










Serviços Personalizados
Journal

artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista Latinoamericana de Psicología
versão impressa ISSN 0120-0534
Rev. Latinoam. Psicol. v.39 n.1 Bogotá abr. 2007
ARTÍCULOS
Corteza frontopolar humana: área 10
Human frontopolar cortex: area 10
Efraín Buriticá-Ramírez; Hernán José Pimienta-Jiménez 1, 2
Universidad del Valle, Cali, Colombia
ABSTRACT
The frontopolar cortex, specifically area 10, has been related to complex cognitive processes such as planning, introspection, retrospective and prospective memories, and problem solving that implies simultaneous tasks. This information has been derived from functional neuroimaging. In spite of this knowledge, the cellular organization of area 10, the neurochemistry and the connection pattern has been less established. In the present work we attempt to summarize some relevant information related to anatomical, cytoarchitectural, chemoarchitectural and connectivity of area 10. We included some observations of our group derived from postmorten human tissue. This information may be important in order to establish a structural and functional correlation in the context of the organization of the human frontal lobe.
Keywords: Prefrontal cortex, Human, Area 10, Citoarchitecture, Connectivity, Complex cognitive processes.
RESUMEN
La corteza frontopolar, y en particular, el área 10, está vinculada con procesos cognitivos muy complejos como la planeación, la introspección, la memoria retrospectiva y prospectiva, la disociación de la atención y resolución de problemas que implican tareas simultáneas. Esta información se ha derivado de estudios con neuroimagenología funcional. A pesar de lo anterior, su organización celular, neuroquímica y patrón de conexiones han sido poco reconocidos. En el presente trabajo se pretende una descripción anatomo-funcional del área 10, cubriendo aspectos anatómicos, citoarquitectónicos, quimioarquitectónicos y de conectividad. Se incluyen algunas observaciones de nuestro grupo en tejido humano postmortem. La información considerada es útil para establecer una aproximación estructural y funcional de esta área en el contexto de la organización del lóbulo frontal del humano.
Palabras clave: Corteza prefrontal, Humano, Área 10, Citoarquitectura, Conectividad, Procesos cognitivos complejos.
INTRODUCCIÓN
Aunque la corteza cerebral presenta algunos patrones de organización comunes en diferentes sectores, es una estructura con variaciones regionales muy amplias, estableciéndose fundamentalmente dos extremos, la alocorteza, conformada por el paleocórtex y el arquicórtex, y la neocorteza. El arquicórtex está representado por el giro hipocampal, mientras que el paleocórtex corresponde a las regiones olfatorias. De acuerdo con Sanides (1969, en Escobar y col., 1998), a partir de este ancestro cortical se da una progresión incrementando el número de láminas hasta llegar al patrón hexalaminar de la neocorteza. Es necesario señalar que existen zonas de transición entre los dos extremos, como el mesocórtex, característico del giro cíngulo, y el proisocórtex característico de la corteza entorrinal en la base del lóbulo temporal (Pandya y Yeterian, 1990; Barbas, 1992). Las áreas corticales que surgen desde el arquicórtex hasta alcanzar el neocórtex se vinculan con la función espacial, y se relaciona con la pregunta DÓNDE. La corteza de origen olfatorio que progresa hacia el neocórtex, se relaciona con el proceso de identificar los objetos o estímulos, respondiendo a la pregunta QUÉ (Yeterian y Pandya, 1991). Las zonas más complejas de la neocorteza, corresponden a áreas multimodales del lóbulo temporal, y especialmente del lóbulo frontal, en las cuales se integra información de varias modalidades incluyendo aspectos emocionales y cognitivos.
La neocorteza caracteriza por su desarrollo a los mamíferos y, particularmente, a los primates no humanos y al hombre. A través de sus interacciones cortico-corticales, los sistemas de regulación cortico-estriado-tálamo-cortical y las fibras de proyección, la corteza cerebral ejerce influencia directa sobre la planificación, la intención y la ejecución voluntaria del movimiento. Además se relaciona con la esfera consciente, el lenguaje, el pensamiento, la memoria, la función emocional y, en general con todas aquellas actividades distinguidas como funciones mentales superiores (Kupfermann, 1991).
Es importante destacar que los estudios comparativos entre mamíferos no primates y primates determinan una relativa miniaturización de las áreas primarias sensoriales en los últimos, a favor de un crecimiento de las áreas asociativas, entre estas la corteza prefrontal (Kaas, 1995). Las áreas prefrontales definidas macroscópicamente, atendiendo a accidentes superficiales, corresponden, en el hombre al 29% del total de la corteza, en el chimpancé al 18%, en el macaco al 7% y en el conejo al 3% (Brodmann, 1905; en Pimienta y col., 1999). La región prefrontal de la corteza cerebral en los primates no humanos y en el hombre, se ubica entre el polo frontal y la corteza premotora del lóbulo frontal. En la superficie lateral, se ubican las áreas prefrontales 9, 10p, 11, 47/12 y 46 (ver figura 1); en la superficie medial, las áreas 9, 10p, 10r, 10m, 24, 25 y 32; y en la superficie orbital, las áreas 10p, 11, 47/12, 13, 14 y las porciones anteriores de la ínsula (Petrides y Pandya, 1999; Öngür y col., 2003).
