










Serviços Personalizados
Journal

artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTIIndicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805
Rev. etol. vol.11 no.1 São Paulo 2012
The whistles of the guinea pig: an evo-devo proposal
Os assobios do porquinho-da-índia: uma proposta ontogenetico-evolutiva
Clara CoratI; Rita Carolina Rodrigues Branco TaralloI; Carine SavalliII; Rosana Suemi TokumaruIII; Patrícia Ferreira MonticelliI,†; César AdesI, ‡
IUniversidade de São Paulo
IIUniversidade Federal de São Paulo
IIIUniversidade Federal do Espirito Santo
ABSTRACT
When separated from their mother, wild and domestic cavy pups emit high-pitched whistles. Whistles are also emitted only by the domestic species in response to food-associated stimuli. We compared isolation whistles (IS) emitted by guinea pig pups separated from their mothers to the food-anticipation whistles (FA) emitted by the same adult individuals in response to a feeding routine. Results revealed no significant differences in the structure of the IS and FA whistles, but showed ontogenetic changes along the period. Results are discussed both in relation to the physiological mechanisms controlling whistle vocalizations and their evolutionary origin in the cavies repertoire.
Keywords: isolation call, whistle, domestication, food-associated call, parental behavior, bioacoustics
RESUMO
Quando separados de suas mães, filhotes de preás selvagens e domésticos emitem assobios agudos. Assobios também são emitidos apenas pela espécie doméstica em resposta a estímulos associados à presença de alimentos. Nós comparamos o assobio de isolamento (IS) emitido por filhotes separados de suas mães aos assobios para o tratador (TR) emitidos pelos mesmos indivíduos adultos em resposta a uma rotina de alimentação. Os resultados não revelaram diferenças na estrutura dos assobios IS e TR, mas mostraram mudança ontogenética ao longo do período. Os resultados são discutidos em relação aos mecanismos fisiológicos que controlam a emissão dos assobios e a origem evolutiva desta vocalização no repertório dos preás.
Palavras-chave: chamado de separação, assobio, domesticação, chamado associado ao alimento, comportamento parental, bioacústica
One of the interesting aspects about domestication is the establishment of changes in the species social behavior that usually involves an increase in tolerance to intraspecific and interspecific contact. This tolerance involves both non-vocal and vocal social behavior (Trut, Plyusnina & Oskina, 2004). Guinea pigs - domesticated about 7,500 years ago in the Andean region (Wing, 1986; Lavallée, 1990) from the same ancestral species of C. aperea and C. tschudii (Bonatto, Schneider & Lamb, 1995; Spotorno, Valladares, Marín et al., 2004) - , besides presenting structural changes in their acoustic repertoire in relation to C. aperea (Monticelli & Ades, 2013), also present a specialized use of a whistle emitted in response to stimuli produced by the caretaker at the time of feeding. What is the evolutionary origin of this signal?
In both species, C. porcellus and C. aperea, as in most mammals, mothers are the main caregivers, but allosuckling also occurs among females from the same social group (Takamatsu, Tokumaru & Ades, 2003). Males are very tolerant towards pups and eventually display frisk hops in interaction with them, but do not provide direct care (Beisiegel, 1993; Adrian, Brockmann, Hohoff et al., 2005). King (1956), observing guinea pigs in a "semi-natural" environment, reported that the precocious pups were born in natural shelters on the ground. They remained there, hidden, until three or four days old. When they left, they preferentially followed their mothers. Pups require care only for their first four weeks of life (Rood, 1972). During this period, when visually separated from their mothers they readily emit whistles (Berryman, 1976; Monticelli & Ades, 2013), especially in the first two weeks of life and progressively less until the fourth (Pettijohn, 1979a).
The isolation-whistle (described by Berryman, 1976, as whistle) is emitted by other species of Cavia (C. intermedia and C. magna; unpublished data). In guinea pigs, isolation-whistles present individual characteristics (Tokumaru, Ades & Monticelli, 2004) and can be used by mothers to recognize pups, especially in large clutches (Kober, Trillmich & Naguib, 2008). This signal varies throughout development (Monticelli, Ades, Tokumaru et al., 2003; Monticelli , Tarallo & Ades, 2009) and along a 15 min isolation period, which indicates variation in the motivational state of the pups (Monticelli, Tokumaru & Ades, 2004). Besides the variation in the acoustic structure over time described in the papers cited above, the whistles also present a decrease (Pettijohn, 1979a) in the emission rate. These changes are accompanied by increases in plasma cortisol levels and immobility during a 30 min isolation period (Ritchey & Hennessy 1987; Hennessy 1988; Hennessy & Sharp 1990; Sachser, 1998; Hennessy, Deak & Schmil-Webb, 2001).
