










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.6 n.1 São Paulo jun. 2004
ARTIGOS
Comportamento alimentar de boa constrictor, Epicrates cenchria e Corallus hortulanus (Serpentes: Boidae) em cativeiro
Feeding behavior of Boa constrictor, Epicrates cenchria and Corallus hortulanus (Serpentes: Boidae) in captivity
Rodrigo Roveri ScartozzoniI,*; Flavio de Barros MolinaII,**
I Universidade de São Paulo
II Fundação Parque Zoológico de São Paulo
RESUMO
Descreve-se aqui o comportamento alimentar de Boa constrictor, Epicrates cenchria e Corallus hortulanus, comparando-o com registros discordantes existentes na literatura. O comportamento, descrito a partir dos métodos "todas as ocorrências" e "amostragem de seqüências", foi subdividido em quatro fases: (i) localização, aproximação e bote; (ii) constrição; (iii) inspeção; e (iv) ingestão. Na primeira fase, o dardejar de língua era evidente e os botes desferidos preferencialmente na cabeça dos camundongos. Durante a constrição, não se evidenciou, nas três espécies estudadas, existência de lado dominante do corpo em contato com a presa. Todas as serpentes apresentaram padrão ventral de constrição, sendo um o número de espiras utilizado por B. constrictor e E. cenchria e dois por C. hortulanus. Na fase de inspeção, a serpente tocava a presa com o focinho e dardejava a língua. A ingestão geralmente se iniciou pela cabeça da presa.
Descritores: Comportamento alimentar, Serpentes, Boidae.
ABSTRACT
The feeding behavior of Boa constrictor, Epicrates cenchria, and Corallus hortulanus (Snakes, Boidae) is here described and compared with discrepant data from the literature. Records were obtained according to the "all occurrence" and "sequence sampling" methods. Behavior was divided into four phases: (i) orientation towards prey, approach and strike; (ii) constriction; (iii) inspection; and (iv) swallowing. In the first phase, tongue flicking was evident and the strikes were directed preferentially on the head of mice. During constriction, no side dominance was evident, only a few specimens displayed a preference for the side of the body in contact with the prey. All snakes presented a ventral pattern of constriction and the number of coils utilized was one for B. constrictor and E. cenchria and two for C. hortulanus. The prey was inspected by tongue flicking and snout touching and usually was swallowed headfirst.
Index terms: Feeding behavior, Snakes, Boidae.
As serpentes podem exibir diferentes métodos de detecção, captura, imobilização e ingestão de suas pesas (ver Greene, 1997; Pough et al., 1998; Shine, 1991). Muitas espécies detectam e capturam seu alimento por meio de estímulos químicos (Burghardt, 1970) ou visuais (Czaplicki & Porter, 1974; Lillywhite & Henderson, 1993). Outras, como muitos boídeos e viperídeos, possuem receptores térmicos, sensíveis a raios infravermelhos emitidos pela presa em potencial (Cock Buning, 1983; Stafford & Henderson, 1996; Tolson & Henderson, 1993). Depois de capturada, a presa pode ser morta por envenenamento ou constrição (ou ambos os métodos; Shine & Schwaner, 1985) ou ainda ser ingerida viva (Greene, 1997). As serpentes possuem adaptações cranianas que permitem a ingestão de presas relativamente grandes (Arnold, 1983; Greene, 1983) e os mecanismos utilizados para a ingestão seguem um padrão geral (ver Cundall, 1987).
Diversas serpentes subjugam suas presas por constrição, comportamento que é caracterizado pela imobilização da presa por pressão exercida por um ou mais pontos do corpo da serpente (Greene & Burghardt, 1978). Alguns padrões na formação de espiras durante a constrição foram reconhecidos (Willard, 1977; Greene & Burghardt, 1978). A lateralidade na constrição, que consiste no lado do corpo em contato com a presa, tem recebido atenção especial (Heirich & Klaassen, 1984; Lopes, Rocha & Abe, 1991; Willard, 1977).
