











 (pdf)
(pdf)




 Permalink
PermalinkRevista de Etologia
ISSN 1517-2805
Artigo
Avaliação do comportamento de cutias Dasyprocta azarae e Dasyprocta leporina (Rodentia: Dasyproctidae) em cativeiro e semicativeiro em parques urbanos de Curitiba, Paraná, Brasil
Behavioral evaluation of captive and semi-captive agoutis Dasyprocta azarae and Dasyprocta leporina (Rodentia: Dasyprotidae), on urban parks of Curitiba, Paraná, Brazil
Silvane Krul KaiserI; Teresa Cristina Castellano MargaridoII; Marta Luciane FischerI
I Universidade Católica do Paraná
II Prefeitura Municipal de Curitiba
RESUMO
O conhecimento do repertório comportamental de cutias cativas e livres é fundamental para o melhoramento de técnicas de manejo. Observações em cativeiro foram realizadas no criadouro científico de animais silvestres pertencente ao Museu de História Natural do Capão da Imbuia (MHNCI) e no Parque Municipal Passeio Público, enquanto observações em semicativeiro foram realizadas na área livre no MHNCI e no Parque Jardim Botânico. O etograma de Dasyprocta azarae (Lichtenstein, 1923) e Dasyprocta leporina (Linnaeus, 1758) foi realizado em 120 horas, caracterizando 78 atos comportamentais, quantificados em 96 horas através dos métodos animal focal e scan. As frequências foram analisadas através do teste do qui-quadrado, que revelou diferenças entre os ambientes. Nos semicativeiros, as frequências de alimentação, interações agonísticas, interações reprodutivas e comportamentos de cuidados parentais foram mais elevadas, indicando a disputa pelos recursos, reflexo na reprodução e cuidados com a prole. Em cativeiro, as frequências elevadas com comportamentos autodirigidos, atos exploratórios e relações entre coespecíficos sugerem a falta de estimulação, bem como a proximidade forçada entre os indivíduos. Portanto, existe uma diferenciação na frequência dos comportamentos exibidos, possivelmente devido às diferenças entre os ambientes, que podem estar relacionadas com as condições de manutenção dessas espécies.
Palavras-chave: Dasyprocta sp, etograma, cativeiro, padrão de atividade, bem-estar animal.
ABSTRACT
The knowledge of the behavioral repertoire of captive and free-ranging agoutis is essential for the improvement of management techniques. Observations were made in the field of reproduction of the Museu de História Natural do Capão da Imbuia (MHNCI) and in the Passeio Público, while observations in semi-captivity were held in the MHNCI and the Jardim Botânico. The ethogram of Dasyprocta azarae (Lichtenstein, 1923) and Dasyprocta leporina (Linnaeus, 1758) was carried out through in 120h, characterizing 78 behavioral acts, quantified in 96h by the methods focal animal and scan. The frequencies were analyzed using the chi-square test which revealed differences between the environments. In the semi-captivity the frequencies of feeding, agonistics interactions, reproductive interactions and parental care were higher indicating the resources competition and reflection on the reproduction and care for their offspring. In captivity the high incidence of self-directed behaviors, exploratory acts and coespecifics relations emphasize the lack of stimulation and the forced proximity among the animals. Therefore, there are differences in the frequency of behaviors exhibited, possibly due to differences between environments, which may be related to the maintenance conditions of these species.
Keywords: Dasyprocta sp, ethogram, captivity, activity pattern, urban parks, animal welfare.
A aplicação de atitudes éticas com animais abrigados nos zoológicos, principalmente quanto às condições de cativeiro, tem despertado a atenção da comunidade científica (Almeida, 2004). As circunstâncias de manutenção e os recintos nem sempre permitem a expressão de comportamentos naturais do animal (Bosso, 2007), devido ao ambiente de cativeiro ser limitado em tamanho, imutável, previsível, sem estímulos, em que o animal não tem controle sobre este (Hashimoto, 2008), podendo levá-los a exibir comportamentos como automutilação, canibalismo, agressividade excessiva e dificuldades de movimentação, bem como crescimento anormal e doenças, indicando condições de baixo grau de bem-estar (Broom & Molento, 2004). Logo, a manutenção dos animais silvestres em cativeiro requer o conhecimento da sua biologia, para que assim os seus comportamentos possam ser interpretados e usados como indicativos de bem-estar decorrentes das condições oferecidas.
O gênero Dasyprocta compreende nove espécies: Dasyprocta aurea Cope, 1889, D. catrinae Thomas, 1917, D. fuliginosa Wagler, 1832, D. croconota Wagler, 1831, D. nigriclunis Osgood, 1916, D. prymnolopha Wagler, 1841, D. aguti Linnaeus, 1766, D. azarae Lichtenstein, 1823 e D. leporina Linnaeus, 1758 (Iach-Ximenes, 1999). Esses roedores silvestres apresentam uma vasta distribuição na América Neotropical, ocorrendo em uma grande variedade de habitats (Eisenberg & Redford, 1992; Novak, 1999; Reis et al., 2006; Santos, 2005).
A cutia D. leporina ocorre na bacia amazônica ao sul do rio Amazonas, entre os rios Madeira e Tocantins, e no leste do Brasil, nos estados da Paraíba, Pernambuco, Bahia, Espírito Santo, Rio de Janeiro e São Paulo (Reis et al.,2006). Já a espécie D. azarae, é uma das mais distintas do gênero (Lange, 1998), distribuindo-se do Tocantins ao leste dos Andes (Lange, 1998; Reis et al., 2006). Habitam florestas pluviais (Amazônia e Floresta Atlântica), florestas semidecíduais, Cerrado e Caatinga, sendo a distribuição associada a cursos de água (Reis et al., 2006). Ambas as espécies possuem hábitos diurnos, porém podem estender a atividade quando submetidas à interferência antrópica extrema ou devido à alta pressão de predação (Santos, 2005). Alimentam-se de frutos, sementes, raízes e vegetais, atingem a maturidade sexual aos seis meses, estabelecendo pares monogâmicos (Santos, 2005; Smythe, 1978).
A necessidade de manutenção de cutias em cativeiro para criações comerciais e em zoológicos para conservação ex situ, torna-as um importante objeto de estudo comparativo de seu comportamento em diferentes situações, visando subsidiar planos de ação para obter condições que proporcionem bem-estar. Logo, questiona-se se há diferenças no comportamento exibido por animais cativos e semicativos, e entre as espécies D. leporina e D. azarae. Supõe-se que comportamentos de animais cativos e semicativos sejam diferentes em relação às atividades e relações sociais, pontos importantes para o desenvolvimento da espécie. Portanto, o conhecimento deste aspecto possibilita uma melhor forma de manutenção desses animais em ambos os locais.
