










Serviços Personalizados
Journal

artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkUniversitas Psychologica
versão impressa ISSN 1657-9267
Univ. Psychol. v.7 n.2 Bogotá ago. 2008
Efecto de agonistas y antagonistas de la dopamina en ambientes de reforzamiento variables*
Effects of Dopamine Agonists and Antagonists in Variable Reinforcing Environments
Jorge Arturo Balderrama-Trápaga**, I; Carlos Fernando Aparicio-Naranjo***, II
IUniversidad Veracruzana, México
IIUniversidad de Guadalajara, México
ABSTRACT
The anhedonia hypothesis maintains that pleasure for food-reward is determined by dopamine activity in the brain. The cumulative body of empirical evidence shows that dopamine agonists and antagonists (e.g., Methylphenidate and Haloperidol, respectively) decrease operant behavior maintained by positive reinforcement. The present study used the generalized matching law (Baum, 1974) to assess effects of these drugs on the motor system and the organism’s motivation for food reinforcers. Thirty-two rats searched for food in a choice situation where the reinforcer ratio provided by two alternatives changed everyday. The results confirmed that the drugs do not impede the rats’ rapid adaptation to dynamic changes in the contingencies of reinforcement. Preference favored with more responses the alternative associated to the higher probability of reinforcement, showing an increment in sensitivity of behavior ratio to changes in food ratios. The alternative demanding the higher motor requirement showed a decrement in the number of responses, suggesting effects of the drugs in the organism’s motor system.
Key words authors Methylphenidate, Haloperidol, Choice, Motivation, Reinforcement, Rats.
Key words plus Laboratory research, Dopamine, Dopamine agonists, Dopamine antagonists.
RESUMEN
Según la hipótesis de la anhedonia el valor placentero de la comida está determinado por la actividad de dopamina en el cerebro. La evidencia empírica muestra que los agonistas y antagonistas a la dopamina (e.g., metilfenidato y haloperidol, respectivamente) disminuyen la conducta operante mantenida con reforzamiento positivo. Se utilizó la ley generalizada de igualación (Baum, 1974) para evaluar efectos de estas drogas en la motricidad y motivación del organismo por la comida. Treinta y dos ratas buscaron alimento en una situación de elección donde la razón de reforzamiento que proporcionaban dos alternativas cambia diariamente. Los resultados confirmaron que las drogas no impiden la rápida adaptación de las ratas a los cambios dinámicos en las contingencias de reforzamiento. Se evidenció un incremento en la sensibilidad de las razones de respuestas a los cambios en las razones de los reforzadores. La alternativa de mayor demanda motriz para el organismo mostró una reducción en el número de respuestas, sugiriendo efectos de las drogas en la motricidad del organismo.
Palabras clave autores Metilfenidato, Haloperidol, Elección, Motivación, Reforzamiento, Ratas.
Palabras clave descriptores Investigación de laboratorio, Dopamina, Agonistas de dopamina, Antagonistas de dopamina.
Introducción
La investigación en farmacología conductual sugiere que la dopamina juega un papel determinante como mediador o regulador del valor reforzante que tienen los estímulos primarios como la comida, el agua, el sexo y las drogas de abuso (e.g., Wise, Spindler, deWit & Gerber, 1978). Muchos trabajos reportan que las drogas que bloquean la actividad de dopamina (los neurolépticos) ocasionan decrementos en la conducta operante mantenida por reforzamiento positivo (Liebman & Butcher, 1973; Ettenberg, 1989; Franklin & McCoy, 1979; Fouriezos & Wise, 1976; Fouriezos, Hansson & Wise, 1978; Gallistel, Boytin, Gomita & Klebanoff, 1982; Miller, Wickens & Beninger, 1990; Wise, Spindler, deWill & Gerber, 1978; Wise, 1982; Wise & Colle, 1984; Wise & Raptis, 1986). También se ha mostrado que los neurolépticos interfieren con la conducta de dirigirse al alimento y consumirlo (Aparicio, 1999, 2001; Aparicio & Velasco, 2003; Berridge & Robinson, 1998).
En conjunto, esas investigaciones sugieren que una reducción en la actividad de dopamina ocasiona una disminución en el valor reforzante del estímulo positivo (es decir la comida), como resultado de esto la conducta mantenida por ese reforzador alcanza la extinción. De acuerdo con esta idea, un incremento en la actividad de dopamina debería ocasionar los efectos opuestos, un aumento en el valor reforzante de la comida y un incremento en la frecuencia de la conducta. Contradiciendo esta idea, existen datos que muestran que las dos drogas, agonistas y antagonistas a la dopamina, reducen la tasa de respuestas mantenida por reforzamiento positivo (Heyman, 1983; Wise & Bozarth, 1987). A la fecha no se ha identificado claramente los mecanismos responsables de este efecto, ni tampoco se ha aclarado como actúa la dopamina para regular el valor reforzante de los estímulos positivos (Berridge & Robinson, 1998). Sabemos que en los receptores D1, D2 y D3 la actividad de dopamina incrementa con la administración de drogas agonistas a ésta como la anfetamina; pero los antagonistas a la dopamina como el pimozide casan una disminución en la disponibilidad de dopamina en esos mismos receptores (Heyman, 1983; Wise & Bozarth, 1987).
Otros autores sugieren que los neurolépticos reducen la emisión de conductas operantes porque inducen en el organismo un déficit motriz (Tombaugh, Tombaugh & Anisman, 1979; Wise & Bozarth, 1987). Consistente con esta idea, se dice que la tasa de respuestas incrementa con la administración de la meta-anfetamina porque esta droga causa un aumento en la concentración de receptores dopaminérgicos presinápticos, lo cual produce una elevación en la actividad motriz. Diversos trabajos han mostrado que el incremento en la tasa de respuestas que se observa bajo el efecto de la anfetamina depende de la tasa de respuestas en la línea base, del programa de reforzamiento que se haya utilizado para mantenerla y de la especie que se estudie. Específicamente, la anfetamina puede incrementar la tasa de respuestas bajo programas que usualmente producen un nivel bajo de respuestas, como los programas de reforzamiento diferencial de tasas bajas y los de intervalo variable (Clark & Steele, 1966; Lucki & DeLong, 1983). Los efectos de la anfetamina en la conducta operante también dependen de los requisitos impuestos por las contingencias de reforzamiento (Dews, 1958; Dews & Morse, 1961; Clark & Steele, 1966; Kelleher & Morse; 1968; McMillan, 1968). Por ejemplo, una misma dosis de meta-anfetamina que no afecta la tasa de respuestas en un programa de reforzamiento diferencial de tasas bajas, cuando se administra en un programa de razón fija (Heffner, Drawbaugh & Zigmond, 1974; Owen, 1960) o en uno de intervalo variable (Bradshaw, Ruddle & Szabadi, 1981; Lucki, 1983; Lucki & DeLong, 1983) produce un incremento en la tasa de respuestas.
Además, se ha mostrado que los efectos de la anfetamina dependen tanto de la dosis administrada, como de la tasa de respuestas previa (es decir, la línea base) a su administración (Clark & Steele, 1966). Para explorar esta posibilidad, Heyman (1983) entrenó ratas para responder en una palanca y producir comida de acuerdo con programas múltiples de reforzamiento positivo. Después de obtener una línea base estable de esa respuesta, administró a las ratas intraperitonealmente varias dosis de pimozide y anfetamina. Los efectos de esas drogas en el sistema motriz y la motivación de por la comida, Heyman los estimó con los parámetros (k y r0, respectivamente) de la relación hiperbólica de Herrnstein (1974). Sus resultados mostraron que las dos drogas en dosis intermedias causaron un incremento en la tasa de la respuesta de presionar la palanca, pero en dosis altas las dos drogas causaron lo opuesto, un decremento en la tasa de respuestas, pero no su extinción. Los resultados de Heyman no apoyaron la hipótesis de la anhedonia, ninguna de las dos drogas eliminó el interés de las ratas por la comida (Wise, 1985) ni tampoco su valor reforzante (Cousins, Wei & Salamone, 1994). Esto es importante porque Heyman esperaba un incremento en la tasa de respuestas con la dosis más alta de anfetamina, pero no el efecto opuesto. Su explicación fue que un déficit motor causado por el incremento en la actividad de la dopamina, interfirió con la conducta de presionar la palanca causando su decremento (Heyman, 1983).
El presente estudio exploró esa posibilidad con ratas que respondieron en dos palancas para obtener el alimento que éstas proporcionaban en una situación de elección dinámica. Para determinar si el valor reforzante del alimento cambia con una alteración en la actividad de dopamina, a las ratas se les administró vía intraperitoneal una droga agonista y otra antagonista a la dopamina (metilfenidato y haloperido, respectivamente).
Las estimaciones de los efectos de esas drogas en el sistema motor y la motivación de las ratas por el alimento, se hicieron con la ley de igualación generalizada (Baum, 1974)
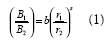
que expresada en logaritmos toma la siguiente forma,
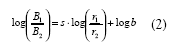
donde B1 and B2 son asignaciones de conducta (medidas en tiempo o respuestas) a las Alternativas 1 y 2; r1 y r2 son tasas de alimento obtenidas de las Alternativas 1 y 2; b es una medida del sesgo hacia una de las dos alternativas que surge de factores diferentes a r1 y r2; y, s es la sensibilidad de la razón de conducta a los cambios en la razón de tasas de alimento. Un valor de s igual a 1 corresponde a un empate o igualación estricta, un valor de s mayor a 1.0 representa una sobreigualación donde los cambios en la razón de respuestas son más extremos que los esperados en la igualación estricta y un valor de s menor a 1.0, representa la subigualación donde los cambios en la razón de respuestas son menos extremos que los esperados en la igualación estricta. En estudios conducidos en nuestro laboratorio (Aparicio, 1999, 2001; Aparicio, Velasco & Balderrama, 2004; Aparicio & Velaso, 2003), los efectos del haloperidol, un antagonista a la dopamina, en el sistema motor y la motivación de las ratas por la comida, se han evaluado analizando cambios en los valores de los parámetros b y s, respectivamente.
Experimento 1
El Experimento 1 comparó los efectos del haloperidol con los del metilfenidato en la conducta motora y la motivación de las ratas por la comida. Implementó un medio ambiente de reforzamiento semiestable para evaluar si éste es un factor que determina el efecto de estas drogas en la conducta de elección.