As serpentes dos gêneros Boa, Epicrates e Corallus possuem ampla distribuição na América do Sul (Henderson, Micucci, Puorto & Bourgeois, 1995). Estas espécies podem ser terrestres e semi-arborícolas (e. g. Boa e Epicrates) ou arborícolas (e. g. Corallus) e geralmente são ativas à noite (Henderson, 1992). Os itens alimentares consumidos são aves e mamíferos, podendo se alimentar também de lagartos e anfíbios, que geralmente subjugam por constrição (Henderson 1993a, 1993b; Henderson et al., 1995; Stafford & Henderson, 1996; Tolson & Henderson, 1993). O comportamento alimentar de Boa constrictor, Epicrates cenchria e Corallus hortulanus ainda não foi descrito em detalhes. Alguns autores descreveram e analisaram a fase de constrição nas duas primeiras espécies (Heirich & Klaassem, 1985; Lopes et al., 1991; Willard, 1977) e, de forma muito vaga, em Corallus hortulanus (Willard, 1977). Os resultados obtidos mostraram-se discordantes quanto à questão da lateralidade.
Este trabalho visa (a) descrever todas as fases do comportamento alimentar de Boa constrictor, Epicrates cenchria e Corallus hortulanus; b) comparar os dados obtidos sobre a fase de constrição com os registros discordantes existentes na literatura (Heirich & Klaassem, 1985; Lopes et al., 1991; Willard, 1977); (c) analisar, de forma comparativa, as diferentes fases do comportamento nas três espécies estudadas; (d) sugerir alguns aspectos do comportamento alimentar que merecem novas considerações em estudos futuros, inclusive visando o possível aprimoramento do manejo utilizado em cativeiro, como proposto por Murphy e Campbell (1987).
Método
Este trabalho foi realizado na Fundação Parque Zoológico de São Paulo, entre os meses de novembro de 1999 e fevereiro de 2000. Serpentes adultas (n = 18) foram mantidas individualmente em terrários de madeira com visores na parte frontal e superior. A sala onde estavam os terrários com exemplares de E. cenchria (n = 6) e B. constrictor (n = 4) apresentava temperatura entre 24 e 26º C e umidade relativa do ar entre 65 e 70%. Os terrários com exemplares de C. hortulanus (n = 8) foram mantidos em sala com temperatura entre 27 e 29º C e umidade relativa do ar entre 85 e 95%. Quando necessário, a temperatura e umidade eram mantidas artificialmente. As luzes permaneciam acesas das 07:30 as 17:00h. Galhos de árvores foram dispostos dentro dos terrários com indivíduos de C. hortulanus. Um pote com água fresca era mantido em cada terrário. Para a alimentação, foram utilizados camundongos albinos (Mus musculus) pesando entre 35 e 45 gramas, sendo que estes eram oferecidos em intervalos semanais, durante a manhã.
Em face dos hábitos secretivos das serpentes, estudos sobre comportamento alimentar vêm sendo realizados em laboratório já há cerca de quarenta anos (Ford, 1996; Mushinsky, 1987). Esses estudos têm considerado espécies de diferentes grupos taxonômicos ou com variadas características ecológicas e comportamentais, como no caso de espécies que utilizam diferentes ambientes, ou que exibem diferentes estratégias de caça e táticas de subjugação da presa (e. g. Chiszar, Lee, Radcliffe & Smith, 1992; Savitzky, 1992; Willard, 1977). Diversos estudos com espécies rodentívoras têm utilizado camundongos e ratos criados em biotério, sem qualquer prejuízo para os dados coletados (e. g. Chiszar et al., 1992; Kardong, 1993; Lopes et al., 1991).