Assim, o objetivo do presente estudo foi avaliar as diferenças no comportamento de D. azarae e D. leporina, em ambiente de cativeiro e semicativeiro.
Método
Área de Estudo
A avaliação do comportamento de D. azarae e D. leporina foi realizada em quatro locais na cidade de Curitiba, no Estado do Paraná: no Museu de História Natural do Capão da Imbuia (MHNCI) em sua área aberta, no criadouro cientifico de animais silvestres pertencente também ao Museu, no Jardim Botânico (JB) e no Parque Muncicipal Passeio Público (PP).
O Museu de História Natural Capão da Imbuia compreende uma de área total de 34.800 m2, com uma área de mata natural caracterizada por um pequeno fragmento do bioma floresta ombrófila mista. Através do projeto “Animais Silvestres nas áreas verdes de Curitiba”, da Prefeitura Municipal de Curitiba com o apoio da Fundação O Boticário de Proteção à Natureza, desenvolveram-se criação e pesquisa de animais silvestres, repovoando a área do Museu com a espécie D. azarae. Atualmente, ocorrem neste local cerca de 60 indivíduos, que são mantidos pela própria vegetal nativa, bem como por frutas e verduras fornecidas pelos funcionários responsáveis pela manutenção dos animais (Lange, 1998).
O Criadouro Científico de Animais Silvestres do MHNCI é constituído por um conjunto de 10 recintos, dispostos em duas séries de cinco, com um corredor de serviço no centro, ocupando uma área de 240m2 (Lange, 1998). Cada recinto mede aproximadamente 20 m2, sendo que dois deles contêm, no máximo, quatro animais, sendo dois casais em que os machos são castrados. Os demais foram divididos ao meio, abrigando de um a dois indivíduos coespecíficos. Compõem os recintos manilhas dispostas nas laterais, que servem como abrigo e pontos de fuga, sendo uma para cada indivíduo. Também existe uma piscina de areia, circundada por pedras e coberta por folhiço. A população fundadora é proveniente de instituições do Paraná, oficialmente O Criadouro Científico de Animais Silvestres do MHNCI é constituído por um conjunto de 10 recintos, dispostos em duas séries de cinco, com um corredor de serviço no centro, ocupando uma área de 240m2 (Lange, 1998). Cada recinto mede aproximadamente 20 m2, sendo que dois deles contêm, no máximo, quatro animais, sendo dois casais em que os machos são castrados. Os demais foram divididos ao meio, abrigando de um a dois indivíduos coespecíficos. Compõem os recintos manilhas dispostas nas laterais, que servem como abrigo e pontos de fuga, sendo uma para cada indivíduo. Também existe uma piscina de areia, circundada por pedras e coberta por folhiço. A população fundadora é proveniente de instituições do Paraná, oficialmente registradas junto ao IBAMA. O cativeiro abriga atualmente 22 indivíduos da espécie D. azarae. Os animais são alimentados no início da manhã com ração comercial balanceada e milho, e no final da tarde com abóbora, batata-doce, cenoura, aipim, pinhão e banana (Lange, 1998).
O Jardim Botânico de Curitiba engloba um remanescente de floresta ombrófila mista, que representa 40% (71.000 m2) da área total do parque. O sub-bosque é denso devido ao processo de regeneração, possuindo dois estratos nítidos: um superior com árvores entre 1,5 a 20 m e um inferior com formação herbácea-arbustiva não ultrapassando 1,5 m. A camada de serrapilheira tem aproximadamente 2 cm de altura, e o sombreamento é constante durante todo o dia (Prefeitura Municipal de Curitiba, 2007). As cutias abrigadas no Jardim Botânico foram criadas no cativeiro do MHNCI e soltas na proporção sexual de 1:1, em 1994 (Lange, 1998). Os animais utilizam as trilhas e a vegetação fechada para locomoção, se alimentam das frutas do bosque (Araucaria angustifolia – pinhão; Syagrus romanzoffiana – coquinhos de jerivá; Luhea divaricata – fruto de açoite-cavalo; Eugenia uniflora – pitanga; Passiflora actinia – maracujá) (Lange, 1998), e também dos alimentos fornecidos pelos funcionários do Departamento de zoológico, com os mesmos itens alimentares para os animais do MHNCI, em quatro locais diferentes do bosque, sempre no período da manhã.
O Parque Municipal Passeio Público, localizado no centro de Curitiba, mantém um grupo de cutias da espécie D. leporina em um único recinto coletivo com uma área de 360m2, com um total de 27 indivíduos, sendo 13 machos (jovens e adultos) e 14 fêmeas (jovens e adultas). No interior do recinto há uma única fonte de água e alimento que são limpos todos os dias. A alimentação é fornecida duas vezes por dia, uma em cada período, e os alimentos que compõem a dieta são frutas (banana, maça, mamão), tubérculos (batata doce) e milho. Há 12 abrigos feitos de troncos de madeira, uma piscina de folhiço e galhos, que é cercada por troncos, e uma pequena construção de alvenaria que serve como ponto de fuga e local para manejo pelos médicos veterinários.
Procedimento
As observações em cativeiro foram realizadas no criadouro científico de animais silvestres pertencente ao MHNCI e no PP. As observações em semicativeiro foram feitas em área aberta do MHNCI e no JB.
O estudo foi realizado em duas etapas. Na primeira foi elaborado o etograma com a descrição qualitativa do repertório comportamental de D. azarae e D. leporina, através da observação dos animais pelo método Ad Libitum (Roll et al., 2006) por um total de 120 horas, em diferentes horários e condições ambientais. Os padrões motores foram agrupados em categorias comportamentais, e estas em grupos de comportamentos, sendo os padrões fotografados, permitindo a elaboração dos desenhos sob as fotos, semelhante aos mesmos traços realizado por Smythe (1978).
Na segunda etapa foi realizada a análise quantitativa, levando-se em consideração diferenças entre os locais, condições de manutenção e as espécies. Para as amostragens no MHNCI, tanto área aberta quanto o criadouro, e no JB estipulou-se 1 hora e 30 minutos de observação, utilizando o método animal focal (Roll et al., 2006). Para o PP, optou-se por aplicar em conjunto o método animal focal e scan (Roll et al., 2006). Primeiramente os animais foram amostrados individualmente por 5 minutos, sem intervalo entre as amostragens, durante 45 minutos. Em seguida aplicou-se o método de observação scan.durante 45 minutos, realizandos-se 16 amostragens de 2 minutos com intervalo de 1 minuto.
As amostragens nos locais foram realizadas em uma mesma semana, afim de se padronizar os registros, sendo os atos quantificados em 96 horas. As observações foram realizadas em dois períodos: pela manhã (7h30 às 9h30 e 10h às 11h30) e pela tarde (12h30 às 14h e 14h30 às 16h), durante 16 semanas, para homogeneizar as amostras.