Método
Sujetos
Dieciséis ratas albinas macho cepa Wistar, de 90 días de edad aproximadamente, fueron alojadas en habitáculos independientes y mantenidas en un régimen de privación de alimento que controló el peso corporal al 85% del nivel mostrado en alimentación libre. Durante el experimento las ratas tuvieron acceso libre al agua y permanecieron en una habitación que alternó ciclos de iluminación/ oscuridad de 12 horas y mantuvo una temperatura ambiente aproximada entre los 18 y 22 ° C.
Aparatos
Se utilizaron cuatro cajas operantes modulares para ratas (Coulbourm E10-18TC), de 31 cm de largo por 26 cm de ancho y 32 cm de altura, con paredes anteriores y posteriores de aluminio y paredes laterales de acrílico transparente. En el centro de la pared anterior de caja a 2 cm del piso y a 20 cm del techo de la caja, se montó un comedero (E14-01) de 3cm de ancho y 4 cm de largo. Dos palancas no retráctiles (Coulbourn E23-17) que requerirán una fuerza de 0.2 N para ser operadas, se instalaron en la misma pared a 10 cm del piso, 13 cm del techo de la caja y a 2.5 cm de cada una de las paredes laterales izquierda y derecha. Se colocaron dos focos de luz blanca (E11-03) de 24 V DC: uno, 2 cm arriba de la palanca derecha y otro 2 cm arriba de la palanca izquierda. Un dispensador de alimento (E14-24) arrojó pellas de 45 mg (P. J. Noyes Resarch Diets, NY) en el comedero. Una bocina (E12-01) de 2.6 cm de ancho por 4 cm de alto, montada en la pared anterior y alineada al centro a 2 cm del techo, permitió la presentación de un ruido blanco constante. Una palanca no retráctil (Coulbourn E21-03) que requerirá de una fuerza de 0.2 N para ser operada, se montó centrada en la pared posterior a 10 cm del piso y a 13 cm del techo de la caja. Una rejilla de malla metálica (E10-18NS) de 27 cm de largo por 28.5 cm de ancho formó el piso de la caja. Cada caja operante se introdujo dentro de un cajón aislante a prueba de ruidos de 78 cm largo por 54 cm de ancho y 51cm de altura. Una interfase (L18-16S/C) instalada en una computadora de escritorio (Compaq) se conectó en serie con una caja de distribución (L18-16XVC) y un controlador de eventos (Habitest EZ Linc L9 102S). Un paquete de software Grafic State 1.0 (Coulbourn instrument) sirvió para programar y recolectar los eventos de estímulo y respuesta con una aproximación a la décima de segundo.
Agentes farmacológicos
Dos dosis de Haloperidol de 0.05 y 0.10 mg/kg se obtuvieron mezclando porciones de haloperidol y de ácido tartárico respectivamente en una solución salina fisiológica 60 ml. Las mezclas resultantes se disolvieron en vapor de agua a una temperatura promedio de 40° C. Cada dosis se administró por separado en un volumen de 1 ml/kg por vía intraperitoneal (ip) 30 minutos antes de iniciar la sesión (tiempo que en nuestra experiencia es suficiente para que la droga actúe a su nivel máximo durante una sesión de 60 minutos). Las dosis (0.05 y 0.10 mg/kg) se evaluaron en dos ocasiones en órdenes ascendente y descendente con dos días intercalados entre una y otra dosis para evitar efectos acumulativos.
El Clorhidrato de metilfenidato se disolvió en una razón de 1 comprimido por cada 1 ml de solución salina fisiológica a una temperatura ambiente, la solución resultante fue una concentración de 10 mg/ml, para garantizar su homogeneidad se sonicó la solución durante 60 minutos quedando compuestos insolubles (excipientes). Después de sonicar la solución, ésta se filtró por un papel millipore de 0.22 µm de poro para obtener dos dosis de exposición de 5 y 10 mg/kg que se administraron por separado en un volumen de 1 ml/kg por vía intraperitoneal (ip) 30 minutos antes de iniciar la sesión experimental. Las dosis (5 y 10 mg/kg) se evaluaron en órdenes ascendente y descendente con un día intercalado entre una y otra para evitar efectos acumulativos de la droga.
Entrenamiento
De acuerdo a un criterio aleatorio, las ratas se formaron en cuatro grupos de cuatro ratas cada uno y en la primera sesión recibieron entrenamiento en el comedero en presencia de la palanca izquierda. Durante esa sesión, un programa de reforzamiento continuo (RFC) reforzó con comida la conducta de presionar esa palanca hasta que las ratas obtuvieron 30 reforzadores consecutivos. Después, la palanca izquierda se retiró de la caja y se montó la palanca derecha. En otra sesión el mismo programa de RFC sirvió para reforzar las presiones en la palanca derecha hasta que las ratas obtuvieron otros 30 reforzadores consecutivos; lo cual ocasionó que la palanca derecha se retirara de la caja y se montara la palanca de cambio en la pared posterior la caja. La tercera sesión también reforzó con comida las presiones en la palanca de cambio hasta que las ratas obtuvieron 30 reforzadores de acuerdo con el mismo programa de RFC. Al siguiente día se inició el experimento propiamente dicho.
Procedimiento
El procedimiento implementó una situación de elección basada en el método de Davison y Baum (2000). Un programa concurrente con dos componentes de intervalo variable (Conc IV 11 s – IV 11 s) proporcionó comida (reforzadores) en las dos palancas frontales cada 11 segundos en promedio. Para estos fines, la computadora revisaba cada 3 segundos un circuito lógico que asignaba un reforzador en la palanca izquierda o en la derecha de acuerdo con siete pares de probabilidades: 0.27- 0.01; 0.25-0.03; 0.210-.07; 0.14-0.14; 0.07-0.21; 0.03-0.25 y 0.01-0.27 (nótese que en cada par la suma es .28 y que el producto de 3 dividido entre 0.28, es 10.7 el valor promedio de los intervalos). Así, los siete pares de probabilidades proporcionaron comida en las palancas de acuerdo a siete razones de reforzadores (27:1; 9:1; 3:1; 1:1; 1:3; 1:9 y 1:27). Cada razón se asignó por separado a las palancas en orden aleatorio y entregó 50 reforzadores por día de acuerdo a un método de elección forzada (Stubbs & Pliskoff, 1969); esto significa que cuando se asignaba un reforzador a una palanca, no se preparaba otro reforzador en la misma o en la otra palanca hasta que la rata obtenía el que ya estaba asignado. En todo momento de la sesión la rata podía abandonar la palanca en la que estaba respondiendo y trasladarse a la otra palanca para buscar reforzadores que no se ponían disponibles en esa palanca hasta que la rata se trasladaba a la pared posterior de la caja y presionaba la palanca de cambio que ahí se encontraba. En cinco condiciones que duraron 42 sesiones cada una, la palanca de cambio requirió 1, 4, 8, 16 y 32 respuestas para permitir la disponibilidad del reforzador en una palanca después de que la rata abandonó de la otra palanca.
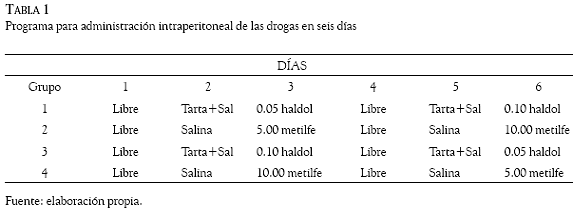
En las 42 sesiones de cada una de las 5 condiciones, cada razón de reforzadores permaneció 6 días vigente en las palancas durante los cuales se implementó el programa de administración de droga como sigue: 1) en los días 1 y 4 a las ratas no se les inyectó solución alguna; 2) los días 2 y 5 fueron de vehículo en los que a las ratas se le inyectó intraperitonealmente (ip) 1 ml/kg de una de dos soluciones, ácido tartárico disuelto en agua salina (control de haloperidol), o sólo se les inyectó agua salina (control de metilfenidato) 30 minutos antes de que iniciara la sesión para estimar cualquier posible efecto que la inyección haya tenido como estímulo incondicionado sobre la ejecución concurrente de las ratas; 3) los días 3 y 6 fueron de droga en los que a las ratas se les inyectó (ip) haloperidol o metilfenidato 30 minutos antes de iniciar la sesión. La Tabla 1 muestra el programa de droga para los cuatro grupos que difieren con respecto a la droga (haloperidol o metilfenidato) administrada y a la dosis de ésta que recibieron en los días 3 y 6. El programa de droga se repitió con cada una de las siete razones de reforzador arriba señaladas, de manera que en 42 días las dos dosis de una misma droga se evaluaron con todas las razones de reforzadores y con el requisito de cambio de una respuesta. En los siguientes 42 días con el requisito de cambio de 4 respuestas y las siete razones de reforzadores, las dos dosis de una misma droga se volvieron a evaluar de acuerdo con el diseño que muestra la Tabla 1. El mismo ciclo de 42 días se repitió con los requisitos de cambio de 8, 16 y 32 respuestas. En total, 210 días de experimentación fueron necesarios para evaluar las dos drogas con las siete razones de reforzadores y los cinco requisitos de respuesta de cambio.
Análisis de los datos
Los efectos del haloperidol y el metilfenidato en la conducta de elección de las ratas, se analizaron con los parámetros (s y b) de la ley generalizada de la igualación (Baum, 1974). Para cada sujeto y en cada condición, se contó por separado el número de respuestas emitidas en cada visita en las palancas izquierda y derecha, la duración de cada visita, el tiempo de traslado de una palanca a la otra y el número veces que las ratas cambiaron de palanca. Con los cómputos del número de respuestas y duraciones de visita se obtuvieron las razones de respuestas y tiempos invertidos en las palancas, las mismas que se transformaron en logaritmos con base 10 y entraron en la Ecuación 2 como los factores de la variable dependiente. Las siete razones de reforzadores también se transformaron en logaritmos con base 10 y entraron en la misma ecuación como los factores de la variable independiente. Por medio de regresiones lineales se estimaron los valores de los parámetros de sensibilidad al reforzamiento (s) y sesgo (b) en los siete componentes (razones de reforzamiento) con cada uno de los requisitos de respuesta de cambio (RCA). El análisis permitió hacer comparaciones entre las dosis de una misma droga y entre las drogas. También se analizaron los efectos de las interacciones entre el requisito de respuesta de cambio y la droga administrada.