O comportamento foi descrito baseando-se no método de "todas as ocorrências" e "amostragem de seqüências" (Lehner, 1996). Foram acompanhados, pelo menos, três episódios alimentares para cada serpente, totalizando aproximadamente 22 horas cronometradas de observações. A duração de cada fase do comportamento foi citada sempre em minu-tos:segundos. Os padrões de constrição e o lado do corpo usado em contato com a presa foram anotados (ver Heirich & Klaassem, 1985; Willard, 1977, para detalhes). Uma espira foi considerada completa quando o corpo da serpente envolvia a presa por inteiro. Em todas as alimentações, o observador se manteve a aproximadamente dois metros de distância do terrário.
Diferenças estatisticamente significativas entre os lados usados durante a constrição da presa foram testadas pelo Qui-quadrado, com um grau de liberdade e nível de significância de 1%. Os testes foram feitos dentro de cada espécie, considerando-se o total de episódios alimentares observados: Boa constrictor (n = 24), Epicrates cenchria (n = 26) e Corallus hortulanus (n = 39).
Resultados
A seqüência do comportamento alimentar observada para todas as espécies pode ser dividida em quatro fases: i) localização, aproximação e bote; ii) constrição; iii) inspeção; e iv) ingestão. Tanto as fases, quanto sua duração (tabela I), mostraram-se muito semelhantes para os três gêneros estudados.
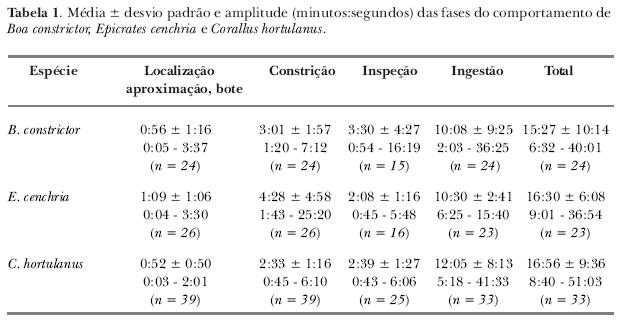
A primeira fase tinha início quando a presa era colocada dentro do terrário, a uma distância aproximada de 40 cm do predador. Neste momento, a serpente iniciava o dardejar de língua e, às vezes, se aproximava lentamente do camundongo utilizando locomoção retilínea. Indivíduos de C. hortulanus geralmente permaneciam sobre os galhos dispostos no terrário e se aproximavam do camundongo, que se encontrava no chão do terrário, pela distensão da parte anterior do corpo. Em alguns casos, a movimentação do camundongo pareceu estimular a aproximação da serpente, porém ela também podia aguardar até que a presa se aproximasse o bastante de seu local de espera.
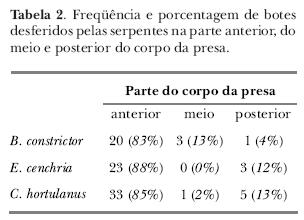
Antes do bote, era evidente o dardejar de língua e a postura da região anterior do corpo retraída em forma de "S". A maioria dos botes foi desferida na região anterior do corpo da presa (tabela II) quando esta se encontrava a uma distância de aproximadamente 10 cm da serpente.
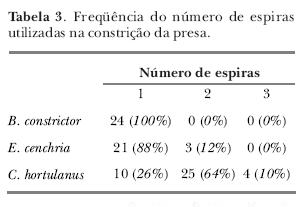
Não foram observadas diferenças estatisticamente significativas entre os lados do corpo utilizados durante a fase de constrição nas espécies analisadas. Somente cinco indivíduos (28%) utilizaram sempre o mesmo lado do corpo, que pode ser o direito ou esquerdo. Todas as serpentes apresentaram padrão ventral de constrição, que consiste na formação de espiras com o ventre voltado para a sua própria cabeça. O número de espiras utilizadas foi, geralmente, de uma para B. constrictor e E. cenchria e duas para C. hortulanus (tabela III).
Um indivíduo de E. cenchria, que imobilizou a presa irregularmente, isto é, com sobreposição na formação de espiras e estas tanto com o lado direito quanto o esquerdo em contato com a presa (ou sobre uma espira anterior), teve o tempo total do comportamento prolongado para 36:54 minutos (tabela I).