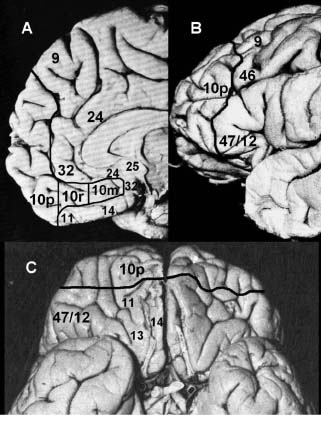
Figura 1. Localización anatómica aproximada de la corteza frontopolar o área 10p en el cerebro humano. A: Ubicación del área 10p en la superficie medial de la corteza prefrontal, de acuerdo con la nomenclatura de Öngür y col. (2003). B: Ubicación del área 10p en la superficie lateral de la corteza prefrontal, de acuerdo con la nomenclatura de Petrides y Pandya (1999). C: Ubicación del área 10p en la superficie orbital de la corteza prefrontal, de acuerdo con la nomenclatura de Öngür y col. (2003).
Las funciones de la corteza prefrontal (CPF) dependen de las interacciones intrafrontales, de las conexiones con otras regiones corticales y con estructuras subcorticales, principalmente el estriado y el tálamo; sin embargo, a las diferentes superficies se les ha vinculado con algunas funciones particulares. De esta forma, las áreas que constituyen la cara medial se asocian con la atención, la motivación (interés) y con el control autonómico (Pandya y Yeterian, 1990; Öngür y Price, 2000; Van Eden, 2000). La región orbitaria se relaciona con la integración sensorio-emocional, con algunos aspectos de la personalidad y con la conducta social (Pandya y Yeterian, 1990; Cavada y col, 2000; Öngür y Price, 2000). Por su parte a la superficie lateral se la considera fundamental en los procesos cognitivos, memoria operativa y atención selectiva (Pandya y Yeterian, 1990; Fuster, 2000).
El área 10, que incluye sectores del polo frontal y extensiones en la corteza medial, está presente en la corteza prefrontal de todos los primates; sin embargo, los estudios comparativos de Semendeferi y col. (2001), han demostrado que un mayor grado de desarrollo de esta región del lóbulo frontal caracteriza al cerebro humano. Además, recientemente se han empezado a demostrar algunas particularidades anatómicas, citoarquitectónicas y de conectividad, las cuales pueden distinguirla de las demás áreas prefrontales, particularmente orbitales y mediales. Desde el punto de vista funcional, el área 10 se ha vinculado con aspectos considerados propiamente humanos como la "divagación" mental, la Teoría de la Mente y la evaluación de los estados mentales internos y externos (Ramnani y Owen, 2004). Lo anterior también es de importancia clínica, dado que algunas de estas funciones se alteran en desordenes psiquiátricos como la esquizofrenia.
Aunque el área 10 tiene representación en las tres superficies de la corteza prefrontal, asociadas cada una con funciones particulares, esta región parece estar vinculada funcionalmente con procesos más cognitivos que emocionales tales como la introspección y la coordinación del pensamiento orientado por estímulos, y el pensamiento independiente de los mismos (Pandya y Yeterian, 1990; Fuster, 2000; Ramnani y Owen, 2004). En contraste, las áreas orbitales adyacentes (11, 12, 13, 14 y las porciones anteriores de la ínsula), se asocian con el control de impulsos, la toma de decisiones, el juicio y el análisis en contexto de la información visceral y emocional; y las regiones mediales posteriores (áreas 24, 25 y 32) se relacionan con respuestas visceromotoras, y el control de la motivación y la atención durante tareas de alta precisión (Öngür y Price, 2000; Van Eden, 2000).
Dado estos nuevos conocimientos, en el presente trabajo se pretende destacar algunas características del área 10 que la distinguen de las áreas prefrontales más posteriores respecto de la organización laminar de las neuronas, el patrón de conexiones cortico-corticales y la integración de los circuitos cortico-estriado-tálamo-corticales. Este análisis es importante porque permite aproximaciones estructurales a las hipótesis funcionales del área 10, como la "hipótesis de la compuerta" (Gateway Hypothesis) (Gilbert y col., 2005).
UBICACIÓN ANATÓMICA EN EL HUMANO
De acuerdo con la nomenclatura de Brodmann (1909), el área 10 cubre el polo frontal en las superficies medial y lateral. Walker (1940), utilizando corteza cerebral de macacos, extendió a la cara basal de la corteza prefrontal el área 10. Según el estudio de Barbas y Pandya (1989), el área 10 estaría dividida en 2 sectores, diferenciados por su origen filogenético cortical: a) El área 10 orbital haría parte de la tendencia ventro-lateral que se originó en el paleocórtex; y b) el área 10 de las superficies medial y lateral que formaría parte de la tendencia medio-dorsal originada en el archicortéx (Barbas y Pandya, 1989). Recientemente, Öngür y col. (2003), extienden aún más el área 10 humana en la superficie medial hasta alcanzar el giro cíngulo en la vecindad del rostro del cuerpo calloso, y la subdividen en tres porciones en sentido rostro caudal, partiendo desde el polo frontal: polar (10p), rostral (10r) y medial (10m). En el presente trabajo, el área 10 hará referencia a la porción 10p, de acuerdo con la nomenclatura de Öngür y col. (2003) de la corteza prefrontal del humano.