Resultados
Los datos de todas las sesiones se organizaron y clasificaron por sujeto, día, razón de reforzadores, requisito de cambio, droga y dosis administrada. En cada sesión se contó, reforzador por reforzador, el número de respuestas en la palanca izquierda y las emitidas en la palanca derecha. Esos conteos se usaron para computar razones de respuestas (izquierda /derecha) que se transformaron en logaritmos (base 10) y se graficaron en las figuras 1 a 4 en función del requisito de cambio (1, 4, 8, 16, ó 32 respuestas). Los números en los paneles indican la razón de reforzadores, la línea punteada, la indiferencia en la elección (es decir, la razón de respuestas es 1.0 y su logaritmo 0); los cuadrados, los datos promedio en días de no inyección; los círculos, los obtenidos con el vehículo y los triángulos, los correspondientes al haloperidol (Figuras 1 y 2) y al metilfenidato (Figuras 3 y 4).
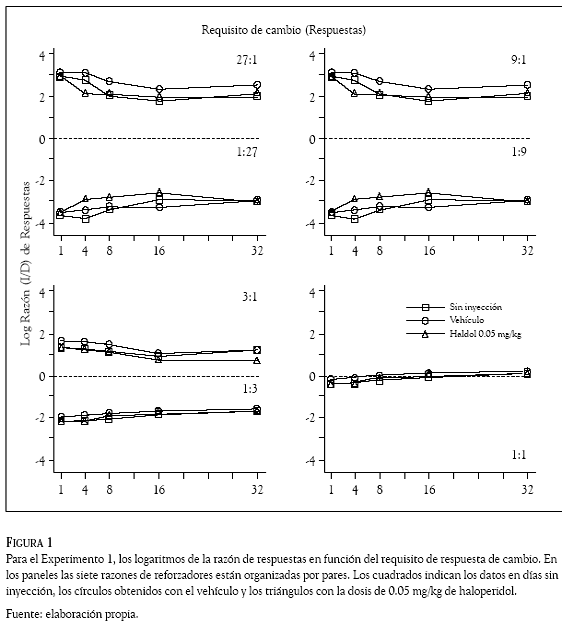
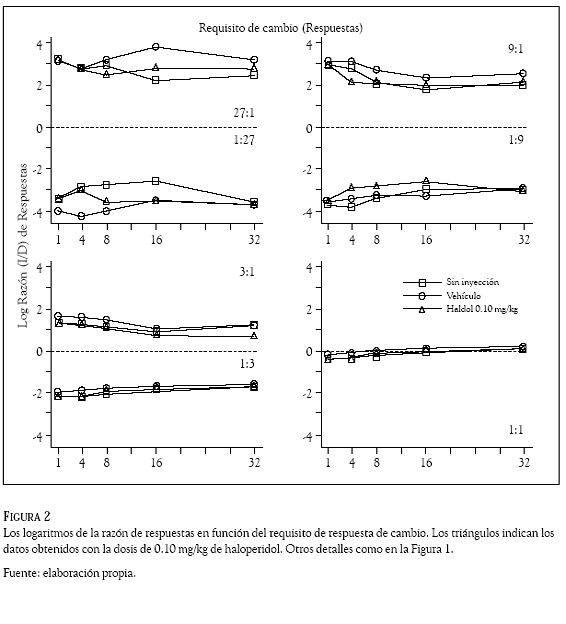
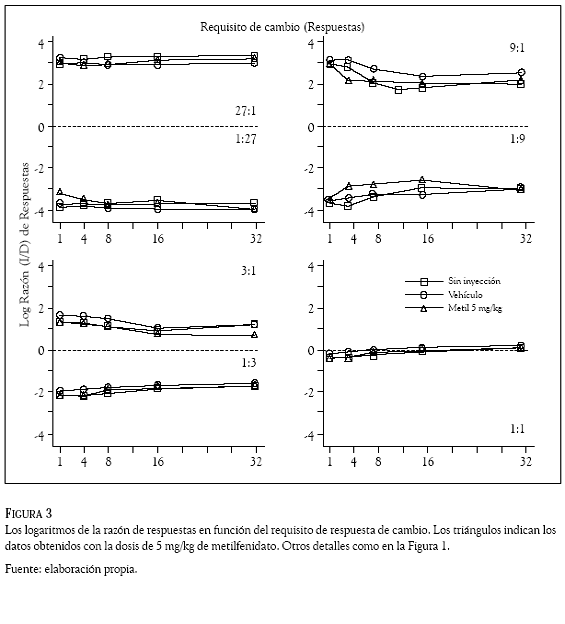
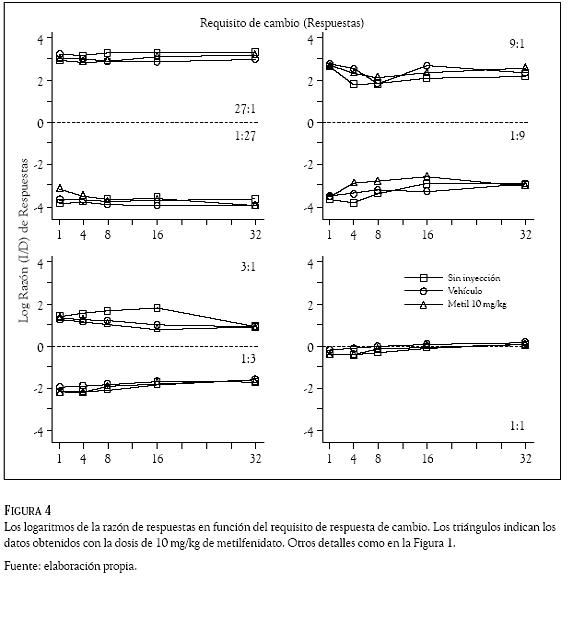
La preferencia de las ratas (en otras palabras, la razón de respuestas) cambió de acuerdo con la probabilidad de reforzamiento que arreglaron las palancas. Cuando ésta fue mayor en la palanca izquierda que en la palanca derecha (razones 27:1; 9:1 y 3:1), las ratas emitieron más respuestas en la palanca izquierda que en la palanca derecha; en esos componentes de reforzamiento las Figuras 1 a 4 muestran valores positivos en los logaritmos de la razón de respuestas, indicando una preferencia por la palanca izquierda en todos los requisitos de cambio de las condiciones sin inyección, vehículo y droga. El resultado opuesto ocurrió cuando la probabilidad de reforzamiento en palanca derecha fue mayor que la palanca izquierda (razones 1:27; 1:9 y 1:3), la preferencia favoreció con más respuestas a la palanca derecha, las Figuras 1 a 4 muestran valores negativos indicando preferencia por esa palanca en todos los requisitos de cambio de las condiciones sin inyección, vehículo y droga. La razón de reforzadores 1:1 (panel inferior derecho) muestra razones de respuestas con valores cercanos a cero (entre 0.125 y – 0.150), revelando una indiferencia de las ratas por responder en una o en la otra palanca.
Las dos dosis de haloperidol (0.05 y 0.10 mg/ kg) y las de metilfenidato (5 y 10 mg/kg) no afectaron la preferencia que las ratas mantuvieron respondiendo a las 7 razones de reforzador con los 5 requisitos de cambio. Una inspección visual de las Figuras 1 a 4 revela que las razones de respuestas generadas en los días de droga (los triángulos) fueron muy similares a las generadas en los días sin droga (cuadrados) y con el vehículo (círculos); la variabilidad en la razón de respuestas que generó la administración del haloperidol y el metilfenidato, no fue considerablemente mayor a la variabilidad en la razón de respuestas que ocasionó el incremento en el requisito de respuesta de cambio.
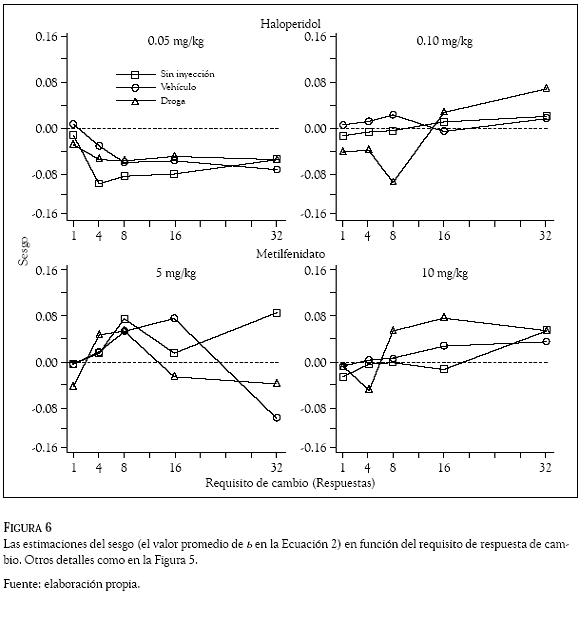
La sensibilidad al reforzamiento y el sesgo de las ratas por una de las dos palancas, se estimó con la Ecuación 2 que se aplicó por separado para cada requisito de cambio con las razones de respuestas obtenidas en los días sin inyección, de vehículo y con la administración de las drogas. De acuerdo con la Ecuación 2, el método de los cuadrados mínimos se utilizó para obtener los valores promedio de la pendiente (del parametro s) y el sesgo (el valor del parametro b) que se graficaron en las figuras 5 y 6, respectivamente, en función del requisito de cambio. Los paneles superiores muestran las estimaciones con las dosis de haloperidol y los inferiores las correspondientes a las dosis de metilfenidato. Los cuadrados indican estimaciones en días sin inyección, los círculos las obtenidas con el vehículo y los tríangulos las estimaciones con las drogas. En la Figura 5 la línea punteada (intersección en 1.0 de la ordenada) indica un empate o igualación entre las razones de respuestas y las razones de reforzadores y en la Figura 6 (intersección en zero de la ordenada), la línea punteada indica la indiferencia de las ratas por responder en cualquiera de las dos palancas.