Durante a inspeção, a serpente tocava repetidas vezes a presa com o focinho e dardejava muito a língua. O dardejar de língua pareceu aumentar de freqüência comparativamente às fases anteriores da seqüência alimentar. Quando o bote era desferido diretamente na cabeça da presa, esta fase algumas vezes não ocorria e a serpente iniciava a ingestão imediatamente após a constrição. Na maioria das vezes, os indivíduos de C. hortulanus iniciaram a fase de inspeção com a presa ainda fortemente presa entre as espiras de constrição, fato dificilmente observado para as outras espécies, as quais geralmente desfazem as espiras.
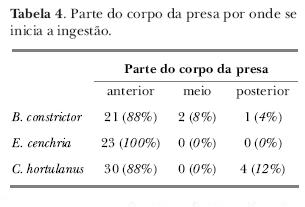
Na fase de ingestão, a presa geralmente era engolida pela parte anterior (tabela IV). Em algumas observações, os camundongos foram ingeridos pelo meio do corpo ou pela região caudal. Nestes casos, em três observações de B. constrictor, o processo foi prolongado para 30:00; 30:15 e 36:25 minutos (tabela I). Em quatro observações com C. hortulanus, o mesmo prolongamento de tempo foi registrado (14:04; 23:03; 38:45 e 41:33 minutos).
Discussão
A detecção da presa pelas espécies analisadas parece ser feita por uma associação entre os estímulos térmicos, visuais e químicos. O dardejar de língua permite à serpente captar informações químicas provenientes tanto do ambiente quanto da presa em potencial (Burghardt, 1970). Porém, em várias observações, os movimentos do camundongo pareceram desencadear o ataque predatório. Em outros boídeos ou mesmo em outros grupos de serpentes (e.g. alguns viperídeos), tanto os estímulos visuais quanto os térmicos são importantes no comportamento que antecede o bote predatório (Cock Buning, 1983; Sazima, 1989). Contudo, estudos mais específicos devem ser feitos para se avaliar, com mais precisão, qual (ou quais) estímulo poderia estar determinando ou ser mais importante nas fases anteriores ao bote.
A tendência de capturar roedores pela parte anterior do corpo, notificada neste estudo, pode ser explicada como um fator de diminuição das possibilidades de retaliação por parte da presa, como demonstrado em estudos com o colubrídeo Elaphe quadrivirgata (Moori, 1991). Bothrops jararaca também desfere a maioria dos botes na região anterior de camundongos (Sazima, 1989). Kardong (1982), em estudo com o elapídeo Naja haje, verificou que esta espécie geralmente captura roedores pela parte posterior do corpo, os quais respondem com mordidas, fazendo com que as serpentes, em muitos casos, percam o interesse pela presa.
O lado do corpo da serpente usado na constrição da presa é motivo de controvérsia. Willard (1977) não encontrou preferência ou dominância no uso do lado esquerdo ou direito, em nível específico ou individual, em qualquer das 95 espécies de serpentes (Colubridae e Boidae) estudadas. Heirich e Klaassem (1985), em estudo com representantes das mesmas famílias, concluíram que, para algumas espécies, existe preferência para um determinado lado na constrição. Para estes autores, Boa constrictor é uma espécie tipicamente destra, o que não pode ser levado em consideração uma vez que as suas observações foram feitas em apenas um indivíduo. Lopes et al. (1991), em estudo com Boa constrictor e Epicrates cenchria, verificaram a preferência de lado somente em nível individual e em baixa freqüência. O presente estudo concorda com os dados encontrados por Lopes et al. (1991) para Boa constrictor e Epicrates cenchria e acrescenta dados relativos a Corallus hortulanus demonstrando, para as três espécies, a inexistência de preferência em nível específico e, em baixa freqüência, a existência de preferência em nível individual.