Procedimentos legais
O presente estudo foi realizado de acordo com a legislação vigente no país, tendo autorização das unidades onde os estudos foram realizados e aprovação do Comitê de Ética no Uso de Animais CEUA - PUCPR (124.07).
Análise estatística
As semelhanças dos padrões motores em cada grupo entre as espécies foram analisadas através do índice de similaridade de Sorensen (1948), dados pela fórmula 2 * (n° comportamentos comuns entre as espécies dentro de cada grupo – excluindo comportamentos exclusivos)/nº de comportamentos para o 1º parâmetro (comportamentos para D. azarae) + nº de comportamentos para o 2º parâmetro (comportamento para D.leporina). Este mesmo índice foi utilizado para verificar as semelhanças entre os ambiente, 2 * (n° comportamentos comuns entre os dois ambientes dentro de cada grupo)/ n° de comportamentos executados em cativeiro para cada grupo + n° de comportamentos executados em semicativeiro para cada grupo.
Já as frequências dos comportamentos foram comparadas entre os períodos de observação, entre os diferentes ambientes, entre as espécies, através do teste do qui-quadrado.
Resultados
Descrição qualitativa do comportamento de D.azare e D.leporina
O etograma de D.azarae e D.leporina foi elaborado em 120 horas de observação, sendo composto por 5200 eventos, caracterizado por qualquer movimento do animal. Os 77 atos comportamentais foram agrupados em nove categorias, as quais compuseram três grupos de comportamentos: manutenção, exploração do ambiente e interações sociais.
Manutenção
O grupo manutenção foi caracterizado por 39 atos comportamentais (ac), agrupados nas categorias: alimentação, repouso e cuidados autodirigidos (Tabela 1).
Tabela 1. Padrões motores registrados nas categorias comportamentais dos grupos Manutenção, Exploração e Interações sociais de Dasyprocta azarae (•) e Dasyprocta leporina (*) em cativeiros e semicativeiros presentes no município de Curitiba, PR e atos de Dasyprocta sp já descritos em literatura (∆).

Durante a alimentação, o alimento foi manipulado com as patas dianteiras, transportado e ingerido. Todos os atos envolveram repouso e o posicionamento da cabeça acima ou abaixo da linha dorsal do corpo, dependendo da altura em que estava o alimento (Fig.1, Tabela 1). O repouso foi composto por atos comportamentais em que o animal podia estar deitado, sentado ou parado, envolvendo o apoio da região posterior ou lateral do corpo e dos membros locomotores. As patas dianteiras e traseiras podiam ficar estiradas ou dobradas abaixo do corpo (Fig.2, Tabela 1). Durante os cuidados autodirigidos, os animais foram registrados sacudindo o corpo, lambendo, coçando, esticando, defecando ou urinando, utilizando os membros locomotores, dentes e língua para lamber, coçar ou ajeitar os pelos (Fig.3, Tabela 1).
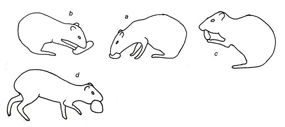
Figura 1. Categoria alimentação (Feeding category) a - abocanhar alimento; b – roer alimento; c – comer com patas dianteiras na boca; d – carregar alimento.
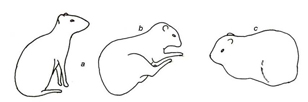
Figura 2. Categoria repouso (Rest category) a – sentado; b – deitado lateral; c – deitado sobre os membros.
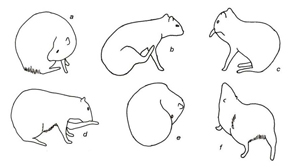
Figura 3. Categoria cuidados autodirigidos (Category corporal care) a – lamber dorso; b – coçar dorso com pata traseira; c – lamber pata dianteira; d – lamber pata traseira; e – lamber ventre; f – esticar o corpo.
Exploração do ambiente
Este grupo é caracterizado pela exibição de atos locomotores e exploratórios, sendo diferenciados em visualização do ambiente, farejar e exploração alimentar. Os atos foram desempenhados em repouso, com movimentação da cabeça e focinho para verificação do ambiente, e também em movimentação corporal, pela utilização das patas dianteiras para alterar aspectos ao seu redor, principalmente os relacionados à procura e guarda do alimento (Fig.4, Tabela 1).
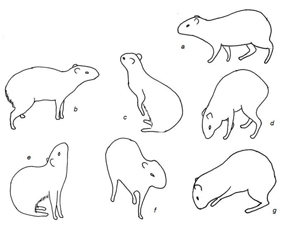
Figura 4. Categoria locomotora (Locomotive category) a – caminhada. Categoria exploração (Exploring category) b – observar ambiente; c – farejar apoiado nos membros traseiros; d – farejar superfície; e – farejar ao ar; f - Parado quadrúpede com pata dianteira elevada; g – cavar superfície.
Interações sociais
As interações sociais foram compostas de interações com coespecíficos, interações agonísticas, interações reprodutivas e cuidados parentais.
Diante de coespecíficos, os animais exibiram atos envolvendo reconhecimento e o grooming. Estes atos podem ser exibidos em momentos de repouso, em que um animal se aproxima do outro iniciando a ação ou sendo o estímulo para ação do outro, bem como podem estar relacionados com cuidados corporais dirigidos a outros indivíduos (Fig.5, Tabela 1). Nas interações agonísticas foram registradas a comunicação sonora, defesa de território, defesa de recursos e luta. Os atos envolveram locomoção, exploração do ambiente e contato com coespecíficos (Fig.6, Tabela 1). Nas interações reprodutivas, foram registrados atos relacionados ao cortejo, à cópula, e simulação de cópula. Na simulação de cópula, o macho, em posição de repouso (sentado), observou a fêmea, expôs seu órgão reprodutivo, realizando movimentos de cópula, porém sem a presença da fêmea (Fig.6, Tabela 1). Os cinco atos que envolveram cuidados com a prole foram relativos à alimentação, ao cuidado do filhote e atos interações filhote/mãe. A fêmea se posicionou deitada lateralmente, permitindo que o(os) filhotes mamassem. Em outras ocasiões, a fêmea que se alimentava permitiu que juvenis pegassem o alimento da sua boca; durante os cuidados com o filhote, a fêmea ficou sentada frente ao mesmo, protegendo de coespecíficos que tentavam se aproximar; já a interação filhote com a mãe se realizou subsequentemente ao ato da fêmea ceder alimento para filhote, quando este se locomove até uma curta distância, enterrando o item, retornando a fêmea e farejando sua boca, sendo estes atos de aproximar-se da mãe acompanhados de cabeçadas do filhote na região do pescoço (Fig.6, Tabela 1).