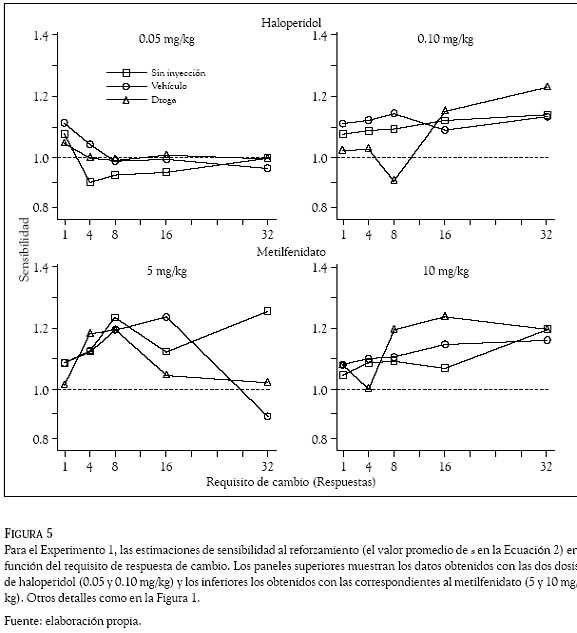
La Figura 5 muestra que en general la sensibilidad al reforzamiento (el valor promedio del parámetros) aumentó con los incrementos sucesivos en el requisito de cambio. Los días sin inyección (los cuadrados) muestran una tendencia de la sensibilidad a incrementar con los incrementos en el requisito de respuesta de cambio. El panel superior izquierdo muestra una excepción en el requisito de cambio de una respuesta, el valor de sensibilidad promedio (1.1) es mayor al generado con los otros requisitos de cambio. Otra excepción la muestra el panel inferior izquierdo en el requisito de 16 respuestas, el valor promedio de s (1.1) es menor al generado (1.22) con el requisito de cambio de 8 respuestas. Las inyecciones del vehículo (los círculos) ocasionaron variabilidad en el valor promedio del parámetros; por ejemplo, en los requisitos de 1 y 4 respuestas el panel superior izquierdo muestra valores promedio de sensibilidad de 1.12 y 1.06, respectivamente; en los requisitos de 8 y 16 respuestas el valor promedio de s permanece en 1.0, pero con el requisito de 32 respuestas el valor de s disminuye a 0.96.
Las inyecciones de haloperidol y metilfenidato no anularon la sensibilidad de las ratas al reforzamiento, en la mayoría de los casos el valor promedio del parámetro s se mantuvo por arriba de 1.0, indicando una sobreigualación de la razón de respuestas a los cambios en la razón de reforzadores. La única excepción la muestra el panel superior derecho en la dosis de 0.10 mg/kg de haloperidol, en el requisito de cambio de 8 respuestas el valor promedio de s es 0.87 (indicando una subigualación), pero en los requisitos de 16 y 32 respuestas, las sensibilidades promedio fueron de 1.18 y 1.22, respectivamente (mostrando una clara sobreigualación). De manera interesante, en la mayoría de los requisitos de cambio y respondiendo a las siete razones de reforzadores, las ratas bajo el efecto de 0.10 mg/kg de haloperidol generaron valores promedio de s que fueron mayores a los obtenidos con la dosis de 0.05 mg/kg (nótese que la dosis 0.05 mg/kg de haloperidol generó valores promedio de s semejantes a los obtenidos en los días de vehículo). El panel inferior derecho muestra un resultado semejante con la dosis de 10 mg/kg de metilfenidato, en la mayoría de los requisitos de cambio esa dosis generó valores promedio del parámetro s que fueron mayores (alrededor 1.2) que los obtenidos (promedio alrededor a 1.1) con la dosis de 5 mg/ kg de metilfenidato. Una comparación entre los paneles inferiores y superiores de la Figura 5, revela que a través de los diferentes requisitos de cambio, las dos dosis de metilfenidato (5 y 10 mg/kg) generaron valores promedio del parámetro s mayores (promedio superior a 1.1) que los generados (promedio cercano a 1.0) por las dosis de haloperidol (0.05 y 0.10 mg/kg).
Para todas las condiciones, los paneles de la Figura 6 muestran un sesgo por la palanca izquierda en el requisito de cambio de una respuesta, nótese los valores positivos en las estimaciones promedio del parámetro b con ese requisito de respuesta de cambio. El panel inferior derecho muestra dos excepciones en las condiciones de vehículo y sin inyección con valores promedió del parámetro b cercanos a cero (e.g., a la indiferencia). El incremento en el requisito de cambio de 1 a 4 respuestas, ocasionó que el sesgo de las ratas por la palanca izquierda cambiara en dirección a la palanca derecha; para la condición de 4 respuestas de cambio la Figura 6 muestra valores negativos en todas las estimaciones promedio del parámetro b. El sesgo de las ratas por la palanca derecha se fortaleció con los requisitos de cambio mayores a 4 respuestas; para las condiciones sin inyección (cuadrados) y vehículo (círculos) los paneles de la Figura 6 muestran valores negativos en las estimaciones del parámetro b en los requisitos de cambio mayores a 4 respuestas. Dos excepciones ocurrieron con los requisitos de 16 y 32 respuestas (paneles del lado derecho) que muestran un sesgo (e.g., valores promedio de b positivos) de las ratas por responder en la palanca izquierda.
Las administraciones de haloperidol y metilfenidato ocasionaron que el sesgo de las ratas por una de las dos palancas variara de manera no sistemática a través de los diferentes requisitos de respuesta de cambio (nótese que el valor promedio de s tomó valores positivos en algunos requisitos de cambio y en otros tomó valores negativos). La variabilidad en los valores promedio del parámetro b fue mayor con las dosis altas de esas drogas (0.10 y 10 mg/kg, respectivamente) que con las dosis bajas (0.05 y 5 mg/kg, respectivamente). De manera interesante, la dosis de 10 mg/kg de metilfenidato ocasionó que el sesgo de las ratas por la palanca derecha cambiara por un sesgo en favor de la palanca izquierda, para esa condición el panel inferior derecho muestra estimaciones promedio del parámetro b con valor positivo en los todos los requisitos de respuesta de cambio.
Experimento 2
El experimento 2 evaluó el efecto del haloperidol y el metilfenidato en una situación de elección que aleatoriamente varió el requisito de respuesta de cambio dentro de una misma sesión. La idea fue modelar un medio ambiente dinámico que combinó el esfuerzo físico que se requiere para cambiar de una alternativa a otra con el posible impedimento motor causado por el haloperidol o bien con la facilitación motriz que se supone ocasiona la administración del metilfenidato. El principal objetivo fue determinar empíricamente si la manipulación aleatoria del requisito de respuesta cambio dentro de una misma sesión, tiene un efecto en la preferencia similar al encontrado en el Experimento 1
Método
Sujetos
Dieciséis ratas albinas macho cepa Wistar de aproximadamente 90 días de edad, sirvieron como sujetos. Las ratas se mantuvieron en las mismas condiciones que en el Experimento 1. Antes de iniciar los estudios, los sujetos fueron divididos aleatoriamente en cuatro grupos de 4 ratas cada uno.
Aparatos
El Experimento 2 utilizó los mismos aparatos descritos en el Experimento 1.
Agentes farmacológicos
En el Experimento 2 los agentes farmacológicos, el método para su preparación, las dosis derivadas de éste y el método de su administración fueron los mismos que se utilizaron en el Experimento 1.
Fase entrenamiento
El Experimento 2 entrenó a las ratas a presionar las palancas utilizando el mismo método de entrenamiento que implementó el Experimento 1.
Procedimiento
El Experimento 2 básicamente implementó el mismo procedimiento que utilizó el Experimento 1, la única diferencia fue que los cinco requisitos de cambio (1, 4, 8, 16 y 32 respuestas) se programaron para ocurrir dentro de una misma sesión. Al inicio de las sesiones la computadora seleccionaba al azar y sin reemplazo a uno de los cinco requisitos de cambio que permanecía vigente por 10 reforzado res que las ratas obtenían respondiendo en las palancas frontales de acuerdo a una razón de reforzadores que el experimentador previamente había seleccionado. La entrega del décimo reforzador ocasionaba que se apagaran las luces arriba de las palancas y la luz general de la caja durante 1 minuto. Al finalizar este tiempo, la computadora seleccionaba al azar a otro requisito de cambio diferente para que controlara los cambios de una palanca a la otra en los siguientes diez reforzadores que terminaban con otro tiempo fuera de 1 minuto. El mismo procedimiento se repitió con los otros requisitos de respuesta de cambio, lo cual significa que la sesión terminaba con la entrega del cincuentavo reforzador (10 reforzadores por cada requisito de cambio) de acuerdo la misma razón de reforzadores.
Para los cuatro grupos de ratas el Experimento 2 implementó el mismo programa de administración de droga que el Experimento 1 (ver Tabla 1). De manera que en seis días consecutivos se evaluaron dos dosis de una misma droga, en cada uno de esos días los cinco requisitos de respuesta de cambio regularon la alternación de palancas que proporcionaron reforzadores de acuerdo a una misma razón de reforzadores. Esto significa que se necesitaron 42 días para evaluar las dos drogas en las siete razones de reforzadores (6 días por cada razón de reforzadores).
Análisis de los datos
El mismo análisis de datos que implementó el Experimento 1, se efectuó en el Experimento 2 que también incluyó el uso de la Ecuación 1 para estimar el efecto de las drogas en la motivación por la comida (el parámetro s) y en la motricidad del organismo (el parámetro b).
Resultados
Los logaritmos de las razones de respuestas (izquierda /derecha) se graficaron en las Figuras 7 a 10 en función del requisito de respuesta de cambio. Los números en los paneles indican la razón de reforzadores, la línea punteada la indiferencia en la elección, los cuadrados los datos promedio en días de no inyección, los círculos los obtenidos con el vehículo y los triángulos los correspondientes al haloperidol (Figuras 7 y 8) y al metilfenidato (Figuras 9 y 10).