Como se mencionó anteriormente, el área 10 corresponde al polo frontal; por lo tanto incluye tres superficies que convergen en el extremo anterior, una lateral, una medial y una basal. La superficie lateral, se relaciona posteriormente con la región anterior del área 9, correspondiente a la región anterior del giro frontal superior; con la región anterior del área 46 perteneciente al giro frontal medio (ver figura 1); y con el área 47/12 del giro frontal inferior (Petrides y Pandya, 1999). La basal está limitada posteriormente por los giros orbitarios correspondientes al área 11 (Öngür y col., 2003). Los limites y relaciones de la superficie medial del área 10 son más extensos y complejos, dado que este sector del área 10 no se restringe al polo frontal sino que se prolonga en dirección caudal hasta aproximarse al área 25, en la vecindad del rostro del cuerpo calloso, en la denominada región septal (Öngür y col., 2003). De acuerdo con lo anterior, las relaciones de este sector son las siguientes: dorsalmente y en sentido antero-posterior, es vecina sucesivamente del área 9 medial, el área 32, el área 24 y el área 25; ventralmente, el área 10 en la superficie medial se relaciona sucesivamente con la región medial del área 11 y la prolongación medial del área 14 (Öngür y col., 2003).
Por su extensión, el área 10 distingue a la corteza prefrontal del humano de la del resto de primates (Semendeferi y col., 2001). De las cortezas de alto nivel de integración de la información (cortezas multimodales y supramodales), las neuronas de las áreas prefrontales laterales (incluyendo el área 10) exhiben el mayor grado de ramificación dendrítica, y la más alta densidad de espinas, en las dendritas basales de las neuronas piramidales de la lámina III (Jacobs y col., 2001). Inclusive, la superficie ocupada por las dendritas basales de las neuronas piramidales de la lámina III del área 10, es significativamente mayor que en las áreas vecinas 11 y 12 (Elston, 2000). Por lo tanto, se ha sugerido que las regiones más anteriores de la corteza prefrontal en los humanos tienen más espacio para conexiones extrínsecas e intrínsecas en las capas supragranulares, lo cual puede indicar una tendencia evolutiva al incremento en la comunicación entre la corteza prefrontal dorsolateral y otras áreas asociativas de orden superior, como aquellas ubicadas en las superficies medial y basal del lóbulo frontal, en el lóbulo temporal, y en la transición parieto-temporo-occipital (Barbas, 1995; Semendeferi y col., 2001).
ORGANIZACIÓN CELULAR Y NEUROQUÍMICA
Citoarquitectura
En los últimos cinco años, se comenzó a precisar la organización laminar de la corteza frontopolar humana (Semendeferi y col., 2001; y en nuestro grupo, Perlaza y col., 2005). A continuación se resumen los principales hallazgos en este sentido. La lámina I del área 10 es delgada, con un espesor relativo del 11% del espesor total cortical (ver figura 2). La lámina II es también delgada, fácilmente identificable, con un espesor relativo cercano al 9%, incluye células con formas piramidales y granulares pequeñas. La lámina III es la más extensa en el polo frontal, con 31% de espesor relativo; las células más superficiales son pequeñas mientras que en los sectores más profundos de la capa, el tamaño de las células es mayor. La lámina IV es delgada (entre 6% y 9% del espesor cortical total), y sus límites con las láminas III y V son fácilmente discernibles; contiene predominio de células granulares pequeñas. La lámina V posee un espesor relativo del 17%, con predominio de células piramidales grandes. Esta lámina presenta dos subcapas, Va y Vb, y la densidad celular en esta última subcapa (más cerca de la lámina VI) es menor que en la primera. La lámina VI tiene 23% de espesor relativo; e incluye células piramidales medianas y fusiformes; sus bordes con la lámina V son evidentes.
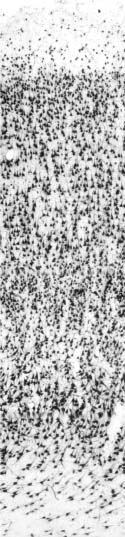
Figura 2. Microfotografía de una sección de tejido de 50 µm de espesor, correspondiente al área 10 humana. La inmunomarcación con el anticuerpo AntiNeuN, permite ver toda la población neuronal y su distribución laminar. SB: sustancia blanca. La barra es equivalente a 200 µm.
Si sumamos las láminas II y III (capas supragranulares) y las láminas V y VI (capas infragranulares), se encuentra que las primeras tienen un espesor relativo del 40-43%, y las segundas, 39-41% (Semendeferi y col., 2001; Perlaza y col., 2005). Como lo veremos, esto distingue al área 10 de otras áreas prefrontales adyacentes, y puede tener implicaciones en los patrones de conectividad asociados a las funciones cognitivas y emocionales.