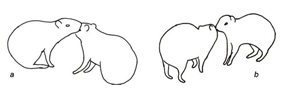
Figura 5. Categoria coespecíficos (Co-specific category) a – lamber outro indivíduo; b – farejar outro individuo.
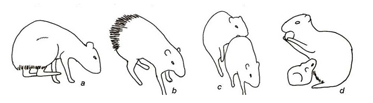
Figura 6. Categoria agonística (Agonistics category) a - Bater patas traseiras na superfície e b - piloereção; Categoria reprodutiva (Reproductive category) c - cópula; Categoria cuidados parentais (Parental Care category) d – amamentação.
O número de atos comportamentais dentro de cada comportamento foi similar entre as duas espécies nas interações sociais (0,96), manutenção (0,93) e exploração do ambiente (0,91). Comparando o comportamento de D. azarae em cativeiro e em semicativeiro, foi encontrada maior similaridade na manutenção (0,89) e na exploração do ambiente (0,86) quando comparadas com as interações sociais (0,55) (Tabela 1).
Análise quantitativa do comportamento de D.azare e D.leporina
As frequências dos grupos comportamentais, bem como das categorias, evidenciaram diferenças nos três locais de amostragem, entre os períodos de observação e entre as espécies.
Local
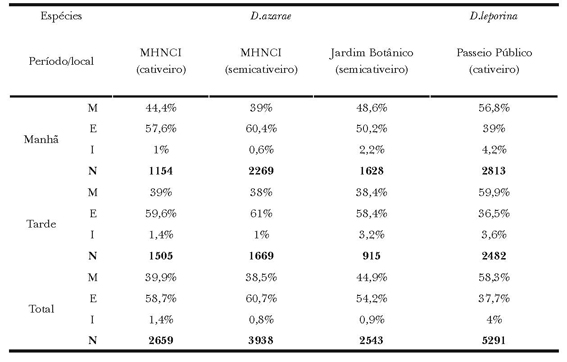
As frequências dos comportamentos de D. azarae mantidos em cativeiro não diferiram dos mantidos em semicativeiro no MHNCI. Ao comparar as frequências dos comportamentos entre os animais mantidos em semicativeiro no MHNCI e no JB (Χ2(2) = 75; P<0,01), a manutenção e as interações foram mais exibidas no JB, e a exploração no MHNCI. A comparação entre animais cativos no MHNCI e em semicativeiro no JB (Χ2(2) = 36,9; P<0,01), revelam que a exibição de atos de manutenção e interações sociais foram mais elevadas no JB e a exploração foi maior em cativeiro (Tabela 2).
Tabela 2. Frequência relativa (%) e total (N) de comportamentos dos grupos Manutenção (M), Exploração (E) e Interações (I) exibidos por D. azarae no Museu de História Natural do capão da Imbuia, no Jardim Botânico e por D. leporina no Passeio Público.
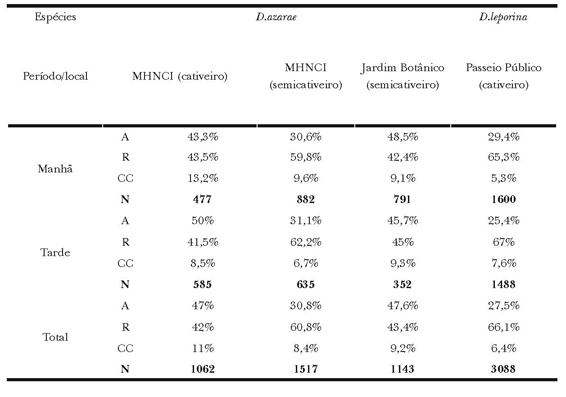
As exibições das categorias de manutenção para D. azarae no MHNCI (Χ2(2) =97; P<0,01), demonstraram mais atos alimentares e cuidados autodirigidos para o cativeiro, e menos repouso em semicativeiro. Ao comparar as frequências entre os semicativeiros Χ2(2) =123 P<0,01), as exibições de repouso e cuidados autodirigidos foram mais repetidas no MHNCI, e a alimentação no JB. Já a comparação entre o cativeiro do MHNCI e o semicativeiro do JB (Χ2(2) =15; P<0,01), revelou que a alimentação foi mais exibida no JB e o cuidado corporal em cativeiro (Tabela 3).
Tabela 3. Frequência relativa (%) e total (N) das categorias Alimentação (A), Repouso (R) e Cuidados autodirigidos (CC) exibidos por D. azarae no Museu de História Natural do capão da Imbuia, no Jardim Botânico e por D. leporina no Passeio Público.
As exibições da exploração do ambiente entre os animais do MHNCI (Χ2(2) = 51;P< 0,01), revelaram maior frequência da locomoção para o semicativeiro e exploratório para o cativeiro. A comparação entre os semicativeiros (Χ2(2) = 57; P<0,01) apresentou maior frequência locomotora no MHNCI, e maior exploratória no JB. Ao comparar o cativeiro do MHNCI e JB (Χ2(2) =19; P<0,01), as exibições de locomoção foram maiores no JB e exploração para MHNCI (Tabela 4).
Tabela 4. Frequência relativa (%) e total (N) das categorias Locomoção (L) e Exploração (E) exibidos por D. azarae no Museu de História Natural do capão da Imbuia, no Jardim Botânico e por D. leporina no Passeio Público..
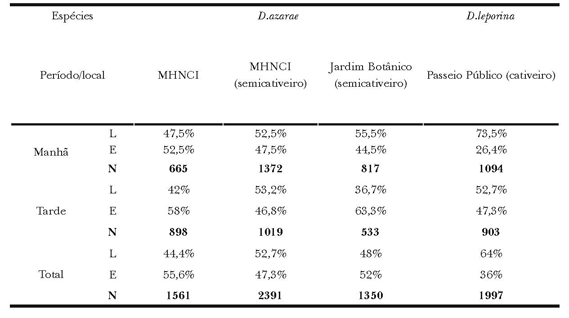
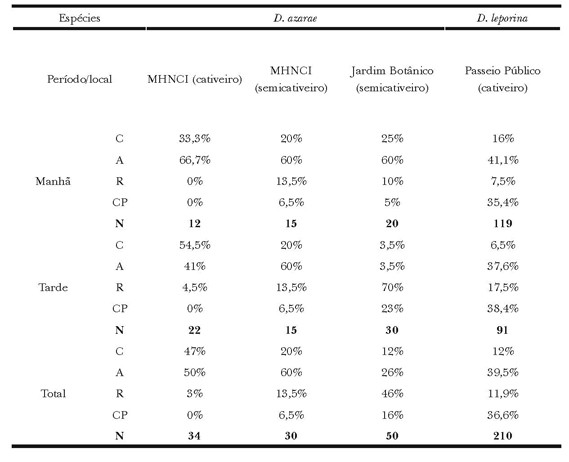
Quanto às interações sociais para D. azarae no MHNCI (Χ2(2) =17; P<0,01), as interações agonísticas, reprodutivas e cuidados parentais foram mais frequentes no semicativeiro, e as relações entre os coespecíficos, em cativeiro. A comparação entre os semicativeiros (Χ2(2) =104; P<0,01) revelou maior frequência das categorias reprodutiva e cuidados parentais no JB, e as relações entre os coespecíficos e agonísticas, maiores no MHNCI. Já a análise entre o cativeiro do MHNCI e o JB (Χ2(2) =284; P<0,01), demonstrou que os atos agonísticos e coespecíficos são mais habituais no cativeiro, e os reprodutivos e cuidados com o filhote, maiores em semicativeiro do JB (Tabela 5).