The food-anticipation whistle - in Portuguese called assobio-ao-tratador (Ades, Tokumaru & Beisiegel, 1994) - is auditorily very similar to the isolation-whistle. It is emitted only by the domestic species in response to the arrival of the caretaker and other stimuli related to food delivery. This vocalization differs from those emitted by other species during foraging. The food-associated vocalizations are emitted by foragers that first encounter a food item and can serve the function of attracting conspecifics to join the caller. Although the caller has some cost in sharing the food, a number of benefits have been demonstrated for the caller in different species (Cly, Smith & Blumstein, 2012). These vocalizations differ from the food-anticipation whistles of guinea pigs because these are emitted before food is encountered; it is emitted in the presence of a stimulus that precedes food. Clay et al. (2012) argue that although some authors claimed a referential function to the food-associated vocalizations due to the variation in call rate in the presence of different food items, this variation can also reflect the caller´s internal state.
Berryman (1976) does not distinguish between the isolation and the food-anticipation whistles. Coulon (1982) places them into two distinct categories - cri de quetë (isolation-whistle) and sifflement d'appel (food-anticipation whistle) - and points out differences in the rhythm of these whistles. According to Coulon (1982), although in different categories, both vocalizations share the same functional class: both serve the function of attracting attention, first of conspecifics and later of the human caretaker. The first response would be reinforced by the mother (or other conspecifics) - although pups emit this vocalization without training when isolated for the first time - and second, by the appearance of the food. Arvola (1974) also differentiated between the two whistles. While the food-anticipation whistle, which he called shrieks of hunger and thirst, was described as an oral emission (emitted with the mouth wide opened), the isolation-whistle, which he called agitating squeal, was described as a naso-oral emission.
The comparison of the two whistles in the literature is, however, restricted to the presentation and description of typical sonograms obtained from recorders and sonographic analyses softwares very different from what we have today (Arvola, 1974; Berryman, 1976; Coulon, 1982). There are insufficient systematic observations about the structure and variability of the acoustic signals that could clarify the relationship between the two whistles: are they really different in structure or is the food-anticipation whistle the result of ontogenetic changes in the isolation-whistle?
Our goal in this paper is to investigate these questions by comparing the food-anticipation whistles to the isolation-whistles emitted by the same guinea pig individuals. The results will be discussed in relation to the question about the origins of the food-anticipation whistle in the guinea pigs' communication repertoire.
Material and Methods
Study area and subjects
In this study we used one randomly selected individual from each of 8 different litters of 3 to 5 pups obtained from commercial or personal breedings (4 females and 4 males). The study was composed of two different phases. On Phase 1 the subjects were maintained in the animal husbandry facility of the Psychology Institute, University of São Paulo, São Paulo, Brazil, with their native colonies until they were 30 days old. On Phase 2 the subjects were adopted and kept as pet by two experimenters (RCRBT e CC). There was always only one animal per time kept as pet on phase two (each new subject was adopted just when data with the previous one had been concluded). In both phases the subjects were kept in 60x60x100 cm plastic or polypropylene cages, received water and guinea pig or rabbit chow ad libitum and fresh vegetables once a day, except on the weekends.
Data Collection
Phase 1 - The isolation-whistle (IS) recording: We performed 3 recording sessions of the isolation-whistle for each pup, around days 7, 14 and 21 of their lives. On the sessions we transported the pups individually to a test room and placed them in a wooden box (50x50x30 cm) for 5 minutes. No food or water was available. After that the animals returned to their living cages.
Phase 2 - The food-anticipation whistle (FA) recording: The subjects were taken to the researcher's homes when they were 30 days old, and they started living as pets, interacting and submitted to the family routine. Every day, a food supplement, preferred by the guinea pigs (fruits, carrots or cabbage) was offered, always at the same time and on the same routine, following Ades, Beisiegel and Tokumaru (1992): the experimenter went to the refrigerator, opened the door, shook a plastic bag producing a characteristic noise, got the food from the bag, closed the door, approached the subject and put the food in its living box. After subjects started whistling to the feeding routine, a recording day was scheduled. The equipment was prepared before the experimenter started the routine and was turned off about 5 minutes after the food delivery All vocalizations emitted during the routine were recorded.