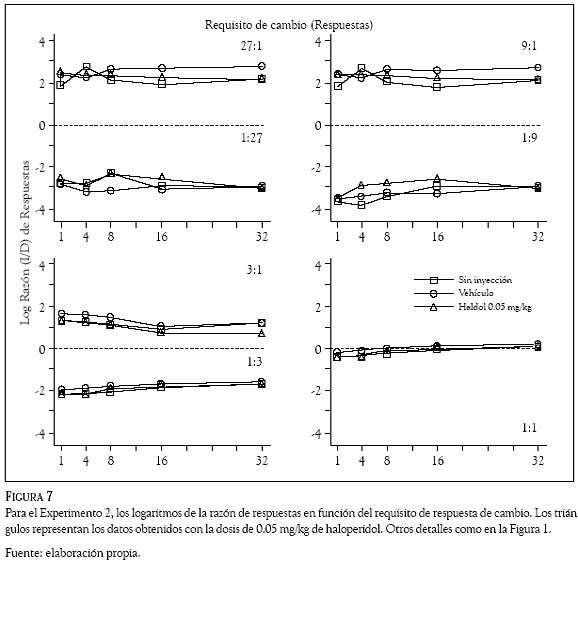
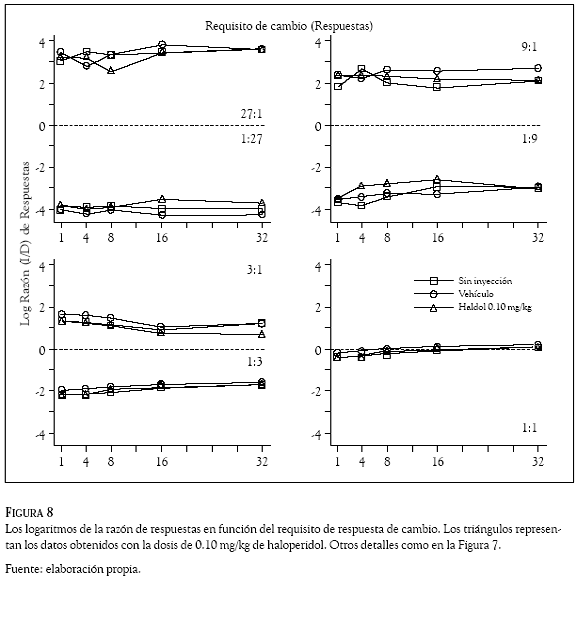
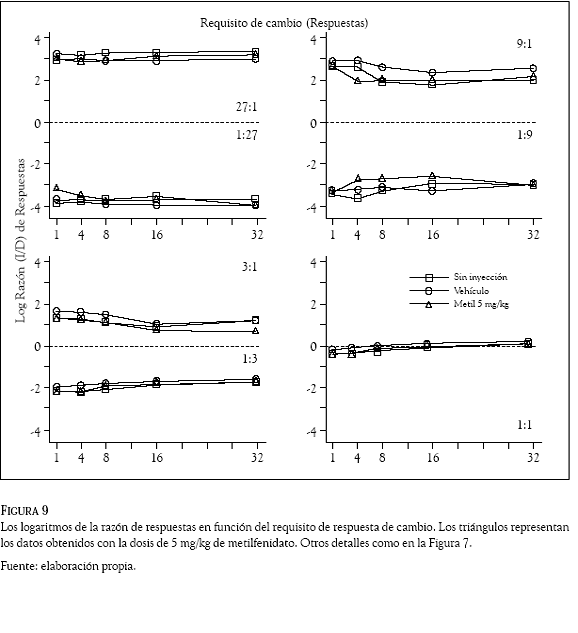
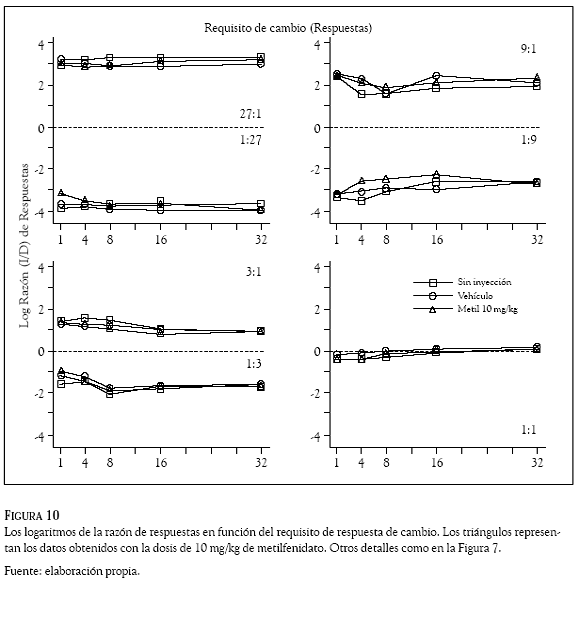
De manera consistente con los datos del Experimento 1, la preferencia de las ratas cambió en función de la probabilidad de reforzamiento asociada a las dos palancas. En las razones de reforzadores de 27:1; 9:1 y 3:1 las Figuras 7 a 10 muestran que las ratas emitieron más respuestas en la palanca izquierda que en la palanca derecha; en esos componentes, los valores positivos en los logaritmos de la razón de respuestas indican una preferencia de las ratas por la palanca izquierda en todos los requisitos de cambio de las condiciones sin inyección, vehículo y droga. En contraste, en las razones de reforzadores de 1:27; 1:9 y 1:3 la preferencia favoreció con más respuestas a la palanca derecha, para estos componentes los paneles muestran valores negativos en todos los requisitos de cambio de las condiciones sin inyección, vehículo y droga. Para la razón de reforzadores de 1:1, los paneles inferiores muestran valores promedio de las razones de respuestas cercanos a cero (entre 0.160 y – 0.290), indicando la indiferencia de las ratas por responder en la palanca derecha o en la palanca izquierda.
Las dosis intraperitoneales de haloperidol (0.05 y 0.10 mg/kg) y metilfenidato (5 y 10 mg/kg), no afectaron las preferencias que las ratas mostraron respondiendo a las 7 razones de reforzadores en los 5 requisitos de cambio. Una inspección visual de las Figuras 7 a 10 revela que los logaritmos de las razones de respuestas obtenidos con las dos drogas (los triángulos) tienen valores similares a los que muestran las condiciones sin inyección (cuadrados) y de vehículo (círculos); la variabilidad en la distribución de las respuestas generada por la administración del haloperidol y el metilfenidato, no es considerablemente mayor que la variabilidad en las razones de respuestas que los requisitos de cambio ocasionaron en las condiciones sin inyección y de vehículo.
De la misma manera que en el Experimento 1, la Ecuación 2 se aplicó por separado para cada requisito de cambio con las razones de respuestas obtenidas en los días sin inyección, de vehículo y con la administración de las drogas. Los valores promedio del parámetro s (sensibilidad al reforzamiento) y los correspondientes al sesgo (valor del parametro b), se graficaron en las Figuras 11 y 12, respectivamente, en función de los requisitos de cambio. Los paneles superiores muestran las estimaciones obtenidas con las dosis de haloperidol (0.05 y 0.10 mg/kg) y los inferiores las correspondientes a las dosis de metilfenidato (5 y 10 mg/kg). Los cuadrados representan las estimaciones en los días sin inyección, los círculos las obtenidas con el vehículo y los tríangulos las evaluaciones con las drogas. En la Figura 11 la línea punteada (intersección en 1.0 de la ordenada) indica un empate o igualación entre las razones de respuestas y las razones de reforzadores y en la Figura 12 (intersección en zero de la ordenada), la línea punteada indica la indiferencia de las ratas por responder en cualquiera de las dos palancas.
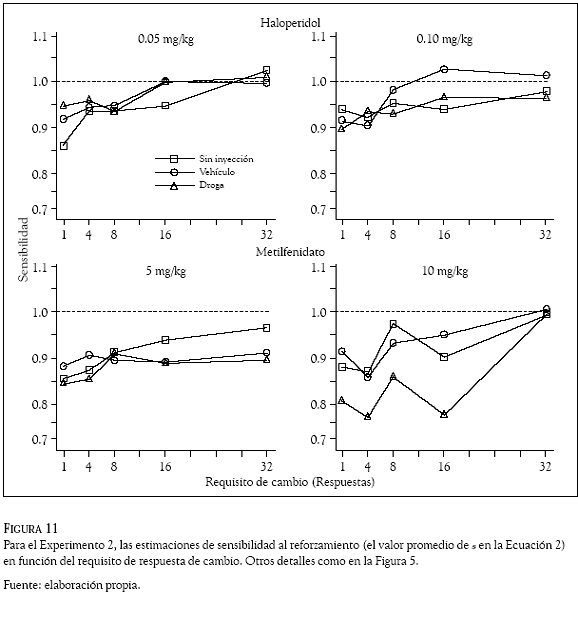
En las condiciones, la Figura 11 muestra que la sensibilidad al reforzamiento (valor promedio del parámetro s) aumentó con los incrementos en el requisito de respuesta de cambio. La dosis de metilfenidato de 10 mg/kg generó variabilidad en la sensibilidad al reforzamiento, para esta condición el panel inferior derecho muestra un valor promedio de 0.8 en los requisitos de cambio de 1, 4, 8 y 16 respuestas; sin embargo, con el requisito de cambio de 32 respuestas el valor promedio del parámetro s fue de 1.0, indicando una igualación (empate) entre la distribución de las respuestas y la distribución de los reforzadores obtenidos en las palancas.
Las estimaciones de sensibilidad al reforzamiento con las dos dosis de haloperidol (0.05 y 0.10 mg/kg) y con la dosis de 5 mg/kg de metilfenidato, muestran valores muy similares a los obtenidos con el vehículo, indicando que la motivación de las ratas por la comida no cambió con la administración de las drogas. A diferencia del Experimento 1 donde las dos dosis de metilfenidato generaron valores del parámetro s más altos (mayores a 1.2) que los generados por las dos dosis de haloperidol (cercanos a 1.0), en el Experimento 2 las dos drogas generaron valores promedio de sensibilidad menores a 1.0, indicando una clara subigualación entre las razones de respuestas y las razones de reforzadores. Las excepciones ocurrieron con los requisitos de cambio de 16 y 32 respuestas con la dosis de 0.05 mg/kg de haloperidol (panel superior izquierdo) que muestran un valor promedio de 1.0 en el parámetro s (una perfecta igualación). De manera interesante, las ratas respondiendo bajo el efecto de 10 mg/kg de metilfenidato a un requisito de cambio de 32 respuestas, distribuyeron su conducta en las dos palancas de manera que los cambios en las razones de respuestas correspondieron a los cambios en las razones los reforzadores que arreglaron las palancas en los 7 componentes; para esta condición el panel inferior derecho muestra una perfecta igualación (e.g., un valor de 1.0 en el parámetro s). Una comparación entre los paneles inferiores y los superiores de la Figura 11, revela que a través de los diferentes requisitos de cambio la dos dosis (5 y 10 mg/kg) de metilfenidato generaron valores promedio del parámetro s más bajos (entre 0.84 y 0.96) que los generados (entre 0.90 y 1.0) por las dos dosis (0.05 y 0.10 mg/kg) de haloperidol.