Tabela 5. Frequência relativa (%) e total (N) das categorias Coespecíficos (C), Agonístico (A), Reprodutivo (R) e Cuidados Parentais (CP) exibidos por D.azarae no Museu de História Natural do capão da Imbuia, no Jardim Botânico e por D.leporina no Passeio Público.
Período do dia
A análise entre os períodos variou apenas no JB e no PP, sendo que no JB (Χ2(2) =62; P<0,01) a maior frequência das exibições de manutenção foi verificada no horário da manhã, e explorações e interações, mais frequentes à tarde; no PP (Χ2(2) =10; P<0,01) a manutenção foi mais elevada no período da tarde e as explorações no período da manhã.
A comparação das categorias de manutenção entre os períodos revelou que a alimentação foi mais frequente pela manhã no cativeiro do MHNCI (Χ2(2) =97; P<0,01) e no PP (Χ2(2) =21,6; P<0,01). O repouso foi mais frequente pela manhã no semicativeiro do MHNCI (Χ2(2) =6,3; P<0,01), e os atos de cuidados autodirigidos mais frequentes pela manhã no MHNCI, e pela tarde no PP (Χ2(2) =21,6; P<0,01) (Tabela 3).
Já nas categorias de exploração do ambiente, a locomoção foi mais frequente nos horários matutinos para os cativeiros do MHNCI (Χ2(2) =11; P<0,01), do PP (Χ2(2) =204; P<0,01) e no JB (Χ2(2) =75; P<0,01) e a exploração no período da tarde em todos os locais (Tabela 4).
Quanto ao comportamento de interações, a exibição das relações agonísticas foi mais frequente pela manhã no JB (Χ2(2) =149; P<0,01) e PP (Χ2(2) =17; P<0,01). Os atos reprodutivos foram registrados com maior exibição no período da tarde nos mesmos locais. Os demais valores não apresentaram diferenças significativas (Tabela 5).
Comparação entre as espécies
A comparação entre as espécies (Χ2(2) =571; P<0,01), demonstrou que a manutenção e as interações sociais foram mais frequentes em D. leporina, e a exploração maior em D. azarae. Com relação à manutenção (Χ2(2) = 731; P<0,01), a alimentação e o repouso foram mais exibidos por D. azarae e os cuidados autodirigidos, por D. leporina. Para as frequências em exploração (Χ2(2) =311; P<0,01), a locomoção ocorreu mais em D. leporina, e o exploratório em D. azarae. Quanto às interações sociais (Χ2(2) =125; P<0,01), as interações com coespecíficos e interações agonísticas se mostraram mais frequentes em D. azarae, e os reprodutivos e cuidados com o filhote, elevados em D. leporina.
Discussão
A avaliação do comportamento das cutias em cativeiro e semicativeiro evidenciam a importância do tamanho do recinto para exibição de padrões motores relacionados com a manutenção, exploração e interações sociais, fundamentais para o bem-estar do animal.
A manutenção observada nas duas espécies englobou um número elevado de atos quando comparados com literatura. Santos (2005) descreveu para D. leporina somente três padrões para alimentação, um para o repouso e dois para os cuidados autodirigidos, e, ainda, não os diferenciou em categorias. Para D. ruatanica foram registrados apenas dois atos relacionados à alimentação, dois para repouso e um para limpeza corporal, sendo que a exploração foi incorporada junto com a nutrição, denominada de forrageio (Lee et al., 2000). Smythe (1978), em seu estudo com D. punctata, descreveu o forrageio em cinco atos (um para repouso e quatro para cuidados), denominados auto-grooming. O presente estudo possibilita uma visão mais ampla do conhecimento da variação de movimentos exibidos por D. azarae e D. leporina, o que é extremamente importante para o diagnóstico das condições do cativeiro, principalmente em se tratando do comportamento de manutenção, o qual está diretamente relacionado com o bem-estar animal.
O comportamento de exploração envolve a investigação do meio em que o animal está inserido, possivelmente à procura de recursos alimentares, abrigos ou parceiros sexuais. Os resultados do presente estudo corroboram os dados de Smythe (1978), que descreve os atos de enterrar alimentos e ressalta a importância do farejar no processo alimentar. As informações sobre o desenvolvimento destes atos demonstram sua importância na interação e adaptação do animal com o ambiente na sua maneira de viver e obter recursos. No entanto, o registro de apenas 11 atos foi bem menor do que o registrado por Monticelli (2005) em interações intra e interespecífica entre préas e cobaias (N=34). Segundo o autor, a exploração foi mais frequente em preás, enquanto que as cobaias exibiram mais o contato e o social, o que possivelmente está relacionado com o processo de domesticação.
As interações sociais de D. azarae e D. leporina foram caracterizadas por menos atos comportamentais (N= 20) quando comparadas com Smythe (1978) (N=25 atos), e mais quando comparado com Santos (2005) (N= 4). Deve-se considerar, porém, que enquanto o primeiro autor realizou seus estudos em campo, o segundo foi em semicativeiro. A diferenciação em categorias foi melhor observada por Smythe (1978), que verificou que as relações agonísticas de D. punctata, como a eruniração (comportamento de urinar em outro indivíduo), os relacionados à luta (agressão com unha na região posterior do oponente, mordidas e combate) e os cuidados parentais (grooming perianal e o auto-grooming do filhote) não foram observados para D. azarae e D.leporina. Dubost (1988) descreveu para D. leporina o play-dead, um ato possivelmente relacionado à submissão, no qual o animal pode ficar por 30 minutos ou mais em estado de semicatalepsia. Deve-se considerar que mesmo grupos taxonômicos distintos como cobaias e preás podem exibir atos comportamentais semelhantes durante as interações sociais (N=19) (Furnari, 2006). A exibição de atos comportamentais relacionados com interações sociais está diretamente relacionada com a presença de diferentes animais, principalmente interações com a prole. Em estudos com D. punctata livres, Clark e Galef (1977) verificaram que os padrões de interação social podem variar tanto em relação à sazonalidade reprodutiva, quanto devido à disponibilidade do recurso alimentar. Enquanto Galef e Clark (1976) registraram que a interação da mãe com o filhote nas primeiras semanas de vida determina a sua área de forrageamento. Logo, deve-se considerar que o tamanho da população e do local são requisitos importantes. Assim, é esperado que animais castrados, como os mantidos no cativeiro do MHNCI, apresentem menos interações.