Equipment
Sound recordings were made using a Sony professional portable digital audio tape recorder (DAT - TCD-D8; sampling rate: 48 kHz, frequency response: 20 - 24,000 Hz ± 1.0 dB; dynamic range >87 dB) and a hypercardioid directional microphone supported by a tripod and situated 30 cm above the floor (Sennheiser ME 67 with K6 powering module and bass roll-off, with the filter switch on).
Sound Analysis
All of the acoustic signals produced were digitized with 8-bit accuracy from DAT to a personal computer using Raven 1.4 (Cornell Lab of Ornithology: http://www.birds.cornell.edu/brp/raven/AwardsProgram.html) via a Sound Blaster Audigy A/D converter at a sampling rate of 48 kHz. Spectrograms were generated and analyzed using the following settings: Hamming window; 1,024 point fast Fourier transform (FFT); 100% screen size; 93.75% overlap; 47Hz frequency resolution; 0 to 24Hz frequency band; 1.33ms time resolution.
We defined as measurable notes of whistle (IS and FA) only those that had at list three harmonics and the three parts - A, B and C described in Tokumaru, Monticelli & Ades (2004). We selected the first 10 consecutive notes of each recording period (IS7, IS14, IS21 and FA) to be measured.
The acoustic parameters measured in all the calls were: the number of visible harmonics on initial part of the note - A, following Tokumaru et al. (2004) - , low frequency, higher frequency, dominant frequency, note duration, inter-note interval, rhythm (number of notes/second) and average power (the value of the spectrogram's power spectral density, as defined by Raven software). We used the first harmonic of each element of each note to delimit and measure the parameters.
Data Analysis
We first described the acoustic parameters of the whistles in the four recording periods (IS7, IS14, IS21 and FA) using average (standard deviation), minimum and maximum values. Previously to all analyses, the 10 notes of each individual and each recording period were summarized by the mean.
To all statistical analyses, we considered the data from five individuals. Three of the individuals could not be included due to different reasons (a female got sick when it was 14 days old; the recordings of one female were damaged and one male never emitted FA on phase 2).
To compare isolation (IS) and food-anticipation (FA) whistles, we summarized the three first recording periods IS7, IS14 and IS21 by the means and compared the data to those from the recording period FA using the Wilcoxon signed rank test (Hollander & Wolfe, 1998). The use of a non-parametrical test was due to the low number of subjects.
We also evaluated whether there were tendencies for each acoustic parameter across the recording periods indexed by time using the longitudinal growth curve data methodology (Winer, 1971; Singer & Andrade, 1986). This technique allows incorporating the statistical dependence between observations in the same individual in the estimation of the tendency. We used the compound symmetry structure for the variance-covariance matrix that assumes constant correlation between any two measures of the same animal in a line regression model. The assumption of normality was evaluated on the residuals of the model by Shapiro-Wilk and Kolmogorov-Smirnov. Only variables for which the normality assumption was satisfied were analyzed using this methodology. For the other variables, only a descriptive analysis of the individual profiles was considered.
Results
In the isolation context, all pups presented vocalizations in the first recording period, at 7 days old and one pup had not whistled when it was 21 days old. The analyses of the recording periods in this context (and in phase 2) revealed the presence of the five categories of vocalizations described by Berryman (1976) in the same context, occurring concomitantly to the whistles: chut and chutter - described as contact calls by Monticelli & Ades (2013) - , whine, low whistle (less than three harmonics) and the tweet.
Pups started to emit the food-anticipation whistle when they were still living in the colony, at about 14 days of life, in response to the feeding routine. After being taken home as pets, the individuals started responding to the feeding routine, in average, after 10 days in the new environment.
We noticed a positive relation between the whistle emission rate and the proximity of the food delivery: the rate of vocalization was accelerated as the experimenter approached the animal.
