las relaciones bihemisféricas entre estructuras cerebrales similares se da por descontada. En el hemisferio derecho se produjo una interesante relación entre la amígdala, la corteza orbitofrontal y el fascículo uncinado (conectividad) en su parte dorsal.
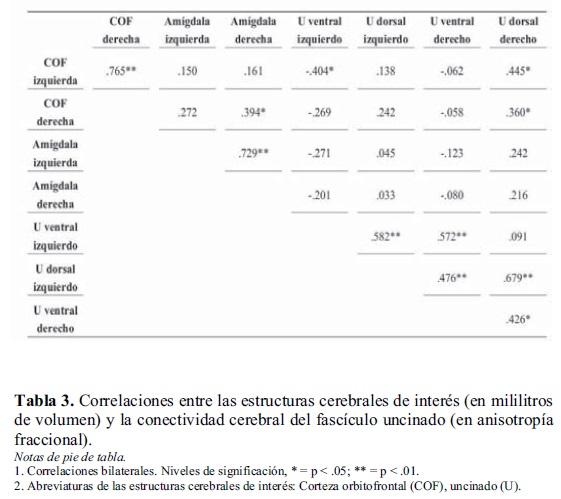
Para profundizar en estas relaciones se realizó un análisis de regresión múltiple. Se ingresaron como variables independientes la amígdala y la parte dorsal del fascículo uncinado, y como variable dependiente el volumen de la corteza orbitofrontal. Se encontró un modelo significativo de predicción [F=4,437; p< .05; gl= 2, 29; R2= .234], los valores de β fueron .332 para el volumen de la amígdala y .288 para la anisotropía fraccional de la parte dorsal del fascículo uncinado. Cuando se analizó la colinealidad entre ambas variables, se observó que hubo alta tolerancia. Eso significa que cada variable tuvo un aporte independiente a la explicación de la variable dependiente y contribuyó de forma significativa al modelo. Un resultado llamativo fue que la corteza orbitofrontal izquierda se relacionó de forma negativa significativa con la parte ventral del fascículo uncinado izquierdo y de forma positiva significativa con la parte dorsal del fascículo uncinado derecho.
En las correlaciones entre las porciones dorsal y ventral del hemisferio izquierdo y derecho del fascículo uncinado, se aprecia que las únicas porciones que no están relacionadas son las mencionadas: el fascículo uncinado ventral izquierdo y el dorsal derecho. Sin embargo, el volumen de la corteza orbitofrontal izquierda se relaciona de forma inversa con las dos partes. En función de estos resultados, se realizó un análisis de regresión múltiple. Se colocó como variable dependiente el volumen de la corteza orbitofrontal izquierda y como variables independientes la anisotropía fraccional del fascículo uncinado ventral izquierdo y dorsal derecho. Se obtuvo un modelo altamente predictivo [F=9,558; p= .001; gl= 2,29; R2= .401]. El valor de β de la porción ventral en el hemisferio izquierdo fue - .448 y en la porción dorsal del hemisferio derecho de .487. La tolerancia fue sustantiva y ambas variables ingresaron en el modelo como predictoras (p< .01).
3.3. Estadísticos inferenciales
En el análisis de mediación mediante la macro Sobel (Preacher & Hayes, 2004), se ingresó como mediador la anisotropía fraccional del fascículo uncinado, en sus diferentes porciones. El procedimiento respetó lo siguiente: cuando la variable independiente fue la amígdala total, la variable dependiente fue cada una de las porciones de la la corteza orbitofrontal, se analizó una cada vez. Luego se respetaron los hemisferios de pertenencia de cada estructura, por ejemplo cuando se ingresó la variable independiente amígdala izquierda, se colocó como variable dependiente la corteza orbitofrontal izquierda y como posible mediador los valores de anisotropía del fascículo uncinado izquierdo. Esto se llama principio de conectividad estructural (Sporns, Chialvo, Kaiser, & Hilgetag, 2004).