Heirich e Klaassem (1985) compararam a preferência no uso de um determinado lado na constrição nas serpentes estudadas por eles, com o fato de existirem destros e canhotos em humanos (e.g. Springer & Deutsch, 1981) ou dominância de lado em outros animais (e.g. Marx, 1983). Porém, parece claro não existir um padrão espécie-específico para esta característica em Boa constrictor, Epicrates cenchria e Corallus hortulanus. Em nível individual, a freqüência de serpentes com lado dominante (direito ou esquerdo) foi baixa (Lopes et al., 1991; este estudo), fator que merece novas investigações considerando-se, inclusive, a existência de variações temporais.
Todas as serpentes estudadas apresentaram um padrão ventral de constrição, concordando com os dados de Lopes et al., (1991) e Willard (1977). Na constrição, o número de espiras parece estar relacionado com o tamanho da serpente ou da força aplicada (Canjani, Andrade, Cruz-Neto & Abe, 2003; Lopes et al., 1991). Segundo esses autores, quando a presa oferecia algum tipo de resistência, a serpente imediatamente respondia com uma espira extra ou com o aumento da força das espiras já utilizadas, como também observado para alguns indivíduos analisados neste estudo.
As espécies analisadas provavelmente utilizam estímulos químicos (dardejar de língua) ou tácteis para achar a cabeça da presa, antes de iniciarem a ingestão. Estudos com crotalíneos indicam que a diferenciação entre a cabeça e a parte caudal da presa ocorre, principalmente, por meio de estímulos químicos da presa envenenada (Cock Buning, 1983). Bothrops jararaca, além de dardejar a língua, também toca repetidas vezes com o focinho o corpo da presa envenenada, provavelmente para sentir a direção de seus pelos e localizar a região da cabeça (Sazima, 1989). Greene (1976) sugere que serpentes ofiófagas possam se orientar, na localização da região anterior da presa, através da posição de suas escamas.
O comportamento de inspeção de C. hortulanus ainda com a presa entre as espiras de constrição provavelmente se deve ao fato desta espécie ser arborícola, mostrando-se muito bem adaptada a se alimentar em cima de galhos de árvores. Será interessante, portanto, em estudo futuro, observar o comportamento alimentar da espécie em terrários com e sem galhos, analisando-se, em cada caso, a eficiência do comportamento predatório, com a finalidade básica de definir o melhor ambiente a ser utilizado durante o manejo alimentar.
A ingestão da presa é a última fase do comportamento alimentar. A maioria das serpentes ingere as presas a partir da cabeça (Greene, 1989; Salomão, Santos & Puorto, 1995; Voris & Voris, 1983), assim como as serpentes observadas neste estudo. Desta maneira, seus membros ficam comprimidos paralelamente em relação ao corpo, facilitando sua passagem pela boca e esôfago. A ingestão da presa é feita com movimentos alternados dos lados opostos das maxilas, com movimentos de abdução e adução ("advance phase" e "close phase", conforme Kardong, 1977). Quando, eventualmente, a serpente inicia a deglutição pela parte posterior, este processo se torna muito demorado, podendo ser desvantajoso para a serpente em termos de tempo despendido e gasto extra de energia.