Comparison between isolation and food-anticipation whistles
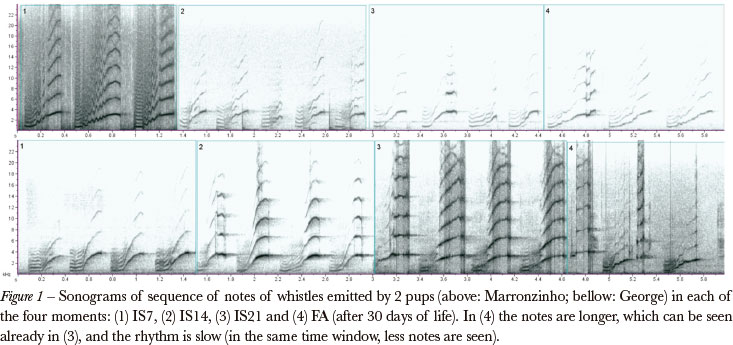
The modulated frequency of the whistle notes allow us to divide them in three parts, as described in Tokumaru et al. (2004). Element A presents a smooth ascending modulation (0.36 kHz in average), element B presents a marked ascending modulation (from 1.03 kHz to 3.42 kHz, in average) and element C presents a varied modulation, generally descendant (until 2.64 kHz, in average). Figure 1 presents sonograms of whistles emitted in the isolation and food-anticipation contexts. Although some variation is visually apparent, no significant differences were obtained from the Wilcoxon signed rank test (Table 1).
Ontogenetic change in the whistle
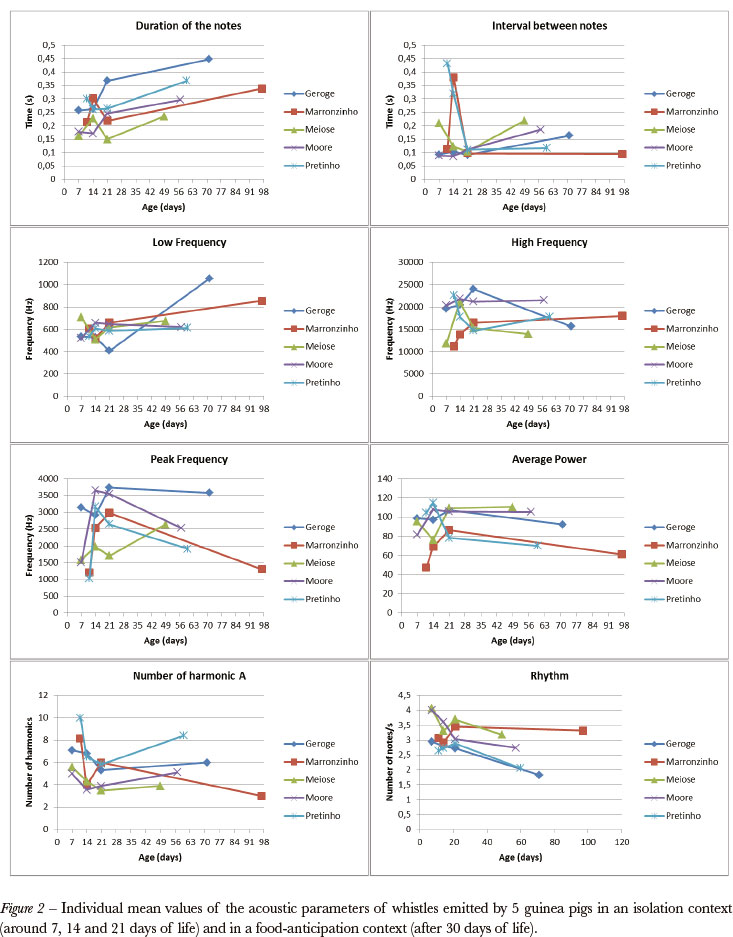
Considering the absence of differences between the isolation and the food-anticipation whistles revealed by the previous analyses, we decided to look for changes over time considering the dependency between observations in the same individual. Table 2 shows the results of the descriptive statistics of the parameters of the whistles emitted by our subjects along the recording periods.
The parameters for which the normality assumption was satisfied were evaluated in search of tendencies over time. From the results displayed in Figure 2 and Table 2, it can be observed that whistles presented a significant tendency of increase in duration and low frequency and of decrease in rhythm.
Discussion
Our results indicate that there are no statistical differences between the structure of the isolation and the food-anticipation whistles. The differences noted by other authors (Arvola, 1974; Coulon, 1982) were probably related to their methods of analysis based on the description of typical sonograms. On the other hand, our results also show that the basic structure of the whistles changes ontogenetically. As guinea pigs grew, whistle notes became longer and presented higher low frequency, while the rhythm of whistles emission became slower.
These changes are consistent with anatomical changes in the vocal tract that occur as the animal grows: the increase in the larynx length and the thickening of the vocal cords produce narrower band calls (a significantly rise in the low frequency and a tendency of fall in the maximum and peak frequencies). The increase in lung capacity would facilitate the emission of longer, more intense notes that are consequently produced in a slower rhythm. Our results are similar to those displayed in the literature for other species - squirrel monkeys in Winter, Handley, Ploog et al. (1973); Lieblich, Symmes, Newman et al. (1980); vervet monkeys in Seyfarth & Cheney (1986); pigmy marmoset in Elowson, Snowdon & Sweet (1992) - that describe variations in the duration and frequency of the calls along development.