Las medidas que se obtuvieron fueron el efecto directo, el efecto indirecto y el valor de mediación. El test de Sobel se basa en la normalidad de la distribución de las variables. Tal requisito fue comprobado mediante los análisis de curtosis y asimetría. Para establecer que existe una mediación se debe hacer el procedimiento inverso, colocar como variable independiente la que se conjetura dependiente. En ese caso, no debería haber significación si habría una mediación en el sentido predicho. Este procedimiento se utiliza de control. Cabe recordar que los valores de anisotropía fraccional no indican la direccionalidad de la conectividad. La conectividad estructural amígdala- corteza orbitofrontal se presenta unidireccional en nuestra hipótesis de trabajo. Responde a los modelos de procesamiento de regulación de las emociones más asentados (Goldin, McRae, Ramel, & Gross, 2008). Sin embargo, hay evidencias de que la conectividad tiene características bidireccionales (Kalisch, 2009).
En este caso se utilizaron mediaciones simples (Baron & Kenny, 1986). Teniendo en cuenta que el test de Sobel necesita de muestras grandes para resultar lo suficientemente conservador, se utilizó el procedimiento Bootstrapping de 5000 remuestreos, tal como sugieren autores de referencia (Preacher & Hayes, 2004, 2008). Además, el remuestreo mediante Bootstrapping permitió eliminar los defectos de la propuesta de Baron & Kenny (Baron & Kenny, 1986), que parten del supuesto de que la variable independiente debe predecir de forma significativa la variable dependiente para que pueda haber efectos indirectos. Para ver más detalles del aporte del Bootstrapping y sus aportes al test de Sobel se recomienda leer Preacher & Hayes (Preacher & Hayes, 2008).
Mediación del fascículo uncinado izquierdo
Cuando se ingresó la amígdala izquierda como variable independiente, la corteza orbitofrontal izquierda como variable dependiente y la anisotropía fraccional del fascículo uncinado dorsal como mediador, no hubo efectos de ningún tipo.
En cambio, cuando se introdujo como variable mediadora el fascículo uncinado ventral, se encontraron efectos significativos sobre el volumen de la corteza orbitofrontal izquierda [b(YM.X) Coef. B = -6,8078 (ds= 3,0576) t=-2,2265 p< .05; R2= .163]. Estos efectos indicaron que a mayor anisotropía fraccional del fascículo uncinado izquierdo ventral, menor el volumen de la corteza orbitofrontal izquierda, siendo controlado el volumen de la amígdala izquierda. El significado de este resultado fue que la conectividad de la parte ventral uncinada determinó en parte el volumen de la corteza orbitofrontal izquierda, de forma independiente al volumen de la amígdala. El test de Sobel dio negativo con ambas partes del fascículo uncinado. De tal modo se puede decir que no mediatiza la relación entre el volumen de la amígdala y el de la corteza orbitofrontal.
Mediación del fascículo uncinado derecho
Se encontró un efecto total del volumen de la amígdala sobre el volumen de la corteza orbitofrontal [b (YX) Coef. B= 0,560 (ds= 0,238) t= 2,234 p< .05 R2= .155]. Cuando se ingresó la parte ventral del fascículo uncinado tampoco se encontraron efectos de mediación y el test de Sobel volvió a ser negativo. Esto implicó que la parte dorsal del fascículo uncinado no fue mediador de la relación positiva entre el volumen de la amígdala y el volumen de la corteza orbital prefrontal.