Referências
Arnold, S. J. (1983). Morphology, perfomance and fitness. American Zoologist, 23, 347-361. [ Links ]
Burghardt, G. M. (1970). Chemical perception in reptiles. In: J. W. Johnston, D. G. Moulton, & A. Turk (Eds.). Communication by Chemical signals (pp. 241-308). New York: Appleton-Century-Crofts. [ Links ]
Canjani, C., Andrade, D. V., Cruz-Neto, A. P. & Abe, A. S. (2003). Aerobic metabolism during predation by a boid snake. Comparative Biochemical Physiology, A 133, 487-498. [ Links ]
Chiszar, D., Lee, R. K. K., Radcliffe, C. W. & Smith, H. M. (1992). Searching behaviors by rattlesnakes following predatory strikes. In: J. A. Campbell & E. D. Brodie Jr. (Eds.). Biology of the Pitvipers (pp. 369-382). Tyler: Selva. [ Links ]
Cock Buning, T. C. (1983). Thermal sensitivity as a specialization for prey capture and feeding in snakes. American Zoologist, 23, 363-375. [ Links ]
Cundall, D. (1987). Functional morphology. In: R. A. Seigel, J. T. Collins, & S. S. Novak (Eds.), Snakes: Ecology and Evolutionary Biology (pp. 106-140). New York: MacMillan Publ. Comp. [ Links ]
Czaplicki, J. A., & Porter, R. H. (1974). Visual cues mediating the selection of goldfish (Carassius auratus) by two species of Natrix. Journal of Herpetology, 8, 129-134. [ Links ]
Ford, N. B. (1996). Behavior of garter snakes. In: D. A. Rossman, N. B. Ford & R. A. Seigel (Eds.). The Garter Snakes: Evolution and Ecology (pp. 90-116). Norman: University of Oklahoma Press. [ Links ]
Greene, H. (1976). Scale overlap, a directional sign stimulus for prey ingestion by ophiophagous snakes. Zeitschrift fur Tierpsychologie, 41, 113-120. [ Links ]
Greene, H. W. (1983). Dietary correlates of the origin and radiation of snakes. American Zoologist, 23, 431-441. [ Links ]
Greene, H. (1989). Ecological, evolutionary and conservation implications of feeding biology in Old World cat snakes, genus Boiga (Colubridae). Proc. Calif. Acad. Sci. 46, 193-207. [ Links ]
Greene, H. W. (1997). Snakes: the evolution of mystery in nature. Berkeley: University of California Press. [ Links ]
Greene, H. W., & Burghardt, G. M. (1978). Behavior and Philogeny: constriction in ancient and modern snakes. Science, 200, 74-77. [ Links ]
Henderson, R. W. (1992). Activity patterns, temperature, relationships, and habitat utilization in Corallus enydris (Serpentes Boidae) on Grenada. Caribbean Journal of Science, 28, 229-232. [ Links ]
Henderson, R. W. (1993a). On the diets of some arboreal boids. Herpetological Natural History, 1, 91-96. [ Links ]
Henderson, R. W. (1993b). Foraging and diet in West Indian Corallus enydris (Serpentes: Boidae). Journal of Herpetology, 27, 24-28. [ Links ]
Henderson, R. W., Micucci, T. W. P., Puorto, G., & Bourgeois, R. W. (1995). Ecological correlates and patterns in the distribution of Neotropical boines (Serpentes: Boidae): a preliminary assessment. Herpetological Natural History, 1, 15-27. [ Links ]
Heirich, M. L., & Klaassen, H. E. (1985). Side dominance in constricting snakes. Journal of Herpetology, 19, 531-533. [ Links ]
Kardong, K. V. (1977). Kinesis of the jaw apparatus during swallowing in the cottonmouth snake, Agkistrodon piscivorus. Copeia, 338-348. [ Links ]
Kardong, K. V. (1982). Comparative study of changes in prey capture behavior of the Cottonmouth (Agkistrodon piscivorus) and Egyptian Cobra (Naja haje). Copeia, 337-343. [ Links ]
Kardong, K. V. (1993). The predatory behavior of the northern pacific ratlesnake (Crotalus viridis oreganus): laboratory versus wild mice as prey. Herpetologica, 49, 457-463. [ Links ]