Together, these results lead us to conclude that the isolation-whistle is the same as the food-anticipation whistle; both present the same basic structure. A question that follows from this conclusion is the occurrence of this vocalization in two different contexts. During separation from conspecifics only pups emit whistles, and in response to food cues both pups and adults emit whistles. What is the mechanism modulating the emission of this vocalization?
A possible mechanism can be related to the functioning of the pituitary - adrenocortical (PAC) system, responsible for stress responses. It is well known that, during a 30 min isolation period, as the rate of guinea pig pup´s whistles emission decrease, cortisol levels and immobility increase (Ritchey & Hennessy, 1987; Hennessy, 1988; Hennessy & Sharp, 1990; Sachser et al., 1998; Hennessy et al., 2001). The increase in the cortisol level seems to be a response to the separation of the mother and from the surrogate environment indicating that this is a stressful event to the pup (Wewers, Kaiser & Sachser, 2003). In species that form attachment bonds, the increase in plasma concentrations of PAC hormones has been demonstrated in response to the separation from the attachment object (Henessy, Maken & Graves, 2000). But while cortisol level presents a linear increase in response to time in isolation, the emission rate of whistles present a decrease (Pettijohn, 1979a; McInturf & Henessy, 1996). This relation suggests that low levels of stress trigger the emission of whistles, but, as levels increase, vocalization and locomotion decrease. Our proposal is supported by the results of some studies that obtained simultaneously decreases in whistles emission and increases in cortisol levels (Henessy, Ptstick, O'Leary, Maken & Farré, 2001) or the inverse relation (Henessy, Maken & Graves, 2000; Henessy, O'Leary, Hawke & Wilson, 2002; Wewers, Kaiser & Sachser, 2003).
Our suggestion is that mild stress is the physiological trigger for the emission of whistles vocalization and this kind of stress occurs both during brief isolation periods and upon the perception of stimulus related to food delivery (Mistlberger, 1993; Ott, Friedrich, Prilop, Lehnert, Chara, Born & Hallschmid, 2011). We propose that the triggering of whistles by mild stress and its suppression by higher levels of stress was possibly selected in the context of naturalistic mother-pup separations that can occur during foraging. As already demonstrated (Pettijohn, 1979; Berryman, 1981; Coulon, 1973; Tokumaru, Ades & Monticelli, 2006) isolation-whistles function as signals which elicit approach of the mother. Elsewhere (Monticelli, Tokumaru & Ades, 2004), we argued that immediately after separation in a naturalistic foraging context, as the mother is likely to be not very far, it may be advantageous for the pup to keep moving and to emit fast repetitive sequences of whistle notes, maximizing the probability of the vocalization being heard and responded to. As time passes in isolation, probability of recovery by the mother decreases and less conspicuous vocal signals and immobility may represent the best tradeoff between calling and defense against predators. Considering that this hypothesis is correct, we further propose that this mechanism could be activated in the guinea pig under laboratory conditions in the food-anticipation context because it mimicries the mild stress condition produced during brief isolation periods.
This explanation can also be used to understand why both wild and domesticated species of cavies emit the isolation-whistles but only the domesticated guinea pigs emit the food-anticipation whistle. In comparison to at least on wild cavy species (Cavia aperea), guinea pigs showed decreased reactivity of PAC system (Künz & Sachser, 1999). This difference is robust as it was seen even in the comparison between guinea pigs and wild cavies reared in captivity for 30 generations (Künzl, Kaiser, Meier, & Sachser, 2003). The authors argue that the decreased reactivity of the guinea pigs` stress axes is one of the results of domestication that helped these animals to adjust to man-made housing conditions. Reinterpreting these results, we could say that guinea pigs are less responsive to environmental changes or that they have a higher threshold for the activation of the stress axes in response to these changes.
This effect of domestication would give the guinea pigs the opportunity to answer to food-anticipation stimuli with whistle calls because, in comparison to the wild cavies, the human keeper does not enhance their levels of stress. It is probable that both guinea pigs and wild cavies learn to expect food when food-associated stimuli are present. Personal (PFM) observations in a wild cavy colony have shown that animals approach the border of the cage, sniff the air and look in the direction of the keeper when he/she starts the feeding routine. But although both species show food-anticipation behavior and, probably, the corresponding changes in physiology, wild cavies probably present an extra level of stress as food stimuli include the presence of a human keeper. This higher level of stress in response to food-anticipation stimuli would prevent the emission of whistles by wild cavies. If this hypothesis is correct, we could expect that wild cavies treated with antidepressants during exposition to food-anticipation stimuli would emit whistles after a period of habituation. We do intend to perform this experiment to test this hypothesis.