4. DISCUSIÓN
Los resultados indicaron que la lateralización hemisférica es una cuestión central en la relación entre la amígdala y la corteza orbitofrontal. Autores que investigaron esta relación en procesos de toma de decisiones destacaron este aspecto (Gaznick, Bechara, & Tranel, 2014). Postularon que la función principal era cumplida por la corteza orbitofrontal derecha y disminuyeron la relevancia de la parte correspondiente al hemisferio izquierdo. Esto se relaciona con la prevalencia de las emociones negativas en el hemisferio derecho (Nijboer & Jellema, 2012). Incluso, la técnica terapéutica conocida como 'Rapid Eye Movements Desensitization' (Shapiro, 2014) se basa en la diferencia hemisférica en el procesamiento de emociones. Se trata de evocar recuerdos, mantenerlos en el espacio de la memoria de trabajo en conexión con la memoria episódica y fomentar el pasaje de tratamiento de un hemisferio a otro. Parten de la hipótesis de que algunos recuerdos 'obstruidos' y patológicos no pueden ser tratados por la pátina más positiva del hemisferio izquierdo.
Buena parte de la responsabilidad de que esto ocurra es resultado del papel de la amígdala derecha. La recepción y emisión de estimulación emocional negativa tiene evidencias sólidas. En el análisis de mediación que realizamos, se encontraron evidencias de un efecto directo del volumen de la amígdala sobre el volumen de la corteza orbitofrontal derecha. Esto no ocurrió en el hemisferio izquierdo. En este último hemisferio, el gradiente de conectividad del fascículo uncinado en su parte ventral determinó de forma negativa el volumen de la corteza orbitofrontal izquierda. Esto significó que a menor anisotropía en la parte límbica del fascículo uncinado, mayor era el tamaño de la corteza orbitofrontal.
Este resultado incrementó de forma notable su interés por lo siguiente: cuando se analizó la matriz de correlaciones entre las cuatro partes del fascículo uncinado, en consideración bihemisférica, solo una de las cuatro correlaciones no fue significativa (la que vinculaba la parte dorsal derecha con la parte ventral izquierda). La parte ventral conecta el polo temporal, relacionado con la abstracción semántica (Whitney, Kirk, O'Sullivan, Lambon Ralph, & Jefferies, 2011), con la corteza orbitofrontal. Sin embargo, la misma matriz de correlaciones indicó que el tamaño de la corteza orbitofrontal izquierda, desacreditada en su funcionalidad en comparación con la porción derecha, estaba determinada en su volumen por un potente tamaño del efecto compuesto de forma negativa por la conectividad ventral uncinada mencionada y de forma positiva por la parte dorsal uncinada derecha.
Entre ambas porciones predictoras se halló una tolerancia elevada, que resultó consecuente a su nula correlación entre sí. De forma concreta, cuando el uncinado derecho en su parte superior (semántico- emocional) tenía una alta conectividad y al mismo tenía una baja conectividad en su parte inferior izquierda, se pudo hallar un modelo que predijo de forma significativa y elevada para la cantidad de casos el tamaño de la corteza orbitofrontal izquierda.
Esto podría indicar que la parte más ligada a las emociones negativas, el hemisferio derecho, cuando tenía una alta conectividad uncinada superior, e iba acompañada de una baja conectividad inferior uncinada izquierda, provocaría que el rol de filtro de emociones negativas de la corteza orbitofrontal izquierda se incremente. Estos datos resultaron novedosos y de interés, por lo que se propuso iniciar una réplica del estudio para buscar mayores evidencias de lo hallado. De encontrarse más apoyo, el rol del fascículo uncinado como mediador estaría fuertemente centrado en su división dorsal y ventral. También el papel de la corteza orbitofrontal izquierda resultaría de mayor interés en función de la conectividad del uncinado en las porciones señaladas, quizá como un componente de complemento de filtro.
Los modelos de ansiedad y depresión tienen en cuenta la conectividad amígdalo- orbitofrontal. En la parte límbica- amigdalina se produce la 'pátina' emocional que resulta regulada en primera instancia por la corteza orbitofrontal (Buhle et al., 2013). En esto, el papel del etiquetamiento lingüístico es muy potente (Barrett et al., 2007). En el estudio realizado, la causalidad amigdalina- orbitofrontal, en volumen, sumó evidencias, tal como se preveía por la literatura, para el hemisferio derecho. No obstante, este estudio podría aportar índices para nuevos estudios que complejicen la mediación del fascículo uncinado y que permitan comprender el rol (hasta nuestro conocimiento no publicado) sobre la complejidad dependiente del volumen de la corteza orbitofrontal izquierda. Los modelos de mediación más complejos, como la técnica Mediation (Preacher & Hayes, 2008; Romer et al., 2009), servirían para nuestras futuras investigaciones. Se subraya, entonces, la importancia de combinar diferentes secuencias de Resonancia Magnética, en este caso imágenes por tensores de difusión y morfometría basada en voxels (J Ashburner & Friston, 2000; Horwitz, Friston, & Taylor, 2000; Ystad et al., 2011).