Es importante señalar que el área 10 comparte algunas características citoarquitectónicas con sectores adyacentes de la corteza prefrontal dorsolateral y la porción medial del área 9, y difiere en otras, de las regiones orbitofrontales posteriores y las regiones mediales ventrales y caudales a ella. La corteza orbitofrontal, en general, presenta una tendencia en sentido rostro-caudal a tener unas láminas infragranulares más prominentes en relación con las supragranulares, una menor definición de las láminas y una lámina IV que progresivamente se hace imperceptible. Al comparar el área 10 con las regiones prefrontales laterales adyacentes (áreas 9, 46 y 47/12), se encuentra que: 1) la lámina II es más delgada y más densamente poblada; 2) el grado de desarrollo de las láminas III y IV es similar al de las áreas 9 y 46, que corresponde a las zonas más especializadas del lóbulo frontal; 3) la relación entre las capas supragranulares e infragranulares es proporcional (Petrides y Pandya, 1999; Semendeferi y col., 2001; Perlaza y col., 2005). Considerando que las capas supragranulares son la principal fuente de las conexiones cortico-corticales intrahemisféricas e interhemisféricas, y las capas infragranulares, la fuente de las conexiones cortico-talámicas, cortico-estriatales, y en general, cortico-subcorticales; el área 10, al igual que las otras áreas prefrontales dorsolaterales, establecería de forma proporcional conexiones con otras regiones corticales y estructuras subcorticales, vinculándose más en procesos cognitivos, que las regiones mediales y orbitales posteriores, más vinculadas con contenidos emocionales.
La prominencia de la lámina III en el área 10, comparada con el resto de las áreas prefrontales (31%), podría tener implicaciones importantes en la integración bihemisférica considerando que esta capa en los primates constituye el origen principal de las fibras comisurales. Hasta el presente desconocemos si estas interacciones podrían correlacionarse con la actividad cooperativa bihemisférica en tareas de manipulación de información on-line, conocida como la memoria operativa. Observaciones de nuestro grupo han probado una simetría entre la distribución de las láminas III de ambos hemisferios, lo cual podría ser indicativo de complementariedad durante tareas específicas (datos sin publicar).
Quimioarquitectura
Con el desarrollo de la inmunohistoquímica y de marcadores de subpoblaciones neuronales específicas, se inició recientemente la caracterización neuroquímica de diversos sectores de la corteza cerebral de los primates, entre ellos, la corteza prefrontal anterior.
El estudio de la quimioarquitectura del área 10 en la corteza cerebral humana, se ha llevado a cabo con los marcadores para SMI-32, el cual marca proteínas de neurofilamentos en neuronas piramidales, y para parvalbúmina, calbindina y calretinina, los cuales permiten identificar las proteínas que ligan calcio en subpoblaciones neuronales, particularmente interneuronas (ver figura 3). Estas proteínas identifican neuronas que hacen contactos sinápticos inhibitorios sobre las células piramidales (DeFelipe, 1997; Markram y col., 2004).
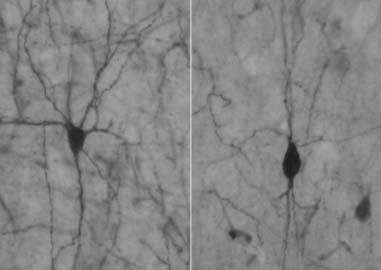
Figura 3. Microfotografía de neuronas inhibitorias de la corteza frontopolar humana. A: Interneurona multipolar inmunorreactiva para el anticuerpo AntiParvoalbúmina. B: Interneurona bipenachada inmunorreactiva para el anticuerpo AntiCalbindina. La barra es equivalente a 20 µm.
La marcación con SMI-32 se ha encontrado, en general, en neuronas piramidales grandes de las capas III y V del área 10 en el polo frontal, tanto en las superficies medial como orbital (Hof y col., 1995; Öngür y col., 2003). En cada una de las superficies de la corteza prefrontal, la marcación con SMI-32 muestra una progresión caudo-rostral, específicamente en la lámina III (Hof y col., 1995; Öngür y col., 2003). Así, las porciones 10m y 10r de la cara medial tienen una menor cantidad de neuronas positivas para SMI 32 que la porción 10p (Öngür y col., 2003). En la superficie orbital, el polo frontal, se caracteriza por la presencia de una población relativamente pequeña de neuronas de marcaje claro en la parte profunda de la lámina III, y una densa población de neuronas marcadas de manera oscura en la parte superior de la lámina V (Hof y col., 1995), lo cual le da una apariencia de doble banda, al igual que en la primera superficie mencionada. La lámina VI tiene pocas neuronas y de menor tamaño que las de la lámina V, y la lámina IV aparece desprovista de inmunomarcación (Hof y cols., 1995). La porción orbital del área 10 presenta una mayor densidad de neuronas inmuno-reactivas para SMI-32, que las demás regiones de esta misma superficie (Hof y col., 1995).
Dombrowski y col. (2001), estudiando diversas áreas prefrontales del Mono Rhesus, identificaron una progresión en la marcación de la parvalbúmina. La densidad de neuronas inmuno-reactivas para parvalbúmina aumenta de las áreas prefrontales clasificadas como agranulares, hacia las disgranulares y eulaminadas, siendo más alta en estas últimas, entre las que se encuentra el área 10 (Dombrowski y col., 2001). Según Öngür y col. (2003), el marcaje con parvalbúmina en sujetos humanos, muestra neuronas inmuno-reactivas en las láminas II a VI, y un plexo denso de fibras en las capas corticales medias. Hof y col. (1995), observaron que el neuropilo marcado con parvalbúmina (también en tejido humano) en las capas III y V fue mucho más pronunciado en las porciones anteriores del giro recto, incluyendo la región frontopolar, que en las porciones posteriores del mismo. Esta diferencia se hace más obvia en las cortezas con un mayor desarrollo de la lámina IV, que separa los dos parches de neuropilo (Hof y col., 1995; Öngür y col., 2003). Los patrones laminares de marcación de la parvalbúmina descritos en el área 10 de la corteza cerebral del Mono Rhesus (Dombrowski y col., 2001), fueron similares a los observados en el área 46 (Arteaga y col., 2004), y en el área 11 (Öngür y col., 2003) de la corteza prefrontal del humano.