In the light of our results and the proposed mechanism controlling whistles emission, we suggest a change in the form of reference to the whistles vocalizations. Researchers have categorized the whistles referring to the context in which they occur. But, as they are structurally the same between contexts, we suggest that the categorization is made in reference to the mechanism triggering the whistles, i.e., anxiety whistles or mild distress whistles. Of course, this suggestion is subjected to the confirmation of the hypothesis described above.
A limitation of this study is that we were not able to collect isolation and food-anticipation whistles from the same individual at all developmental stages. Guinea pig pups do emit isolation whistles and food-anticipation whistles. But while the first ones are easily individually collected, the second occurs when pups are responding to the feeding routine in the colony. We did not try to adopt individuals as pups, because it would imply bottle-feeding them, increasing mortality chances, but it is possible that adopted pups would emit whistles to the feeding routine while in the familiar cage and in response to isolation in an unfamiliar environment. Another possibility is the use of individual microphones permanently attached to the animals to record the whistles emitted in the colony. Both suggestions can be subjected to future testing depending on technical adjustments in our lab.
Although we did not compare isolation and food-anticipation whistles at the same developmental stages, the comparison of adult food-anticipation whistles and pups isolation whistles showed both the absence of structural differences and ontogenetic change in the basic structure. The evo-devo hypothesis offered here to their emission, by guinea pigs in different contexts and by wild cavies only in the isolation context, is supported by these results and others in the literature and offers the possibility of future testing and demonstration.
Ethical note
This work followed the ''Principles of laboratory animal care'' (NIH publication No. 86-23, revised 1985) and was approved by the Committee of Ethics for Animal Use (CEPA) from Instituto de Psicologia [Psychology Institute] of the University of São Paulo (proc. 005.2009).
Acknowledgments
CNPq. Processo 479939/2007-1.
References
Ades, C., Tokumaru, R. S. and Beisiegel, B. M. (1994). Vocalizações antecipatórias da cobaia Cavia porcellus em situação de alimentação. Biotemas 7(1&2): 79-93. [ Links ]
Adrian, O., Brockmann, I., Hohoff, C. and Sachser, N. (2005). Paternal behaviour in wild guinea pigs: a comparative study in three closely related species with different social and mating systems. Journal of Zoology of London 265: 97-105. [ Links ]
Arvola, A. (1974). Vocalizations in the guinea pig Cavia porcellus L. Annales Zoologic Fennici 11(1-96). [ Links ]
Beisiegel, B. M. (1993). Dinâmica familiar e comportamento paterno na cobaia doméstica. Departamento de Psicologia Experimental. São Paulo, Universidade de São Paulo. [ Links ]
Berryman, J. C. (1976). Guinea pig vocalizations, their structure, causation and function. Zeitschrift für Tierpsychologie 41: 80-106. [ Links ]
Berryman, J.C. (1981). Guinea pig responses to conspecific vocalizations: playback experiments. Behavioral and Neural Biology, 31, 476-482. [ Links ]
Bonatto, S. L., Schneider, M. P. and Cordeiro, A. R. (1995). Evolution of isoesterase tissue expression patterns in Cavia (Caviidae, Rodentia). Brazilian Journal of Genetics 18(3): 391-396. [ Links ]
Clay, Z., Smith, C. L. and Blumstein, D. T. (2012). Food-associated vocalizations in mammals and birds: what do these calls really mean? Animal Behaviour, 83(2), 323-330 [ Links ]