Referencias
Ashburner, J. (2007). A fast diffeomorphic image registration algorithm. Neuroimage, 38(1), 95–113. doi:10.1016/j.neuroimage.2007.07.007
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry--the methods. NeuroImage, 11(6 Pt 1), 805–21. doi:10.1006/nimg.2000.0582
Baron, R. M., & Kenny, D. A. (1986). The moderator-mediator variable distinction in social psychological research: conceptual, strategic, and statistical considerations. Journal of Personality and Social Psychology, 51(6), 1173–1182. doi:10.1037/0022-3514.51.6.1173
Barrett, L. F., Lindquist, K. A., & Gendron, M. (2007). Language as context for the perception of emotion. Trends in Cognitive Sciences, 11(8), 327–32. doi:10.1016/j.tics.2007.06.003
Bechara, A., Damasio, H., Tranel, D., & Damasio, A. R. (2005). The Iowa Gambling Task and the somatic marker hypothesis: some questions and answers. Trends in Cognitive Sciences, 9(4), 159–64. doi:10.1016/j.tics.2005.02.002
Bosch, B., Arenaza-Urquijo, E. M., Rami, L., Sala-Llonch, R., Junqué, C., Solé-Padullés, C., … Bartrés-Faz, D. (2012). Multiple DTI index analysis in normal aging, amnestic MCI and AD. Relationship with neuropsychological performance. Neurobiology of Aging, 33(1), 61–74. doi:10.1016/j.neurobiolaging.2010.02.004
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C., Kober, H., … Ochsner, K. N. (2013). Cognitive reappraisal of emotion: a meta-analysis of human neuroimaging studies. Cerebral Cortex, bht154. doi:10.1093/cercor/bht154
Cardinal, R. N., Parkinson, J. a, Hall, J., & Everitt, B. J. (2002). Emotion and motivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neuroscience and Biobehavioral Reviews, 26(3), 321–52. doi:10.1016/S0149-7634(02)00007-6
Catani, M., & Thiebaut de Schotten, M. (2008). A diffusion tensor imaging tractography atlas for virtual in vivo dissections. Cortex, 44(8), 1105–32. doi:10.1016/j.cortex.2008.05.004
Damasio, H., Tranel, D., Grabowski, T., Adolphs, R., & Damasio, a. (2004). Neural systems behind word and concept retrieval. Cognition, 92(1-2), 179–229. doi:10.1016/j.cognition.2002.07.001
Eluvathingal, T. J., Chugani, H. T., Behen, M. E., Juhász, C., Muzik, O., Maqbool, M., … Makki, M. (2006). Abnormal brain connectivity in children after early severe socioemotional deprivation: a diffusion tensor imaging study. Pediatrics, 117(6), 2093–100. doi:10.1542/peds.2005-1727
Eysenck, M. W., Derakshan, N., Santos, R., & Calvo, M. G. (2007). Anxiety and cognitive performance: attentional control theory. Emotion (Washington, D.C.), 7(2), 336–53. doi:10.1037/1528-3542.7.2.336
Feldmann, A., Illes, Z., Kosztolanyi, P., Illes, E., Mike, A., Kover, F., … Nagy, F. (2008). Morphometric changes of gray matter in Parkinson's disease with depression: A voxel-based morphometry study. Movement Disorders, 23(1), 42–46. doi:10.1002/mds.21765
Gaznick, N., Bechara, A., & Tranel, D. (2014). Hemispheric side of damage influences sex-related differences in smoking cessation in neurological patients. Journal of Clinical and Experimental Neuropsychology, 36(5), 551–558. doi:10.1080/13803395.2014.915012