Dombrowski y col. (2001), también encontraron una progresión en la inmunomarcación con calbindina. La densidad de neuronas inmuno-reactivas para calbindina disminuye de las áreas prefrontales clasificadas como agranulares, hacia las disgranulares y eulaminadas, siendo más baja en estas últimas, que incluyen el área 10 medial (Dombrowski y col., 2001). La marcación con calbindina, en las cortezas eulaminadas, muestra un patrón de doble banda: una densamente poblada en capas II-III, con 2 o 3 veces más neuronas que en la banda correspondiente a las capas V-VI (Dombrowski y col., 2001; Arteaga y col., 2004). La mayoría de las neuronas positivas para calbindina tienen forma no-piramidal, y las de formas piramidales más grandes se hallan en la parte profunda de la lámina III y en la parte superficial de la lámina V (Dombrowski y col., 2001; Arteaga y col., 2004). Arteaga y col. (2004), describieron la inmunomarcación con calbindina en el área 46 de la corteza prefrontal dorsolateral humana, como más intensa y más nítida en capas supragranulares, con predominio de formas no-piramidales en estas; mientras en las capas infragranulares la marcación es más tenue, pero más amplia y abarca una gran cantidad de neuronas piramidales medianas. En general, el número de neuronas positivas para calbindina, parece doblar el número de neuronas positivas para parvalbúmina, tanto en capas supragranulares como infragranulares (Dombrowski y col., 2001; Arteaga y col., 2004).
La calretinina está presente en interneuronas localizadas principalmente en las capas II y III; mientras las capas IV, V y VI, tienen una menor densidad de células marcadas (Dombrowski y col., 2001). La distribución de las neuronas y el neuropilo marcados con calretinina son más homogéneos que los de parvalbúmina y calbindina, y no presentan particularidades laminares o regionales entre las cortezas prefrontales de primates (Hof y col., 1995; Dombrowski y col., 2001).
CONECTIVIDAD
El nivel de desarrollo y definición de las láminas corticales en las áreas prefrontales, se ha convertido en un criterio estructural de agrupación de las mismas, que permite predecir la dirección del patrón de conexiones de un área con otras regiones corticales (Barbas y Rempel-Clower, 1997). De esta manera, contando únicamente con las referencias de la conectividad cerebral en primates no humanos, se pueden predecir los patrones de conexiones entre regiones corticales humanas a partir del establecimiento de su organización laminar y celular.
Conexiones cortico-corticales
Barbas y Rempel-Clower (1997), establecieron cinco categorías citoarquitectónicas en la corteza prefrontal, 1-5, atendiendo al grado de granularidad y diferenciación laminar, siendo los niveles más altos los de mayor granularidad y definición laminar. Estos niveles se correlacionan con patrones de conectividad. De acuerdo con el patrón de conexiones cortico-corticales establecido por Barbas y Rempel-Clower (1997), el área 10 tendría conexiones predominantes con las áreas que comparten su mismo nivel de diferenciación laminar (nivel 4), es decir, las porciones laterales de las áreas 12 y 9, y la porción rostral del área 46; seguidas por las conexiones con las áreas pertenecientes al nivel 3 (área 14, área 11, porción orbital del área 12, y porción medial del área 9) y del nivel 5 (área 8 y porción caudal del área 46).
Las conexiones del área 10 con áreas prefrontales mediales y orbitales posteriores serían menos abundantes, al igual que con áreas de otras regiones de la corteza cerebral. Las conexiones del área 10 con otras áreas de niveles con definición laminar inferior, surgen en su gran mayoría de las capas supragranulares, para terminar en capas infragranulares; las conexiones hacia áreas con niveles de definición laminar superior, parten en su mayoría de capas infragranulares, para alcanzar las capas supragranulares. Cuando el área 10 se conecta con las áreas prefrontales que comparten su mismo nivel de definición laminar, no se establece un patrón segregado de conexiones cortico-corticales, por el contrario, las capas supragranulares generan proyecciones que se extienden a las capas supra e infragranulares y viceversa (Barbas y Rempel-Clower, 1997).
Independientemente de la fuerza de las conexiones que establece con otras áreas prefrontales, el área 10 en los primates se conecta efectivamente en su superficie orbital con las áreas 8 y 46 (Petrides y Pandya, 1999), y las regiones de transición entre las áreas 10 y 11, 11 y 13, y 13 y 12 (Cavada y col., 2000); en la superficie lateral se conecta con las áreas 9 y 46 (Petrides y Pandya, 1999; Cavada y col., 2000), y el área 12 (Cavada y col., 2000); y en la superficie medial se conecta con las áreas 9/46 y 46 (Petrides y Pandya, 1999; Cavada y col., 2000), las áreas 8, 32, 14, 24, y las áreas 23, 29 y 30 en la región posterior del giro cíngulo (Morecraft y Van Hoesen, 1993; Cavada y col., 2000). Al parecer, cada área prefrontal se conecta contralateralmente con su área homóloga y las áreas homólogas de sus áreas vecinas más cercanas (Cavada y col., 2000). La mayoría de estas conexiones, al igual que las del resto de la corteza prefrontal, son recíprocas (ver figura 4).