Coulon, J. (1973). Le répertoire sonore du cobaye domestique et as signification comportamentale. Revue du Comportment Animal, 7, 121-132. [ Links ]
Coulon, J. (1982). La communication acoustique du cobaye domestique comparaison avec quelques rongeurs. Journal de Psychologie 1: 55-78. [ Links ]
Elowson, A. M., Snowdon, C. T. and Sweet, C. J. (1992). Ontogeny of trill and J-call vocalization in the pygmy marmoset, Cebuella pymaea. Animal Behaviour 43: 703-715. [ Links ]
Hennessy, M. B. (1988). Both prevention of physical contact and removal of distal cues mediate cortisol and vocalization responses of Guinea Pig pups to maternal separation in a novel environment. Physiology and Behavior, 43: 729-733. [ Links ]
Hennessy, M. B.; Deak, T. and Schmil-Webb, A. (2001). Stress-induced sickness behaviors: an alternative hypothesis for responses during maternal separation. Developmental Psychobiology, 39: 76-83. [ Links ]
Hennessy, M.B., Maken, D.S. and Graves, F.C. (2000). Consequences of the presence of the mother or unfamiliar adult female on cortisol, ACTH, testosterone and behavioral responses of periadolescent guinea pigs during exposure to novelty. Psychoneuroendocrinology 256, 619-632. [ Links ]
Hennessy, M. B.; O'Leary, S. K.; Hawke, J. L. and Wilson, S. E. (2002). Social influences on cortisol and behavioral responses of preweaning, periadolescent, and adult guinea pigs. Physiology & Behavior, 76: 305-314. [ Links ]
Hennessy, M. B.; Pitstick, L.; O'Leary, S.; Maken, D. S. and Farré, A. J. (2001). Effects of a novel potential antidepressant on the behavior and cortisol levels of isolated guinea pig pups. Pharmacology, Biochemistry and Behavior, 69: 527-533. [ Links ]
Hennessy, M. B. and Sharp, K. (1990). Voluntary and involuntary maternal separation in Guinea Pig pups with mothers required to forage. Developmental Psychobiology, 23: 783-796. [ Links ]
King, J. A. (1956). Social relations of the domestic guinea-pigs living under semi-natural conditions. Ecology 37: 221-228. [ Links ]
Kober, M., Trillmich, F. and Naguib, M. (2008). Vocal mother-offspring communication in guinea pigs: females adjust maternal responsiveness to litter size. Frontiers in Zoology 5(1): 13. [ Links ]
Künzl, C. and Sachser, N. (1999). The endocrinology of domestication: a comparison between the domestic guinea-pig (Cavia aperea f. porcellus) and its wild ancestor, the cavy (Cavia aperea). Hormones and Behavior 35: 28-37. [ Links ]
Künzl, C.; Kaiser, S.; Meier, E. and Sachser, N. (2003). Is a wild mammal kept and reared in captivity still a wild animal? Hormones and Behavior, 43: 187-196. [ Links ]
Lavallée, D. (1990). La domestication animale en Amerique du Sud - Le point des connaissances. Bull. Inst. Fr. Étude Andines 19(1): 25-44. [ Links ]
Lieblich, A. K., Symmes, D., Newman, J. D. and Shapiro, M. (1980). Development of isolation peep in laboratory-breed squirel monkeys. Animal Behavior 28: 1-9. [ Links ]
McInturf, S. M. and Hennessy, M. B. (1996). Peripheral administration of a corticotropin-releasing factor antagonist increases the vocalizing and locomotor activity of isolated guinea-pig pups. Physiology & Behavior, 60(3): 707-710. [ Links ]
Mistlberger, R. E. (1994). Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neuroscience & Biobehavioral Reviews, 18(2), 171-195. [ Links ]
Monticelli, P. F. and Ades, C. (2013). The Rich Acoustic Repertoire of a Precocious Rodent, The Wild Cavy Cavia aperea. Bioacoustics 22(1): 49-66. [ Links ]
Monticelli, P. F., Ades, C., Tokumaru, R. S. and Constantinov, L. (2003). Motivational and ontogenetical variability in guinea-pig pups distress whistles. Annals of the XIX International Bioacoustics Congress, 78-79, Belém, Pará [ Links ].