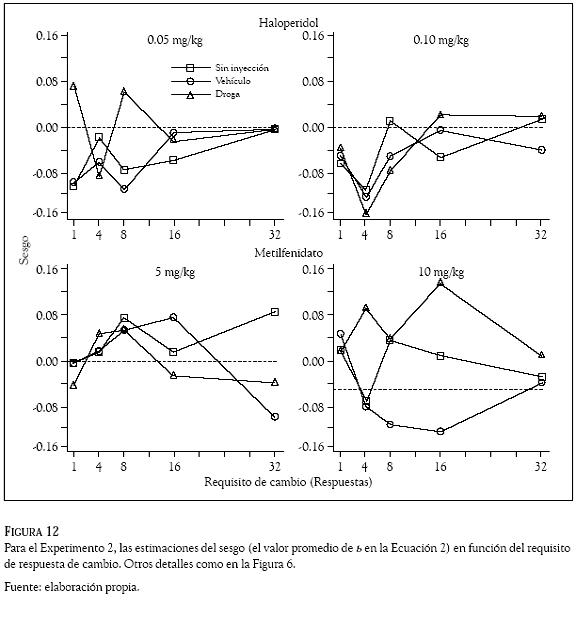
En la mayoría de las condiciones, la Figura 12 muestra un sesgo de las ratas por la palanca derecha (la mayoría de los símbolos está debajo de la línea de indiferencia), nótese que a través de los diferentes requisitos de cambio las estimaciones promedio del parámetro b tomaron valores negativos. Algunas excepciones a este resultado las muestra el requisito de cambio de una respuesta en las condiciones sin inyección y de vehículo con valores positivos en el valor promedio de b, indicando un sesgo inicial de las ratas por la palanca izquierda. En general, la administración de las drogas ocasionó que el sesgo inicial de las ratas por la palanca derecha cambiara en dirección a la palanca izquierda.
Aunque la Figura 12 muestra variabilidad en los valores promedio del parámetro b obtenidos con las dos dosis (0.05 y 0.10 mg/kg) de haloperidol y las de metilfenidato (5 y 10 mg/kg); en esas condiciones y a través de los diferentes requisitos de cambio las estimaciones del parámetro b muestran más casos con valores positivos que con valores negativos, indicando que bajo el efecto de las drogas las ratas desarrollaron un sesgo por la palanca izquierda. De manera interesante, la dosis altas de haloperidol (0.10 mg/kg) y metilfenidato (10 mg/kg) ocasionaron que el sesgo de las ratas por la palanca izquierda cambiara en dirección a la indiferencia, nótese que en esas condiciones los triángulos están sobre la línea de indiferencia.
Discusión General
Los resultados de los Experimentos 1 y 2 extendieron la generalidad de los hallazgos reportados por Aparicio, Velasco y Balderrama (2004) a medios ambientes de reforzamiento dinámico. Los dos experimentos modelaron las contingencias de reforzamiento que operan en ambientes naturales, ya que arreglaron cambios variables en las distribuciones de reforzadores (de un día a otro) y en los requisitos de respuesta de cambio (un RCA diferente cada 10 reforzadores o de una sesión a otra). La rápida adaptación de las ratas a los cambios dinámicos en las contingencias de reforzamiento, fue evidenciada por los cambios sistemáticos en la preferencia (la razón de respuestas) que las ratas mostraron en los 7 componentes respondiendo a los 5 requisitos de cambio (Figuras 1 a 4 y 7 a 10). En todos los casos, la razón de respuestas correspondió con la razón de reforzadores que arreglaron los 7 componentes de reforzamiento. Esto es, las ratas rastrearon de un reforzador a otro a la palanca asociada a la mayor probabilidad de reforzamiento y ahí emitieron el mayor número de respuestas.
Ninguna de las dos drogas, haloperidol (0.05 y 0.10 mg/kg) y metilfenidato (5 y 10 mg/kg) en las dosis administradas impidieron que las ratas se adaptaran rápidamente a los cambios dinámicos en las contingencias de reforzamiento que arreglaron las dos palancas. En los días sin inyección y de administración del vehículo, las distribuciones de respuestas fueron muy similares a las que se observaron con las ratas respondiendo a las palancas bajo el efecto de las drogas. La idea de que el haloperidol y el metilfenidato interfieren con conductas relacionadas con el consumo de alimento, como la velocidad de alimentación, frecuencia de ésta y su manipulación (Koob, Riley, Smith & Robbins, 1978), no recibió apoyo de los presentes estudios que claramente mostraron que el haloperidol y el metilfenidato no anularon la motivación de las ratas por la comida (Salomone, Arizzi, Sandoval, Cervone & Aberman, 2002; Salamone, Mahan & Rogers, 1993).
Las administraciones de halperidol y metilfenidato causaron que la preferencia de las ratas variara de manera no sistemática con las manipulaciones en el requisito de cambio, sugiriendo que estas drogas actuaron sobre el sistema motor de las ratas que permanecieron en la palanca de baja probabilidad de reforzamiento por verse impedidas para completar el número de respuestas requeridas para cambiar a la palanca de mayor probabilidad de reforzamiento. Esta idea recibe apoyo de los estudios que muestran que el bloqueo de la acción inhibitoria de receptores D2 disminuye la actividad del sistema motor debido a una acción extrapiramidal (EEP) en el ganglio basal que afecta la actividad de dopamina en la corteza del núcleo acumbens que se encarga de regular las funciones motoras (Beninger, 1983; Berridge & Robinson, 1998; Fellous & Suri, 2002; Marcus, Malmerfelt, Nyberg & Svensson, 2002; Mobini, Chiang, Bradshaw & Szabani, 2000; Wise, 1982; Wise & Bozarth, 1987).
Son diversos los estudios que han documentado una disminución en la actividad locomotriz del organismo debida a la administración aguda de neurolépticos como el haloperidol (e.g., Aparicio, 1999, 2001; Aparicio & Velasco, 2003; Berridge & Robinson, 1998; Ettenberg, 1989; Fellous & Suri, 2002; Fouriezos, Hansson & Wise, 1978; Fouriezos & Wise, 1976; Franklin & McCoy, 1979; Gallistel, Boytin, Gomita & Klebanoff, 1982; Liebman & Butcher, 1973; Miller, Wickens & Beninger, 1990; Wise, 1982; Wise & Colle, 1984; Wise & Raptis, 1986; Wise, Spindler, deWill & Gerber, 1978). El caso del metilfenidato es diferente porque es un agonista a la dopamina que actúa para incrementar la actividad extracelular en el estriado y núcleo acumbens que son las regiones del cerebro involucradas en la locomoción (Kuczenski & Segal, 2002). La interpretación se complica porque las mismas dosis de metilfenidato que incrementa la locomoción espontánea del organismo, suprimen su conducta operante, sugiriendo que la hiperactividad motriz puede interferir con la emisión de las conductas operantes (Mobini, Chiang, Bradshaw & Szabani, 2000). Sabemos que en dosis orales terapéuticas, el metilfenidato bloquea más de 50% del transportador de dopamina o DAts (Volkow, Fowler & Wang, 1999) aumentando el nivel extracelular de dopamina (Volkow & Fowler, 2000), esta acción se asocia con las propiedades reforzantes de las drogas. Un denominador común entre el haloperidol y el metilfenidato es que las dos drogas alteran la actividad de dopamina, el haloperidol la disminuye y el metilfenidato la aumenta, estas alteraciones en la actividad de dopamina producen irregularidades en la emisión de conductas operantes que se manifiestan en cambios no sistemáticos en la preferencia (la distribución de las respuestas) sin que esto afecte la motivación de las ratas por el alimento (Aparicio, Velasco & Balderrama, 2004; Tombaugh, Tombaugh & Anisman, 1979; Wise & Bozarth, 1987).
En las condiciones sin inyección y de vehículo, el Experimento 1 mostró estimaciones del parámetro s (sensibilidad al reforzamiento) con valores superiores a 1.0, lo cual significa que los cambios en las razones de respuestas fueron más extremos que los cambios en las razones de reforzadores, fenómeno que se conoce como sobreigualación. Este resultado es consistente con el reportado en situaciones de elección que también manipularon el requisito de respuesta de cambio con ejecuciones concurrentes en estado estable (Pliskoff & Fetterman, 1981; Todorov, Acuña & Falcón, 1982) y en transición (Aparicio & Otero, 2004; Aparicio & Balderrama, 2004), lo cual contrasta con la subigualación que es el resultado encontrado con mayor frecuencia en estudios de elección con pichones (Baum, 1979).
La sensibilidad de las ratas al reforzamiento no disminuyó con las administraciones de haloperidol y metilfenidato, por el contrario las estimaciones del parámetro s mostraron valores superiores a 1.0 indicando la sobreigualación. Este resultado sugiere que el costo que los requisitos de cambio le impusieron a la conducta de cambiar de una palanca a la otra, incrementó con las dos dosis de haloperidol y las de metilfenidato, confirmando que cuando la situación de elección le impone un costo adicional a la conducta de desplazarse de un lugar a otro (moverse bajo el efecto de las drogas en el presente estudio) se incrementa la sensibilidad del organismo al reforzamiento (Aparicio, 2001; Aparicio & Balderrama, 2004; Aparicio & Otero, 2004).
La administración de una dosis de 0.10 mg/kg de haloperidol, ocasionó valores en el parámetro s cercanos o mayores a 1.2 en los requisitos de cambio de 16 y 32 respuestas. En contraste, las estimaciones de sensibilidad al reforzamiento con la dosis de 0.05 mg/kg de haloperidol, mostraron valores de 1.0 en todos los requisitos de cambio. Este resultado sugiere que fue la experiencia de las ratas en la situación de elección (recuérdese que la segunda dosis se evaluó después de muchas sesiones) lo que contribuyó a que aumentara la sensibilidad al reforzamiento (Todorov et al., 1983).