Lehner, P. N. (1996). Handbook of ethological methods (2a ed.). Cambridg University Press, pp. 183-193. [ Links ]
Lillywhite, H. B., & Henderson, R. W. (1993). Behavioral and functional ecology of arboreal snakes. In: R. A. Seigel, & J. T. Collins (Eds.). Snakes: Ecology and Behavior (pp. 1-48). New York: McGraw-Hill. [ Links ]
Lopes, H. R., Rocha, C. F., & Abe, A. S. (1991). Constriction Behavior in Snakes: is there a side dominance? Revista Brasileira de Biologia, 51, 853-856. [ Links ]
Marx, J. L. (1983). The two sides of the brain. Science, 220, 488-490. [ Links ]
Moori, A. (1991). Effects of prey size and type on prey handling behavior in Elaphe quadrivirgata. Journal of Herpetology, 25, 160-166. [ Links ]
Murphy, J. B., & Campbell, J. A. (1987). Captive maintenance. In: R. A. Seigel, J. T. Collins, & S. S. Novak (Eds.). Snakes: Ecology and Evolutionary Biology (pp. 165-181). New York: MacMillan Publ. Comp. [ Links ]
Mushinsky, H. R. (1987). Foraging ecology. In: R. A. Seigel, J. T. Collins, & S. S. Novak (Eds.). Snakes: Ecology and Evolutionary Biology (pp. 302-334). New York: MacMillan Publ. Comp. [ Links ]
Pough, F. H., Andrews, R. M., Cadle, J. E., Crump, M. L., Savitzky, A. H., & Wells, K. D. (1998). Herpetology. Prentice Hall, Upper Saddle River, NJ. [ Links ]
Salomão, M. G., Santos, S. M. A., & Puorto, G. (1995) Activity pattern of Crotalus durissus (Viperidae, Crotalinae): feeding, reproduction and snakebite. Studies on Neotropical Fauna and Environment, 30, 101-106. [ Links ]
Savitzky, B. A. C. (1992). Laboratory studies on piscivory in an opportunistic pitviper, the cottonmouth, Agkistrodon piscivorus. In: J. A. Campbell & E. D. Brodie Jr. (Eds.). Biology of the Pitvipers (pp. 347-368). Tyler: Selva. [ Links ]
Sazima, I. (1989). Comportamento alimentar da jararaca, Bothrops jararaca: encontros provocados na natureza. Ciência e Cultura, 41, 500-505. [ Links ]
Shine, R. (1991). Australian Snakes: a natural history. Ithaca: Cornell University Press. [ Links ]
Shine, R., & Schwaner, T. (1985). Prey constriction by venomous snakes: a review and new data on Australian species. Copeia, 1067-1071. [ Links ]
Springer, S. P., & Deutsch, G. (1981). Left brain right brain. San Francisco: W. H. Freeman and Company. pp. 243. [ Links ]
Stafford, P. J., & Henderson, R. W. (1996). Kaleidoscopic Tree Boas: the genus Corallus of Tropical America. Malabar, Florida: Krieger. [ Links ]
Tolson, P. J., & Henderson, R. W. (1993). The natural history of West Indian Boas. Tauton, England: RA Publishing. [ Links ]
Voris, H. K., & Voris, H. H. (1983). Feeding strategies in marine snakes: an analysis of evolutionary, morphological, behavioral and ecological relationships. American Zoologist, 23, 411-425. [ Links ]
Willard, D. E. (1977). Constricting methods in snakes. Copeia 1977, 379-382. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Rodrigo Roveri Scartozzoni
Laboratório de Herpetologia, Instituto Butantan
Av. Vital Brazil, 1500, Butantã
05503-900, São Paulo, SP, Brasil
E-mail: rodrigobuta@hotmail.com
Recebido em 26 de janeiro de 2004
Revisão recebida em 18 de maio de 2004
Aceito em 27de julho de 2004
* Depto de Ecologia, Instituto de Biociências, USP, São Paulo, Brasil.
** Setor de Répteis, Fundação Parque Zoológico de São Paulo. E-mail: fbmolina@uol.com.br
Agradecemos a Otavio A. V. Marques e Maria da Graça Salomão, do Laboratório de Herpetologia do Instituto Butantan, pelas críticas e sugestões feitas ao manuscrito e a Edvaldo Antonio dos Santos, do Zôológico de São Paulo, pelo apoio na manutenção das serpentes.