A ocorrência de atos comportamentais exclusivos (D. azarae: N =3 e D. leporina: N = 6) sugere que apesar das cutias estarem confinadas em ambientes semelhantes, os mesmos podem desencadear respostas específicas adquiridas ao longo da evolução em ambientes distintos (D. leporina: Floresta Amazônica e D. azarae: Floresta com Araucária). Sabatini e Costa (2001), ao estudarem paca em sistema de cativeiro, relatam que mesmo nessas condições alguns comportamentos dependem do tipo e da forma como os recursos são oferecidos. Portanto, o ajuste comportamental é importante para a manutenção e desenvolvimento de populações silvestres mesmo em sistema de confinamento, bem como para a preservação e a exploração racional de cutias (Lopes et al., 2004).
O local de confinamento parece ter mais influência no comportamento do que a condição de manutenção, pois os recursos parecem ser semelhantes. Venturieri e Le Pendu (2006), em estudo com caititus (Tayassu tajacu; Tayassuidae) em confinamento, verificaram que áreas de recintos muito reduzidas podem levar os animais a exibirem maior estereotipa nos seus atos, bem como a exibição de movimentos repetitivos e comportamentos anormais. A possível diferença entre os locais se relaciona às condições sob as quais os animais vivem, e o tamanho dos locais pode ser a principal situação que influencia a diferenciação nas frequências dos comportamentos.
O predomínio de exibição de atos relacionados com as categorias de alimentação, interações entre coespecíficos e agonísticos parece indicar uma relação com o espaço restrito e insuficiente para a população, uma vez que a proximidade forçada entre os indivíduos nas relações alimentares pode elevar o número de encontros agonísticos. Segundo Clark e Galef (1977), as cutias podem ser solitárias ou viver em pares; ser territoriais, estabelecer hierarquias ou ser indiferentes ao coespecífico, dependendo das variáveis ambientais, sendo a presença qualitativa e quantitativa de alimentos o fator que aumenta a frequência de interações agressivas. Essas condições também foram abordadas por Santos et al. (2005), em estudo com capivaras em semiconfinamento, segundo o qual o espaço restrito e a disponibilidade de alimentos poderia levar às disputas. Essas interações aproximadas entre indivíduos também podem levar a uma maior exploração em cativeiro, na realização de estocagem alimentar, pois o número de animais neste sistema de abrigo é mais elevado do que o imposto pelas condições sociais em ambientes naturais. Para cutias em áreas de alta disponibilidade alimentar, a densidade é de 4,2 animais por hectare, ou seja, uma área de vida de 0,24 hectare por animal (Santos, 2005). Segundo Silvius e Fragoso (2003), a área de vida de D. leporina na Amazônia varia de 3 a 8 ha, embora as áreas preferidas estejam relacionadas com disponibilidade de alimento, havendo maior forrageamento e armazenamento em locais com maior abundância das sementes preferidas. Deve-se considerar que os animais avaliados no presente estudo em cativeiro possuem uma área de vida extremamente reduzida, uma vez que no Passeio Público a área é de aproximadamente 0,0013 hectare por animal.
As atividades exploratórias em semicativeiro sugerem uma busca elevada pelo alimento e sua estocagem. O estudo com D. leporina Santos (2005), evidencia que as cutias gastam parte do seu tempo forrageando, correspondendo a frequências de 36,9% dos comportamentos exibidos no Bosque de Jequitibás (SP), contra uma média de 50,3% para os semicativeiros estudados. Já um estudo realizado com D. ruatanica em ambiente natural, porém antropizado, no Panamá, demonstra frequências para farejar de 12,1%, alimentar de 21,9%, e cavar para procura e estocagem em 3,3%, totalizando 37,3% (Lee et al., 2000), sendo os mesmo atos realizados em média, nos semicativeiros, para farejar e explorar, agrupados em 49,6% e alimentar 39%. Portanto, as frequências exploratórias possivelmente relacionam-se à abundância de itens alimentares, sendo amostradas com maiores frequências com relação aos trabalhos citados, o que pode sugerir uma procura por recursos alimentares melhores.
As interações agonísticas no sistema de semicativeiro no MHNCI, podem indicar a disputa por recursos essenciais e limitantes, como água e alimento. As cutias, sendo animais frugívoros territorialistas, podem precisar defender seu território o ano todo, na defesa de seus recursos (Santos, 2005), ou variar conforme a disponibilidade sazonal do mesmo (Clark & Galef, 1977). Porém, esta defesa agonística por obtenção de alimentos pode se refletir em altas frequências de cuidados parentais, verificadas no JB, pois como descrito por Smythe (1978), estes padrões são frequentes enquanto há uma disponibilidade alimentar para as cutias, uma vez que, quanto maior a disponibilidade alimentar, mais energia há para os cuidados com a prole, o que diminui muito a possibilidade de abandono e morte (Smythe, 1978). Segundo Galef e Clark (1976), 50% da atividade diária da mãe é direcionada ao filhote, sendo registrado a fêmea de D. punctata agredindo o filhote encontrado fora do sitio de forrageamento. Portanto, as disputas provavelmente são por recursos melhores, o que exprime êxito na manutenção dessas populações. Deve-se considerar que embora o estresse fisiológico de D. punctata possa ser inferido através do estudo comportamental (Korz, Hendrichs & Militzer, 2004), fêmeas apresentam reações agonísticas especificas para diferentes contextos (Korz, 1991). Para cobaias (Cavia porcellus) Takamatsu, Tokumaru e Ades (2003) também verificaram agressão do adulto contra o filhote, como o registrado para D. leporina e D. punctata, porém, as mesmas foram mais frequentes quando o adulto não era o genitor.
A diferenciação nas frequências de atos entre ambientes de cativeiro e semicativeiro obtidas no presente estudo são equivalentes àquelas obtidas por Santos (2005) para D. leporina em que a frequência de farejar foi 36,7%, contra 36% no cativeiro do PP; de alimentação de 27,9% para o semicativeiro, contra 27,5% em confinamento e o repouso de 15% contra 66,1% para o cativeiro. Estes valores indicam que em confinamento os animais mudam seu padrão de exibição comportamental (Campos, Santos & Stez, 2006; Venturieri & Le Pendu, 2006), sendo as frequências de inatividade elevadas para o cativeiro. Portanto, em confinamento, os animais alteram seus ritmos de atividade, aumentando ou mesmo diminuindo-o, dependendo das condições impostas pelo manejo. Nessas situações é recomendada a aplicação de técnicas de enriquecimento do ambiente, que consiste na criação ou simulação de situações mais naturais, pela introdução de variedades na estimulação física (substrato), sensorial (sons de vocalizações e ervas aromáticas) e social (interação intra ou interespecífica).