Goldin, P. R., McRae, K., Ramel, W., & Gross, J. J. (2008). The neural bases of emotion regulation: reappraisal and suppression of negative emotion. Biological Psychiatry, 63(6), 577–86. doi:10.1016/j.biopsych.2007.05.031
Highley, J. R., Walker, M. A., Esiri, M. M., Crow, T. J., & Harrison, P. J. (2002). Asymmetry of the uncinate fasciculus: a post-mortem study of normal subjects and patients with schizophrenia. Cerebral Cortex, 12(11), 1218–1224. doi:10.1093/cercor/12.11.1218
Horwitz, B., Friston, K. J., & Taylor, J. G. (2000). Neural modeling and functional brain imaging: an overview. Neural Networks : The Official Journal of the International Neural Network Society, 13(8-9), 829–46. doi:10.1016/S0893-6080(00)00062-9
Kalisch, R. (2009). The functional neuroanatomy of reappraisal: time matters. Neuroscience and Biobehavioral Reviews, 33(8), 1215–26. doi:10.1016/j.neubiorev.2009.06.003
Kerr, A., & Zelazo, P. D. (2004). Development of "hot" executive function: the children's gambling task. Brain and Cognition, 55(1), 148–57. doi:10.1016/S0278-2626(03)00275-6
Leemans, A., Jeurissen, B., Sijbers, J., & Jones, D. K. (2009). ExploreDTI: A Graphical Toolbox for Processing, Analyzing, and Visualizing Diffusion MR Data. In 17th Annual Meeting of Intl Soc Mag Reson Med (p. 3537). Hawaii, USA. [ Links ]
Lochner, C., Fouché, J.-P., du Plessis, S., Spottiswoode, B., Seedat, S., Fineberg, N., … Stein, D. J. (2012). Evidence for fractional anisotropy and mean diffusivity white matter abnormalities in the internal capsule and cingulum in patients with obsessive-compulsive disorder. Journal of Psychiatry & Neuroscience : JPN, 37(3), 193–9. doi:10.1503/jpn.110059
Nijboer, T. C. W., & Jellema, T. (2012). Unequal impairment in the recognition of positive and negative emotions after right hemisphere lesions: a left hemisphere bias for happy faces. Journal of Neuropsychology, 6(1), 79–93. doi:10.1111/j.1748-6653.2011.02007.x
Ochsner, K. N., & Gross, J. J. (2005). The cognitive control of emotion. Trends in Cognitive Sciences, 9(5), 242–9. doi:10.1016/j.tics.2005.03.010
Pessoa, L., & Adolphs, R. (2010). Emotion processing and the amygdala: from a "low road" to "many roads" of evaluating biological significance. Nature Reviews Neuroscience, 11(11), 773–783. doi:10.1038/nrn2920
Phan, K. L., Orlichenko, A., Boyd, E., Angstadt, M., Coccaro, E. F., Liberzon, I., & Arfanakis, K. (2010). Preliminary Evidence of White Matter Abnormality in the Uncinate Fasciculus in Generalized Social Anxiety Disorder. Biological Psychiatry, 66(7), 691–694. doi:10.1016/j.biopsych.2009.02.028
Poldrack, R. A. (2007). Region of interest analysis for fMRI. Social Cognitive and Affective Neuroscience, 2(1), 67–70. doi:10.1093/scan/nsm006
Preacher, K. J., & Hayes, A. F. (2004). SPSS and SAS procedures for estimating indirect effects in simple mediation models. Behavior Research Methods, Instruments, & Computers, 36(4), 717–731. doi:10.3758/BF03206553
Preacher, K. J., & Hayes, A. F. (2008). Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behavior Research Methods, 40(3), 879–891. doi:10.3758/BRM.40.3.879
Romer, D., Betancourt, L., Giannetta, J. M., Brodsky, N. L., Farah, M., & Hurt, H. (2009). Executive cognitive functions and impulsivity as correlates of risk taking and problem behavior in preadolescents. Neuropsychologia, 47(13), 2916–26. doi:10.1016/j.neuropsychologia.2009.06.019