En los dos experimentos, el sesgo de las ratas por una de las dos palancas varió de manera no sistemática con las inyecciones de haloperidol y metilfenidato, sugiriendo un efecto de las drogas en el sistema motor (un incremento en el costo del traslado), un resultado ya documentado en estudios que evaluaron el efecto de agonistas y antagonistas a la dopamina en conductas de elección (Heyman, 1983; Heyman & Monaghan, 1987).
En el Experimento 2, los valores de las estimaciones de sensibilidad y sesgo obtenidos con las dos dosis de haloperidol (0.05 y 0.10 mg/kg), no fueron notoriamente diferentes a los valores que se obtuvieron con las dosis de metilfenidato (5 y 10 mg/kg), las dos drogas causaron que las ratas visitaran con más frecuencia la palanca de menor probabilidad de reforzamiento porque el cambio a la otra palanca requería de un número considerable de respuestas, lo cual generó valores menores a 1.0 en el parámetro s (poca sensibilidad al reforzamiento) y variaciones en el parámetro b (oscilaciones en el sesgo) a través de los 7 componentes de reforzamiento. Este resultado es consistente con los hallazgos reportados en situaciones de elección que evaluaron los efectos de antagonistas a la dopamina con técnicas de depleción; en esos estudios la preferencia de las ratas por la alternativa que proporcionaba reforzadores de alto valor, cambio por la alternativa que proporcionaba reforzadores de bajo valor que requerían de menor esfuerzo para ser producidos (e.g., Aparicio & Barajas, 2002; Cousins, Atherton, Turner & Salamone, 1996; Heyman, 1983; Nowend, Arrizzi, Carlson & Salamone, 2001; Salamone, Steinpreis, McCullough, Smith, Grebel & Mahan, 1991).
En conjunto, los presentes resultados son similares a los encontrados en estudios que evaluaron agonistas y antagonistas a la dopamina (el metilfenidato) en la misma situación operante. Por ejemplo, Heyman (1983) midió cambios en los parámetros libres (k y r0) de la ley de la igualación de Herrnstein (1974) causados por la administración de drogas agonistas y antagonistas a la dopamina. En sus estudios, el parámetro k se utilizó para estimar los cambios en la capacidad motora y el parámetro r0 para medir los cambios en la sensibilidad al reforzamiento (Heyman, 1983; Wise & Bozarth, 1987). Aunque los resultados no fueron consistentes a través de las diferentes condiciones, él encontró que las dosis intermedias de agonistas a la dopamina ocasionaron incrementos en la tasa de respuestas, pero las dosis altas de esas drogas causaron disminuciones en la tasa de respuestas; en su interpretación de esos resultados, Heyman (1983) aludió a un déficit motor causado por un incremento en la actividad de la dopamina producto de la administración de una dosis alta de un agonista a la dopamina. Sin embargo, son muchos los factores que pueden alterar la actividad motora del organismo. Por ejemplo, en situaciones en donde se presentan dos tipos de alimento diferente, el organismo puede presentar una conducta estacionaria en el lugar donde se encuentra una de esas comidas, esto podría ser interpretado como un decremento en la actividad motriz causado por la administración de la droga. Otra variable que podría contribuir a una interpretación errónea del efecto de las drogas agonistas y antagonistas a la dopamina en el sistema motor, es un incremento exponencial en el requisito de respuestas (como en los programas de razón fija). En estos casos, el organismo deja de responder a requisitos de razón muy altos, un fenómeno conocido como “el estiramiento del requisito de razón”.
Consistente con el trabajo conducido en nuestro laboratorio (Aparicio, 1998, 1999, 2001; Aparicio & Velasco, 2003; Aparicio, Velasco & Balderrama, 2004), el presente estudio utilizó con éxito los parámetros libres (s y b) de la ley de igualación generalizada para estimar los efectos de agonistas y antagonistas a la dopamina en el sistema motor y la motivación de las ratas por el reforzador, respectivamente. Nuestros resultados confirmaron que el procedimiento de elección dinámica (e.g., Aparicio & Balderrama, 2004; Aparicio & Barajas, 2002; Aparicio & Otero, 2004; Davison & Baum, 2000) es un método idóneo para evaluar los efectos de drogas agonistas y antagonistas a la dopamina, porque éste permite estudiar la adaptación del organismo a condiciones ambientales similares a las que se presentan en medios ambientes naturales (e.g., en la búsqueda de alimento). Trabajos posteriores al presente estudio podrían evaluar los efectos crónicos de estas drogas utilizando la misma metodología, lo cual permitiría medir cambios permanentes en la preferencia del organismo. También, sugerimos que el procedimiento de Davison y Baum (2000), se utilice para evaluar drogas que se asocian a la supresión o interés de los sujetos por el alimento, como es la anfetamina y la sibutramina que disminuyen el apetito afectando la búsqueda alimento y ocasionando reducción del peso corporal del organismo.
Conclusiones
El objetivo del presente estudio fue evaluar los efectos de una droga antagonista (haloperidol) y otra agonista (metilfenidato) a la dopamina en la preferencia de las ratas, particularmente en la motivación por la comida y en el sistema motor. Los resultados mostraron que el haloperidol y el metilfenidato, administrados por vía intraperitoneal, actuaron de manera similar en el sistema motor del organismo ocasionando cambios no sistemáticos en el valor del parámetro b (el sesgo) de la ley de igualación generalizada (Baum, 1974). Sin embargo, ninguna de las dos drogas, en las dos dosis administradas, afectó la sensibilidad de ratas al reforzamiento; el valor del parámetro s de la ley de igualación generalizada no disminuyó con el haloperidol y ni con el metilfeninato, por el contrario la sensibilidad al reforzamiento aumentó con incrementos en las dosis de estas drogas. Los resultados de los Experimentos 1 y 2 mostraron que el haloperidol y el metilfenidato no interfirieron con la adaptación de las ratas a las contingencias de reforzamiento, las razones de respuestas mantenidas por las 7 razones de refozadores no cambiaron como resultado de la administración de las drogas. En todas las condiciones (libres de droga y con droga) los valores del parámetro s (sensibilidad) incrementaron en función del requisito de cambio. Este resultado descartó la idea de que el haloperidol elimina el interés (la motivación) de las ratas por la comida, el Experimento 1 mostró valores del parámetro s mayores a 1.0 con las administraciones del haloperidol y el Experimento 2, confirmó ese resultado mostrando valores del parámetro s cercanos a 1.0 con el haloperidol, que fueron muy similares a los obtenidos en las condiciones sin inyección y de vehículo. Por tanto, concluimos que los parámetros, s y b, de la ley de igualación generalizada son útiles para estimar efectos de agonistas y antagonistas a la dopamina en la motivación por el alimento y en el sistema motor, respectivamente.
Referencias
Aparicio, C. F. (1998). Assessing haloperidol in rats with the Barrier choice paradigm. Suma Psicologica, 5,11, 1-20. [ Links ]
Aparicio, C. (1999). The barrier choice paradigm: Haloperidol reduces sensitivity to reinforcement. Behavioural Processes, 48, 57-67. [ Links ]
Aparicio, C. (2001). Adicción a drogas antipsicosis: una evaluación de los efectos colaterales. Anuario de Investigación en Adicciones, 2, 34-58. [ Links ]
Aparicio, C. & Balderrama, J. (2004). Elección, reforzamiento y requisitos de respuesta de cambio. Universitas Pychologica, 3(2), 121-146. [ Links ]
Aparicio, C. & Barajas, I. (2002). Cambios dinámicos en contingencias de reforzamiento en situación de elección: ¿se requiere la estabilidad en la ejecución concurrente? Revista Mexicana de Análisis de la Conducta, 28, 67-90. [ Links ]
Aparicio, C. & Otero, E. E. (2004). Requisitos de cambio y sensibilidad al reforzamiento en medios ambientes semi-estables y dinámicos. Revista Mexicana de Análisis de la Conducta, 1(30), 23-78. [ Links ]
Aparicio, C. & Velasco, F. (2003). El paradigma de elección con barrera: evaluación del haloperidol con ocho alternativas de respuestas y dos requisitos de traslado. Universitas Pychologica, 2(2), 109-135. [ Links ]
Aparicio, C., Velasco, F. & Balderrama, J. (2004). Haloperidol, elección y requisitos de respuesta de cambio. Suma Psicológica, 11(2), 181-204. [ Links ]
Baum, W. M. (1974). On two type of deviation from the matching law: Bias and undermatching. Journal of the Experimental Analysis of Behavior, 22, 231-242. [ Links ]
Baum, W. M. (1979). Matching, undermatching, and overmatching in studies of choice. Journal of the Experimental Analysis of Behavior, 32, 269-281. [ Links ]
Beninger, R. J. (1983). The role of dopamine in locomotor activity and learning. Brain Research, 368, 62-68. [ Links ]
Berridge, K. C. & Robinson, T. E. (1998). What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Research Reviews, 28, 309-369. [ Links ]
Bradshaw, C. M., Ruddle, H. V. & Szabadi, E. (1981). Relationship between response rate and reinforcement frequency in variable-interval schedule: III. The effect of d-amphetamine. Journal of the Experimental Analysis of Behavior, 36, 29-39. [ Links ]
Clark, F. C. & Steele, B. J. (1966). Effects of d-amphetamine on performance under a multiple schedule in the rat. Psychopharmacology, 9, 157-169. [ Links ]
Cousins, M. S., Atherton, A., Turner L. & Salamote, J. D. (1996). Nucleus accumbens dopamine depletions alter relative response allocation in a T-maze cost/benefit task. Behavior Brain Research, 74, 189-197. [ Links ]
Cousins, M. S., Wei, W. & Salomone, J. D. (1994). Pharmacological characterization of performance on a concurrent lever pressing/feeding choice procedure: Effects of dopamine antagonist, cholinomimetic, sedative and stimulant drugs. Psychopharmacology, 116, 529-537. [ Links ]