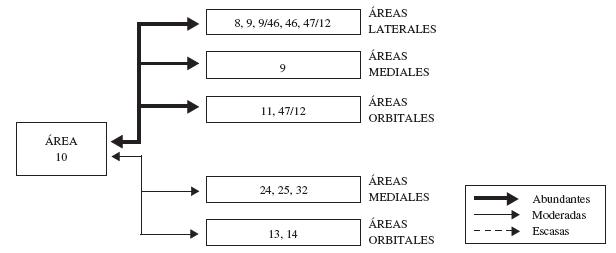
Figura 4. Representación esquemática de la organización de las conexiones del área 10 con otras áreas de la corteza frontal. En esta figura, se indica la superficie en la cual se ubica cada conjunto de áreas, se discriminan estas de acuerdo con la fuerza de sus conexiones (abundantes, moderadas o escasas), así como la dirección de las mismas. Para mayor detalle, ver texto.
Si bien se ha establecido una organización de las proyecciones desde cortezas parietales y occipitales hacia las áreas 8, 46, 11, 12 y 32 de la región prefrontal (Barbas, 1988; Barbas, 1995), no hay evidencia de proyecciones desde dichas regiones hacia la corteza frontopolar.
El área 10 se conecta recíprocamente con el lóbulo temporal (ver figura 5), particularmente con la región parahipocampal (Bachevalier y col., 1997; Cavada y col., 2000), la mitad dorsal del polo temporal (Bachevalier y col., 1997; Kondo y col., 2003), la corteza asociativa de la porción rostral del giro temporal superior (Hackett y col., 1999), y las áreas auditivas ubicadas en éste mismo giro (Bachevalier y col., 1997; Cavada y col., 2000). El área TE1, según los mapas de Seltzer y Pandya (1978), en la parte anterior de la corteza temporal inferior, se conecta de forma moderada con la porción dorsolateral del área 10, y las terminaciones de las fibras de estas conexiones se ubican en su mayoría en las capas supragranulares (Rempel-Clower y Barbas, 2000).
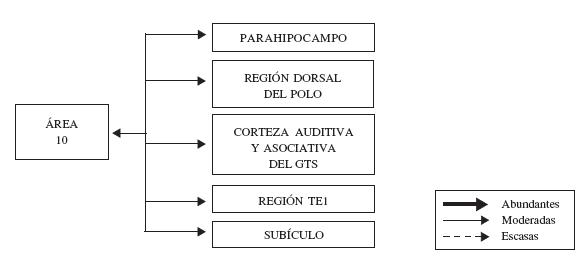
Figura 5. Representación esquemática de la organización de las conexiones del área 10 con el lóbulo temporal. En esta figura, se indica cada región del lóbulo temporal cuya conexión con la corteza frontopolar se ha corroborado, se discrimina la fuerza de estas conexiones (abundantes, moderadas o escasas), así como la dirección de las mismas. Para mayor detalle, ver texto.
En contraste con las regiones prefrontales mediales y orbitales posteriores, las áreas prefrontales dorsolaterales, y 9 y 10 medial, no envían proyecciones a la amígdala (Ghashghaei y Barbas, 2002), aunque si recibe algunas aferencias de esta estructura límbica (Barbas y De Olmos, 1990). Este hecho reviste importancia, porque la corteza frontopolar tendría escasa influencia en el desencadenamiento de acciones autonómicas que surgen desde la amígdala hasta el hipotálamo y centros autonómicos craneales y espinales.
Según Öngür y Price (2000), las áreas prefrontales 11 y 32, se encargan de integrar la información sensorial y límbica que han obtenido de áreas prefrontales mediales y orbitales más caudales, y luego la envían a las áreas de la región frontopolar. Si bien se han encontrado una amplia conexión de las cortezas prefrontales mediales y orbitales, con las estructuras límbicas, las conexiones del área 10 con estas son muy limitadas. Hasta el momento solo se ha encontrado terminaciones de fibras provenientes del subículo (formación hipocampal), en la porción orbital del área 10 (Öngür y Price, 2000).
Conexiones cortico-subcorticales
La corteza prefrontal, en general, se conecta con el tálamo a través de axones que parten de neuronas piramidales de la lámina VI y la parte profunda de la lámina V; y recibe fibras tálamocorticales en la lámina IV y en la parte profunda de la lámina III (Erickson y Lewis, 2004). El área 10 dorsolateral se conecta recíprocamente con la región antero-dorsal del núcleo dorsomediano del tálamo (ver figura 6), específicamente con las porciones densocelular y parvocelular de éste (Bachevalier y col., 1997). Las porciones medial y orbital del área 10, se conectan principalmente con el eje medial del núcleo caudado, éste se conecta con el globo pálido ventral, que a su vez envía fibras a la región medial del núcleo dorsomediano del tálamo, el cual se conecta recíprocamente con las regiones en mención (Ferry y col., 2000).

Figura 6. Representación esquemática de la organización de las conexiones del área 10 con el tálamo. En esta figura, se indican las cuatro regiones talámicas cuyas conexiones con la corteza frontopolar se ha corroborado, se discrimina la fuerza de estas conexiones (abundantes, moderadas o escasas), así como la dirección de las mismas. Para mayor detalle, ver texto.