No período da manhã, as atividades de locomoção demonstraram maior frequência, o que possivelmente se relaciona pela busca alimentar. Esta atividade, no mesmo período, é observada em cotiaras, Myoprocta pratti (Dasyproctidae), nas primeiras horas do amanhecer, porém diferente do descrito para cutias, nas quais a locomoção é mais acentuada no período da tarde (Dubost, 1988). Sendo esta atividade visualizada em maiores frequências também nos dois cativeiros, é possível que os animais mudem seus ritmos de acordo com as condições de manutenção, pois podem estar relacionadas com o horário de ceva (alimentação). A procura alimentar pode elevar o comportamento agonístico pela disputa entre opositores, também mais frequentes neste período. Um manejo no ambiente introduzindo maior número de locais de alimentação poderá diminuir as disputas, porém estimulando a locomoção de maneira adequada, sem comportamentos estereotipados. No período da tarde, as atividades exploratórias sugerem que a procura por alimentos estocados pela manhã seja mais frequente. O estudo de Dubost (1988) relata que as cutias podem ter uma atividade locomotora pela procura alimentar acentuada no horário da tarde, o que pode indicar uma nova fase de alimentação ao entardecer. Nos semicativeiros estudados, este ritmo pode estar relacionado com o horário de ceva realizada pela manhã, na qual a quantidade de alimentos é elevada, induzindo os animais à estocagem.
As análises de frequências entre as espécies demonstram que apesar de estarem expostas às mesmas condições de confinamento, apresentam padrões próprios de cada espécie. As frequências de manutenção e interações sociais para D. leporina indicam a influência do tamanho do recinto e o número de animais mantidos, bem como as explorações para D. azarae, que provavelmente podem estar relacionados também com a proximidade entre os recintos. As frequências de repouso para D. leporina indicam que a distância entre os indivíduos pode ter sido reduzida, porém adaptada pelos animais. Já a alimentação e os cuidados autodirigidos para D. azarae, podem relacionar-se com a disponibilidade alimentar e a falta de estimulação para outra atividade. Na natureza os animais passam a maior parte do tempo à procura de alimentos e outros recursos e interagindo com outros animais, evitando predadores (Bosso, 2007). Portanto, em cativeiro os animais se adaptam a realizar atividades diferentes do esperado em seu ambiente natural. A locomoção acentuada em D. leporina sugere a importância do tamanho do recinto nas frequências comportamentais. Os atos exploratórios mais frequentes para D. azarae podem estar relacionados com a estocagem alimentar, devido à densidade de animais presentes no recinto. Isto também se reflete nas interações entre coespecíficos e agonísticos, pois a limitação e a concentração de recursos (alimento e abrigo) em poucos e pequenos locais, podem acentuar a competitividade entre os indivíduos (Santos et al., 2005). Portanto, altas frequências alimentares e exploratórias em cativeiro podem indicar a restrição estrutural do alojamento. As exibições de interações reprodutivas em D. leporina provavelmente relacionam-se com o número de fêmeas no PP. Apesar de serem animais monogâmicos, em períodos de alta disponibilidade de recursos em ambientes naturais (Smythe, 1978), seu comportamento em cativeiro pode se modificar devido os recursos serem constantes. É possível, portanto, verificar que as espécies têm semelhança comportamental, com uma extensa gama de atos que podem ser exibidos de acordo com as condições de manutenção. Portanto, o plano de manejo para tais espécies pode ter as mesmas características.
Os dados do presente estudo evidenciam que apesar dos padrões motores utilizados por ambas as espécies, nas duas condições de confinamento, e dos três ambientes serem muito semelhantes, há uma diferenciação quanto à frequência de exibição dos mesmos. O predomínio de exibição de atos relacionados com as categorias de alimentação, interações entre coespecíficos e agonísticos parece indicar uma relação com o espaço restrito e insuficiente para a população. Apesar das duas espécies estarem expostas às mesmas condições de confinamento, apresentam seus próprios padrões. Esses resultados sugerem que as condições de manutenção são extremamente importantes e que a quantificação dos comportamentos permite conhecer o quanto o animal está tendo condições de exibir o seu repertório comportamental normal. O manejo adequado de animais cativos deve ter como preocupação principal a elaboração de situações para estimulação dos comportamentos naturais, através do enriquecimento ambiental, que deve proporcionar aos animais circunstâncias de bem-estar, como a criação ou simulação de situações mais naturais, pela introdução de variedades na estimulação física (substrato), sensorial (sons de vocalizações e ervas aromáticas) e social (interação intra ou interespecífica). Assim, sugere-se que para a manutenção de cutias em cativeiro sejam consideradas questões como densidade populacional, proporção de fêmeas, machos e jovens, frequência, horário e forma de oferecimento dos alimentos, relacionar o horário e frequência das atividades dos funcionários como manutenção e o comportamento do animal, proporcionar condições naturais para exibição de comportamentos exploratórios, como presença de troncos caídos para desgaste dos dentes, substratos adequados para estocagem de alimento, maior quantidade e diversidade de abrigos e pontos de fuga e, por fim, levar em consideração os itens alimentares utilizados no seu local de origem como raízes, sementes, flores e frutos. No semicativeiro, deve-se considerar as mesmas questões citadas acima, ressaltando, ainda, a manutenção de um sub-bosque o mais próximo do original e com densidade e diversidade suficientes para manter os recursos de alimentação e abrigo para os animais.
Referências
Almeida, F. M. (2004). Bioética Aplicada aos animais de zoológicos. Fundação RIOZOO. Recuperado em 29 de agost. 2007, da Academia Brasileira de Meio Ambiente: http://www.abma.com.br/2004/notes/208.pdf [ Links ]
Bosso, P. L. (2007). Zoológico de São Paulo. Recuperado em 12 mar 2007, do Zoológico de São Paulo : http://www.zoologico.sp.gov.br/peca7.htm [ Links ]
Broom, D. M, & Molento, C.F.M. (2004). Bem-Estar Animal: Conceito e Questões Relacionadas. Archives of Veterinary Science, 9 (2),1-11. [ Links ]
Campos J. T., Santos, E.F., & Setz, E. Z. F. (2006). Padrão de atividades e o enriquecimento ambiental nos quatis Nasua nasua em cativeiro. [Resumo]. XVI Congresso Interno de Iniciação Científica Unicamp. Recuperado em 23 de out. 2007, da PRP (Pró-reitoria de Pesquisa e Graduação da Unicamp): http://www.prp.unicamp.br. (texto completo). [ Links ]
Clark, M. M, & Galef, B. G. (1977). Patterns of agonistic interaction and space utilization by agoutis (Dasyprocta-punctata). Behavioral Biology, 20 (1), 135-140. [ Links ]
Dubost, G. (1988). Ecology and social life of the red achouchy, Myoprocta exilis; Comparison with the orange-rumped agouti, Dasyprocta leporina. Journal of Zoology of London, 214, 107-123. [ Links ]
Eisenberg, J. F., Redford, K. H., & colaboradores. (Eds). (1992). Mammals of the Neotropics – The Central Neotropics Ecuador, Peru, Bolivia, Brazil. (Vol. 3). Chicago and London: The University of Chicago Press.