Davison, M. C. & Baum, W. M. (2000). Choice in a variable environment: Every reinforcer counts. Journal of the Experimental Analysis of Behavior, 74, 1-24. [ Links ]
Dews, P. B. (1958). Studies of behavior: IV. Stimulant actions of methamphetamine. Journal of Pharmacology and Experimental Therapeutics, 122, 137-147. [ Links ]
Dews, P. B. & Morse, W. H. (1961). Behavioral pharmacology. In W. C. Cutting, R. H. Driesbach & H. W. Elliot (Eds.), Annual review of pharmacology (pp. 145-174). Palo Alto: Annual Reviews Inc. [ Links ]
Ettenberg, A. (1989). Dopamine, neuroleptics, and reinforced behavior. Neuroscience and Biobehavior Reviews, 13, 105-111. [ Links ]
Fellous J. M. & Suri, R. E. (2002). The roles of Dopamine. In M. A. Arbib (Ed.), The Handbook of Brain Theory and Neural Networks (2d ed.). Cambridge, MA: MIT Press. [ Links ]
Fouriezos, G., Hansson, P. & Wise, R. A. (1978). Neuroleptic- induce attenuation of brain stimulation reward. Journal of Comparative and Physiological Psychology, 92, 659-669. [ Links ]
Fouriezos, G. & Wise, R. A. (1976). Pimozide-induce extinction of intracanial self-stimulation: Response patterns rule out motor or performance deficit. Brain Research, 103, 377-380. [ Links ]
Franklin, K. B. J. & McCoy, S. N. (1979). Pimozideinduced extinction in rats: Stimulus control of responding rules out motor deficit. Pharmacology Biochemistry & Behavior, 11, 71-76. [ Links ]
Gallistel, C. R., Boytin, M., Gomita, Y. & Klebanoff, L. (1982). Does pimozide block the reinforcing effect of brain stimulation? Pharmacology, Biochemistry and Behavior, 17, 769-781. [ Links ]
Heffner, T. G., Drawbaugh, R. B. & Zigmond, M. J. (1974). Amphetamine and operant behavior in rats: Relationship between drug effect and control response rate. Journal of Comparative and Physiological Psychology, 86, 1031-1043. [ Links ]
Herrnstein, R. J. (1974). Formal properties of the matching law. Journal of the Experimental Analysis of Behavior, 21, 159-164. [ Links ]
Heyman, G. M. (1983). A parametric evaluation of the hedonic and motoric effects of drugs: pimozide and amphetamine. Journal of the Experimental Analysis of Behavior, 40, 113-122. [ Links ]
Heyman, G. M. & Monaghan, M. M. (1987). Effects of changes in response requirement and deprivation on the parameters of the matching law equation: New data and review. Journal of Experimental Psychology: Animal Behavior Processes, 13, 384-394. [ Links ]
Kelleher, R. T. & Morse, W. H. (1968). Determinants of the specificity of behavior effects of drugs. Ergebnisse der Physiologie, 60, 1-56. [ Links ]
Koob, G. F., Riley, S. J., Smith, S. C. & Robbins, T. W. (1978). Effects of 6-hydroxydopamine lesions of the nucleus accumbens septi and olfactory tubercle on feeding, locomotor activity, and anphetamine anorexia in the rat. Journal of Comparative and Physiological Psychology, 92, 917-927. [ Links ]
Kuczenski, R. & Segal, D. S. (2002). Exposure of adolescent rats to oral methylphenidate: Preferential effects on extracellular norepinephrine and absence of sensitization and cross-sensitization to methamphetamine. Journal of Neuroscience, 22, 7264-71. [ Links ]
Liebman, J. M. & Butcher, L. L. (1973). Effects on self-stimulation behavior of drugs influencing dopaminergic neurotransmission mechanisms. Naunyn-Schmiedeberg’s Archives of Pharmacology, 227, 305-318. [ Links ]
Lucki, I. (1983). Rate-dependent effects of amphetamine on responding under ramdom-interval schedule of reinforcement in the rat. Pharmacology, Biochemistry and Behavior, 18, 195-201. [ Links ]
Lucki, I. & DeLong, R. E. (1983). Control rate of response or reinforcement and amphetamine’s effect on behavior. Journal of the Experimental Analysis of Behavior, 40, 123-132. [ Links ]
Marcus, M. M., Malmerfelt, A., Nyberg, S. & Svensson, T. H. (2002). Biochemical effects of low doses of haloperidol are qualitatively similar to those of high doses. European Neuropsychopharmacology, 12, 379-386. [ Links ]
McMillan, D. E. (1968). Some interactions between sympathomimetic amines and amine-depleting agents on the schedule-controlled behavior of the pigeon and the squirrel monkey. Journal of Pharmacology and Experimental Therapeutics, 163, 172-187. [ Links ]
Miller, R., Wickens, J. R. & Beninger, R. J. (1990). Dopamine D-1 and D-2 receptors in relation to reward and performance: A case for the D-1 receptors as a primary site of therapeutic action of neuroleptic drugs. Progress in Neurobiology, 34, 143-183. [ Links ]
Mobini, S., Chiang, T. J., Bradshaw, C. M. & Szabani, E. (2000). Comparison of the effects of clozapine, haloperidol, chlorpromazine and d-amphetamine on performance on a time-constrained progressive ratio schedule and on locomotor behavior in the rat. Psychopharmacology, 152, 47-54. [ Links ]
Nowend, K. L., Arrizzi, M., Carlson, B. B. & Salamone, J. D. (2001). D1 o D2 antagonism in nucleus accumbens core or dorsomedial shell suppresses lever pressing for food but leads to compensatory increases in chow consumption. Pharmacology, Biochemistry and Behavior, 18, 373-382. [ Links ]
Owen, J. E. (1960). The influence of dt, d, y l-amphetamine and d-methamphetamina on a fixed-ratio schedule. Journal of the Experimental Analysis of Behavior, 3, 293-310. [ Links ]
Pliskoff, S. S. & Fetterman, J. G. (1981). Undermatching and overmatching: The fixed-ratio changeover requirement. Journal of the Experimental Analysis of Behavior, 36, 21-27. [ Links ]
Salomone, J. D., Arizzi, M. N., Sandoval, M. D., Cervone, K. M. & Aberman, J. E. (2002). Dopamineantagonists alter response allocation but do no suppress appetite for food in rats: contrast between the effects of SKF 83566, raclopride, and fenfluramine on a concurrent choice task. Psychopharmacology, 160, 371-380. [ Links ]
Salamone, J. D., Mahan, K. & Rogers, S. (1993). Ventrolateral striatal dopamine depletions impair feeding and food handling in rats. Pharmacology Biochemistry & Behavior, 44, 605-610. [ Links ]
Salamone, J. D., Steinpreis, R. E., McCullough, L. D., Smith, P., Grebel, D. & Mahan, K. (1991). Haloperidol and nucleus accumbens dopamine depletion suppress lever pressing for food but increase free food consumption in a novel food-choice procedure. Psychopharmacology, 104, 515-521. [ Links ]
Stubbs, D. A. & Pliskoff, S. S. (1969). Concurrent responding with fixed relative rate of reinforcement. Journal of the Experimental Analysis of Behavior, 12, 887-895. [ Links ]
Todorov, J. C., Acuña Santaella, L. E. & Falcón Sanguinetti, O. (1982). Concurrent procedures, changeover delay and the choice behavior of rats. Mexican Journal of Behavior Analysis, 2(8), 133-147. [ Links ]
Todorov, J. C., Oliveira-Castro, J. M., Hanna, E. S., Bittencourt de Sa, M. C. N. & Barreto, M. Q. (1983). Choice, experience, and the generalized matching law. Journal of the Experimental Analalysis of Behavvior, 40, 99-111. [ Links ]
Tombaugh, T. N., Tombaugh, J. & Anisman, H. (1979). Effects of dopamine receptor blockade on alimentary behavior: Home cage food consumption, magazine training, operant acquisition, and performance. Psychopharmacology, 66, 219-225. [ Links ]
Volkow. N. D., Fowler. J. S. & Wang. G. J. (1999). Imaging studies on the role of dopamine in cocaine reinforcement and addiction in humans. Journal of Psychopharmacology, 13, 337–345. [ Links ]
Volkow, N. D. & Fowler, J. S. (2000). Addiction, a disease of compulsion and drive: Involvement of the orbitofrontal cortex. Cerebral Cortex, 10, 318–325. [ Links ]
Wise, R. A. (1982). Neuroleptics and operant behavior: The anhedonia hypothesis. Behavior Brain Science, 5, 39-87. [ Links ]
Wise, R. A. (1985). The anhedonia hypothesis: Mark III. Behavioral and Brain Sciences, 8, 178-186. [ Links ]
Wise, R. A. & Bozarth, M. A. (1987). A psychomotor stimulant theory of addiction. Psychological Review, 94, 469-492. [ Links ]
Wise, R. A. & Colle, L. (1984). Pimozide attenuates free feeding. “Best scores” analysis reveals a motivational deficit. Psycopharmacology, 84, 446-451. [ Links ]
Wise, R. A. & Raptis, L. (1986). Effects of naloxone and primozide on initiation and maintenance measures of free feeding. Brain Research, 368, 62-68. [ Links ]
Wise, R. A., Spindler, J., deWit, H. & Gerberg, G. J. (1978). Neuroleptic-induced “anhedonia” in rats: Pimozide blocks reward quality of food. Science, 201(4352), 262-264. [ Links ]
Recibido: 28 de agosto de 2007
Revisado: 13 de abril de 2008
Aceptado: 7 de mayo de 2008
*Resultados parciales del presente trabajo se presentaron en la 33.º Conferencia Anual de la Asociación para el Análisis de la Conducta, San Diego California, 2007. El trabajo en general sirvió como tesis para que el primer autor obtuviese el grado de Doctor en Ciencias del Comportamiento: Opción en Neurociencias en la Universidad de Guadalajara- CUCBA, Departamento de Ciencias Ambientales. La tesis fue financiada por el Proyecto Consejo Nacional de Ciencias y Tecnología CONACyT 42050-H otorgado al segundo autor. Se agradece a CONACyT la beca otorgada al primer autor que le permitió cursar los estudios del Doctorado y elaborar la tesis. La preparación del manuscrito en su versión final se hizo durante la estancia de año sabático que el segundo autor realizó en la Universidad de Carolina del Norte, Wilmington.
**Separatas del artículo pueden ser obtenidas por cualquiera de los dos autores: Jorge Balderrama Trápaga, Universidad Veracruzana, Girasoles 90, Flores del Valle, Veracruz, Veracruz, México 91948. Correos electrónicos: batover@yahoo.com; jbalderrama@uv.mx.
***Universidad de Guadalajara-CUCS-Neurociencias, Sierra Mojada 950, Peatonal 3, Edificio N, PB., Guadalajara, Jalisco, México 44340, TEL. 52.33.36176625 Correo electrónico: aparicio@cencar.udg.mx.