De las áreas dorsolaterales de la corteza prefrontal, el área 10, en sus porciones dorsal y ventral, es el sector más fuertemente conectado con la región medial del núcleo anterior del tálamo (ver figura 6) (Xiao y Barbas, 2002). También se ha corroborado la presencia de proyecciones cortico-talámicas a la porción magnocelular del núcleo ventral anterior (Erickson y Lewis, 2004).
La región medial del núcleo pulvinar del tálamo establece conexiones con amplios sectores de la corteza asociativa, entre ellos con la corteza prefrontal (ver figura 6) (Romanski y col., 1997). Si bien estas conexiones pulvinarprefrontal son menos intensas que con otros sectores corticales, están organizadas topográficamente (Romanski y col., 1997). El área 10 dorsal y medial, al igual que las áreas 9, 12, 24, 25 y 32, son blanco de la parte medial del núcleo pulvinar medial; porción talámica del que también reciben proyecciones el polo temporal, las regiones rostrales del giro y del surco temporal superior, la amígdala y la región anterior del giro cíngulo (Romanski y col., 1997). Las áreas prefrontales también envían fibras a éste núcleo, pero en menor proporción que las conexiones ascendentes (Romanski y col., 1997).
Las conexiones de la corteza prefrontal y el hipotálamo han sido estudiadas por Rempel-Clower y Barbas (1998). Estos trabajos, establecieron que las áreas prefrontales mediales y orbitales posteriores (cortezas disgranulares y agranulares), se comunican ampliamente de manera recíproca, y que las conexiones ascendentes son menos específicas que las descendentes. El área 10 dorsolateral recibe de la región dorsal del hipotálamo (ver figura 7), las divisiones tuberal y posterior de las regiones lateral y medial del mismo, y de los núcleos tubero-mamilar y para-mamilar en la región lateral hipotalámica (Rempel-Clower y Barbas, 1998). Las áreas prefrontales dorsolaterales parecen tener muy pocas o ninguna neurona de proyección que se dirijan hacia las regiones hipotalámicas mencionadas anteriormente (Rempel-Clower y Barbas, 1998).
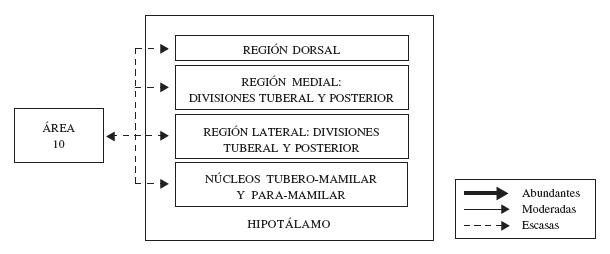
Figura 7. Representación esquemática de la organización de las conexiones del área 10 con el hipotálamo. En esta figura, se indica cada región del hipotálamo cuya conexión con la corteza frontopolar se ha corroborado, se discrimina la fuerza de estas conexiones (abundantes, moderadas o escasas), así como la dirección de las mismas. Para mayor detalle, ver texto.
De la corteza prefrontal, las áreas de la superficie dorsolateral, principalmente las áreas 9 y 46, son las que más proyecciones envían a la región basilar del puente; a través de circuitos ponto-cerebelosos, estas regiones del frontal influyen en la actividad cerebelosa (ver figura 8). El circuito se completa por proyecciones entre los núcleos dentado del cerebelo al núcleo dorsomediano del tálamo, y de este a la corteza frontopolar (Schmahmann y Pandya, 1997; Middleton y Strick, 2001). Este circuito reviste importancia dado que implica al cerebelo en actividades diferentes a la motora, es decir, lo relacionaría con la regulación de la función cognitiva y emocional.
El área 10 medial recibe proyecciones colinérgicas de las porciones anterior, intemedio y posterior de los núcleos Basales de Meynert, y de forma ligera desde el núcleo de la Banda Diagonal (ver figura 8) (Ghashghaei y Barbas, 2001). La porción dorsolateral del área 10 recibe una proyección densa de las divisiones intermedia y posterior de los núcleos Basales de Meynert, moderada de la división anterior, y prácticamente no recibe proyecciones del núcleo de la Banda Diagonal (Ghashghaei y Barbas, 2001). El área 10 no envía proyecciones de vuelta hacia las estructuras basales del telencéfalo mencionadas (Ghashghaei y Barbas, 2001).
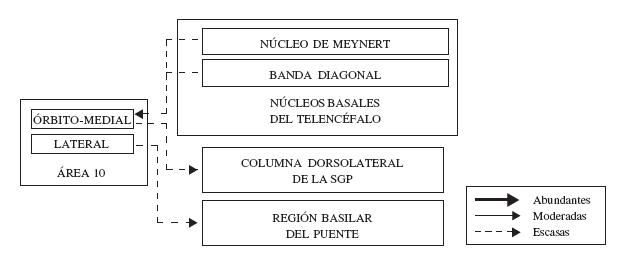
Figura 8. Representación esquemática de la organización de las conexiones del área 10 con estructuras de la base del telencéfalo y el tallo cerebral. En esta figura, se indica dos núcleos basales del telencéfalo y dos estructuras del tallo cerebral cuya conexión con la corteza frontopolar se ha corroborado, se discrimina la fuerza de estas conexiones (abundantes, moderadas o escasas), así como la dirección de las mismas. Para mayor detalle, ver texto.