Monticelli, P. F., Tarallo, R. C. R. B. and Ades, C. (2009). Is food-anticipation whistle of domestic guinea pigs derived from isolation whistle? Bioacoustics, 21(1): 48. [ Links ]
Monticelli, P. F., Tokumaru, R. S. and Ades, C. (2004). Isolation induced changes in Guinea pig Cavia porcellus pup distress whistles. Anais da Academia Brasileira de Ciências, 76(2): 368-372. [ Links ]
Ott, V., Friedrich, M., Prilop, S., Lehnert, H., Jauch-Chara, K., Born, J., et al. (2011). Food anticipation and subsequent food withdrawal increase serum cortisol in healthy men. Physiology & Behavior, 103(5), 594-599. [ Links ]
Pettijohn, T. F. (1979a). Attachment and separation distress in the infant guinea pigs. Developmental Psychobiology 12(1): 73-81. [ Links ]
Ritchey, R. L. and Hennessy, M. B. (1987). Cortisol and behavioral responses to separation in mother and infant Guinea Pigs. Behavioral and Neural Biology, 48: 1-12. [ Links ]
Rood, J. P. (1972). Ecological and behavioural comparison of three genera of argentine cavies. Animal Behavior Monographs 5: 1-83. [ Links ]
Sachser, N. (1998). Of domestic and wild guinea-pigs: studies in sociophysiology, domestication and social evolution. Naturwissenschaften Review Articles 85: 307-317. [ Links ]
Seyfarth, R. M. and Cheney, D. L. (1986). Vocal development in vervet monkeys. Animal Behaviour 34: 1640-1658. [ Links ]
Singer, J. M. e Andrade, D. F. (1986). Análise de dados longitudinais. VII SINAPE: Campinas. [ Links ]
Spotorno, Á. E., Valladares, J. P., Marín, J. C. and Zeballos, H. (2004). Molecular diversity among domestic guinea-pigs (Cavia porcellus) and their close phylogenetic relationship with the Andean wild species Cavia tschudii. Revista Chilena de Historia Natural 77(2): 243-250. [ Links ]
Takamatsu, A. T., Tokumaru, R. S. and Ades, C. (2003). Allosuckling in guinea pigs (Cavia porcellus). Revista de Etologia 5 (Suplemento): 203. [ Links ]
Tokumaru, R. S., Ades, C. and Monticelli, P. F. (2004). Individual differences in infant guinea pig pups isolation whistles. Bioacoustics, 14 (3): 197-278. [ Links ]
Tokumaru, R. S., Ades, C. and Monticelli, P. F. (2006). Can guinea pig mothers learn to discriminate the whistles of individual pups? Revista Brasileira de Zoociências, 8(1): 7-16. [ Links ]
Tokumaru, R. S., Monticelli, P. F. and Ades, C. (1996). Reação de cobaias (Cavia porcellus) a um sinal de alimento. Anais de Etologia 14: 347. [ Links ]
Trut, L. N., Plyusnina, I. Z. and Oskina, I. N. (2004). An Experiment on Fox Domestication and Debatable Issues of Evolution of the Dog. Russian Journal of Genetics 40(6): 644-655. [ Links ]
Wewers, D.; Kaiser, S. and Sachser, N. (2003). Maternal separation in guinea-pigs: a study in behavioral endocrinology. Ethology, 109: 443-453. [ Links ]
Winer, B.J. (1971) Statistical Principles in Experimental Design, 2nd edition, New York: McGraw-Hill Book Co. [ Links ]
Wing, E. S. (1986). Domestication of Andean mammals. High Altitude Tropical Biogeography. F. Vuilleumier and M. Monasterio. Oxford. [ Links ]
Winter, P., Handley, P., Ploog, D. and Schott, D. (1973). Ontogeny of squirrel monkey calls under normal conditions and under acoustic isolation. Behaviour 47: 230-239. [ Links ]
Received in March 28, 2013
Accepted in April 29, 2013
† Corresponding author: Patrícia F. Monticelli University of São Paulo. Av. Bandeirantes, 3900, Psicologia, Anexo do Bloco 3, sala 11 CEP: 14.040-901, Bairro Monte Alegre, Ribeirão Preto, SP. E-mail: pmonticelli@ffclrp.usp.br
‡ In Memoriam. Department of Experimental Psychology, Institute of Psychology, University of São Paulo.
1. This work is the result of the involvement of several people, all supervised by Prof. César Ades. This involvement reflects some questions asked by Prof. César to many of us, his students: what is the whistle that occurs only in the repertoire of the guinea pig, a domesticated species, in response to its human caretaker? Where does it come from? Is it really different from other whistles used among conspecifics? These questions were asked for the first time several years ago, about fifteen, but it took all this time and several people interested in taking a guinea pig home and keeping it as a pet to answer them. Why did we have to take the experimental animals home? Because these vocalizations occur only when the animals are habituated to a feeding routine. This response also occurs under laboratory conditions under a feeding routine. But there the animals are kept in groups, what prevents the registration of individual vocalizations. This technical difficulty has delayed our possibilities of answering those questions for a long time. Now we think we have solved them, at least partially. Unfortunately we did not have the chance to further discuss the answers we present here with our Dear Master. But this paper is dedicated to him as well as our respect. We also attribute to him our involvement in the search of answers about animal behavior.

{kind=link}
{kind=link}
{kind=link}
{kind=link}