Shapiro, F. (2014). The Role of Eye Movement Desensitization and Reprocessing (EMDR) Therapy in Medicine: Addressing the Psychological and Physical Symptoms Stemming from Adverse Life Experiences. The Permanente Journal, 18(1), 71. doi:10.7812/TPP/13-098 [ Links ]
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C. E., … Behrens, T. E. J. (2006). Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. NeuroImage, 31(4), 1487–505. doi:10.1016/j.neuroimage.2006.02.024
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C. (2004). Organization, development and function of complex brain networks. Trends in Cognitive Sciences, 8(9), 418–425. doi:10.1016/j.tics.2004.07.008
Tottenham, N., Hare, T. A., Quinn, B. T., McCarry, T. W., Nurse, M., Gilhooly, T., … Eigsti, I. (2010). Prolonged institutional rearing is associated with atypically large amygdala volume and difficulties in emotion regulation. Developmental Science, 13(1), 46–61. doi:10.1111/j.1467-7687.2009.00852.x
Van Hecke, W., Leemans, A., Sage, C. a, Emsell, L., Veraart, J., Sijbers, J., … Parizel, P. M. (2011). The effect of template selection on diffusion tensor voxel-based analysis results. NeuroImage, 55(2), 566–73. doi:10.1016/j.neuroimage.2010.12.005
Van Marle, H. J. F., Tendolkar, I., Urner, M., Verkes, R. J., Fernández, G., & van Wingen, G. (2011). Subchronic duloxetine administration alters the extended amygdala circuitry in healthy individuals. NeuroImage, 55(2), 825–831. doi:10.1016/j.neuroimage.2010.12.051
Vasic, N., Walter, H., Höse, A., & Wolf, R. C. (2008). Gray matter reduction associated with psychopathology and cognitive dysfunction in unipolar depression: a voxel-based morphometry study. Journal of Affective Disorders, 109(1), 107–116. doi:10.1016/j.jad.2007.11.011
Von Der Heide, R. J., Skipper, L. M., Klobusicky, E., & Olson, I. R. (2013). Dissecting the uncinate fasciculus: disorders, controversies and a hypothesis. Brain : A Journal of Neurology, 136(6), 1692–1707. doi:10.1093/brain/awt094
Whitney, C., Kirk, M., O'Sullivan, J., Lambon Ralph, M. A., & Jefferies, E. (2011). The neural organization of semantic control: TMS evidence for a Distributed Network in Left Inferior Frontal and Posterior Middle Temporal Gyrus. Cerebral Cortex, 21(5), 1066–75. doi:10.1093/cercor/bhq180
Ystad, M., Hodneland, E., Adolfsdottir, S., Haász, J., Lundervold, A. J., Eichele, T., & Lundervold, A. (2011). Cortico-striatal connectivity and cognition in normal aging: A combined DTI and resting state fMRI study. NeuroImage, 55(1), 24–31. doi:10.1016/j.neuroimage.2010.11.016
 Dirección para correspondencia
Dirección para correspondencia
Julián Marino
25 de Mayo 912, piso 1º departamento B, Barrio General Paz
CP 5010, Ciudad de Córdoba, Córdoba, Argentina
E-mail: jmarino@psyche.unc.edu.ar
Artículo recibido: 09/03/2015
Artículo revisado: 10/08/2015
Artículo aceptado: 17/08/2015
Fuentes de Financiación: Centro Privado de Tomografía Computada Córdoba S.A. – Fundación Oulton, Córdoba, Argentina; Secretaría de Ciencia y Tecnología de la Universidad Nacional de Córdoba