Furnari, N. (2006). Corte intra e interespecífica em cobaias (Cavia porcellus) e preás (Cavia aperea). Dissertação de mestrado). Universidade de São Paulo. SP. [ Links ]
Galef, B. G., & Clark, M. M. (1976). Non-nurturent functions of mother-young interaction in agouti (Dasyprocta-punctata). Behavioral Biology, 17(2), 255-262. [ Links ]
Hashimoto, C. Y. (2008). Comportamento em cativeiro e teste da eficácia de técnicas de enriquecimento ambiental (Físico e Alimentar) para jaguatiricas (Leopardus pardalis). Dissertação de mestrado). Universidade de São Paulo. SP. [ Links ]
Korz, V. (1991). Social-relations and individual coping reactions in a captive group of central-american agoutis (Dasyprocta punctata). Zeitschrift fur Saugetierkunde-International Journal of Mammalian Biology, 56 (4), 207-218. [ Links ]
Lange, R. R. (1998). Criação e Relocação de Cutias Dasyprocta azarae Lichtenstein, 1823 (Dasyproctidae, Mammalia) em Área Verde Urbana, Curitiba-PR. Dissertação de mestrado. Universidade Federal do Paraná, Curitiba, PR. [ Links ]
Lee, T. E., Rhodes, K. R., Lyons, J. L., & Brannan, D. K. (2000).The natural history of the Roatan Island agouti (Dasyprocta ruatanica), a study of behavior, diet and description of the habitat. The Texas Jornal of Science, 52, 159–164. Recuperado em 7 de jan. 2008, da http://www.accessmylibrary.com
Lopes, J. B., Cavalcante, R. R., Almeida, M. M., Martins, M. A., Moura, S. G., Dantas Filho, L. A., & Conceição, W. L. F. (2004). Desempenho de Cutias (Dasyprocta prymnolopha) criadas em cativeiro do nascimento até o desmame em Teresina, Piauí. Revista Brasileira de Zootecnia, 33, 2318-2322. [ Links ]
Monticelli, P. F. (2005). Comportamento e comunicação acústica em cobaias e preás. Tese de doutorado, Universidade de São Paulo, São Paulo. [ Links ]
Nowak, R. M. (Ed.). (1999). Walker's mammals of the World. (6a ed.). Order Rodentia (Vol. 2). Baltimore and London: The Johns Hopkins University Press. [ Links ]
Prefeitura Municipal de Curitiba. (2007). Parques e Bosques. Jardim Botânico <http://www.curitiba.pr.gov.br/conteudo/jardim-botanico-secretaria-municipal-do-meio-ambiente/287>. Acesso 10 de junho de 2011. [ Links ]
Reis, N. R., Peracchi, A. L., Pedro, W. A., & Lima, I. P. (2006). Mamíferos do Brasil. Em Oliveria, J. A. & Bonvicino, C. R., Sobre Ordem Rodentia (pp. 347-399). Londrina, PR. [ Links ]
Roll, V. F. B., Rech, C. L. S., Xavier, E. G., Rech, J. L., Rutz, F., & Del Pino, F. A. B. (2006). Comportamento Animal: Conceitos e Técnicas de Estudo. Pelotas: UFPEL. [ Links ]
Sabatini, V., & Costa, M. J. R. P. (2001). Etograma da paca (Agouti paca, Linnaeus, 1766) em Cativeiro. Revista de Etologia, 1(3), 3-12. [ Links ]
Santos, C. R., Filho, H. O., Barbosa, O. R., Cardozo, R. M., Araujo, D. N., Fornari, D. C., Guaragni, M., & Fregadolli, R.M. (2005). Etologia de capivaras (Hydrochaeris hydrochaeris L. 1766) jovens semiconfinadas no Norte do Estado do Paraná. Acta Scientiarum – Animal Scienses. 1(27), 163-169. Recuperado em 10 de jun. 2007, dos Periódicos da UEM (Universidade Estadual de Maringá): http://www.periodicos.uem.br
Santos, E. F. (2005). Ecologia da cutia Dasyprocta leporina (Linnaeus, 1758) em um fragmento florestal urbano em Campinas-SP (Rodentia: Dasyproctidae). Tese de doutorado, Universidade Estadual Paulista, Rio Claro, SP. [ Links ]
Silvius, K. M., & Fragoso, J. M. (2003). Red-rumped Agouti (Dasyprocta leporina) home range use in na Amazonian Forest: implications for the aggregation distribuition of Forest trees. Biotropica, 35, 74-83. [ Links ]
Smythe, N. (1978). The natural history of the Central American agouti (Dasyprocta punctata). Tese of Smithsonian Tropical Research Institue. Smithsonian Contributions to Zoology. [ Links ]
Sorensen, T. (1948). A method of establishing groups of equal amplitude in plant sociology based on similarity in species content and its application to analyses of the vegetation on danish commons. Biologiske. (Kongelige Danske Videnskabernes Selskabs), 5, 1-34. [ Links ]
Takamatsu, A. T., R., Tokumaru, S., & Ades, C. (2003). Allosuckling in guinea pigs (Cavia porcellus), Revista de Etologia, 5, 203. [ Links ]
Venturieri, B., & Le Pendu, Y. (2006). Padrões de Atividades de caititus (Tayassu tajacu) em cativeiro. Revista de Etologia, 1(8), 35-43. [ Links ]
Recebido em: 28/09/2009
Aceito em: 19/07/2010
 Endereço para correspondência
Endereço para correspondência
Silvane Krul Kaiser / Marta Luciane Fischer
Núcleo de Estudos do Comportamento animal
Centro de Ciências Biológicas e da Saúde
Escola de Saúde e Biociências
Pontifícia Universidade Católica do Paraná
Rua Imaculada Conceição, 1155. Prado Velho CEP 80215-901
E-mail: marta.fischer@pucpr.br; silvanekkaiser@hotmail.com
Teresa Cristina Castellano Margarido
Departamento De Zoológico
Secretaria Municipal do Meio Ambiente
Prefeitura Municipal de Curitiba
Av. Presidente Faria, s/n. Centro CEP 80020-290