Finalmente, es importante señalar que las regiones mediales de la corteza prefrontal, entre estas, las áreas 10, 25 y 32, tienen influencia sobre algunas estructuras límbicas como la sustancia gris periacueductal (ver figura 8) (Öngür y Price, 2000). Por esta ruta, la corteza frontopolar ejercería influencia sobre actividades autonómicas asociadas a estadíos emocionales.
ASPECTOS FUNCIONALES
La función del área 10 es de las menos comprendidas. Hay evidencias que la vinculan con el procesamiento cognitivo (Burgess y col., 2005). Las dificultades para establecer una función específica para la corteza prefrontal rostral tienen origen entre otros, en: la falta de estudios sobre la actividad de neuronas frontopolares de monos; la escasez de estudios citoarquitectónicos y quimioarquitectónicos; la imposibilidad de estudios de conectividad en humanos; la ausencia en la naturaleza, de lesiones cerebrales en el humano específicas de esta área cortical. Sin embargo, existe una gran cantidad de datos obtenidos a partir de estudios de neuroimagenología funcional, que son interpretados a la luz de los paradigmas convencionales acerca de la relación estructura-función (Ramnani y Owen, 2004).
Las lesiones del área 10 típicamente no producen signos neurológicos fácilmente reconocibles (tales como hemiparesia, afasia, etc.), a menos que sean resultado de traumas que cubran grandes regiones cerebrales (Burgess y col., 2005). La neuroimagenología funcional es la fuente de la mayoría de los datos relevantes sobre la función del área 10, pero estos presentan un problema, y es que la activación de la corteza frontopolar se encuentra en una amplia variedad de tareas que incluyen desde condicionamiento hasta fenómenos complejos como la memoria y el juicio (Koechlin y col., 1999), o la resolución de problemas (Christoff y col., 2001), lo cual ha impedido la teorización sobre la misma. La siguiente es una lista de las tareas en las que se ha involucrado la activación del área 10: 1) recuperación de información episódica verbal y no verbal, 2) memoria semántica, 3) lenguaje, 4) aprendizaje motor, 5) aprendizaje de normas, 6) aprendizaje por condicionamiento, 7) memoria de trabajo verbal y no verbal, 8) memoria espacial, 9) percepción auditiva, 10) procesamiento de objetos, y 11) razonamiento (Burgess y col., 2005).
Burgess y col. (2005), agruparon las teorías que explican la función del área 10 de la siguiente manera: 1) aquellas que la relacionan con la memoria episódica (Ranganath y col., 2003); 2) las que asocian a esta región con la función metacognitiva y la Teoría de la Mente, la primera entendida como la reflexión consciente acerca de los propios pensamientos dirigidos a metas (Johnson y col., 2002), y la segunda como la atribución a otros de estados mentales diferentes de los propios (Christoff y Gabrieli, 2000); 3) esta categoría incluye dos aspectos: el anatómico, que se fundamenta en las conexiones del área 10 con el área 46 apoyando a esta última en el rescate y manipulación de la información propias de la memoria operativa (working memory) (Fletcher y Henson, 2001), y el de procesamiento, vinculado con el control sobre varios tipos de procesamiento relativamente independientes, necesarios para la consecución de una meta y sus respectivas submetas (Koechlin y col., 1999; 2003; Ramnani y Owen, 2004); y 4) las teorías que relacionan la corteza frontopolar con la valoración de información proveniente del medio interno y externo (Christoff y col., 2004).
Recientemente, ha surgido una nueva hipótesis que pretende sintetizar la mayoría de la evidencia proveniente de los estudios de lesiones cerebrales en el humano e imagenología funcional, así como las teorías acerca de la función del área 10 ya mencionadas. La hipótesis de la "compuerta" ("Gateway Hypothesis"), como ha sido denominada por sus autores, sugiere que la corteza prefrontal rostral está involucrada en la coordinación del pensamiento independiente de estímulos y el pensamiento orientado por estímulos dirigida a la consecución de metas (Gilbert y col., 2005). Dicha coordinación sería indispensable en aquellas situaciones donde las formas predominantes o preestablecidas de respuesta o comportamiento, no conducen a la obtención de los resultados óptimos (Burgess y col., 2005).
De igual manera, recientemente se ha recopilado evidencia que sostiene la disociación funcional entre las superficies medial y lateral del área 10 (Koechlin y col., 2000; Burgess y col., 2005). De acuerdo con los experimentos con pruebas "multitarea" y neuroimagenología funcional, la superficie medial de la corteza frontopolar desvía la atención hacia el pensamiento orientado por estímulos sensoriales, y la superficie lateral centraría la atención en las ideas generadas internamente (Burgess y col., 2005); sin embargo, los autores de la propuesta no descartan la posibilidad de que en ciertas situaciones, ambas formas de pensamiento deban trabajar concertadamente.
REFERENCIAS
Arteaga, G., Pimienta, H.J. & Escobar, M.I. (2004). Neuronas inmunorreactivas a Parvalbúmina y Calbindina en la corteza prefrontal dorsolateral de sujetos humanos. Acta Neurológica Colombian, 20, (3), 105-118. [ Links ]
Bachevalier, J., Meunier, M., Lu, M.X. & Ungerleider, L.G. (1997). Thalamic and temporal cortex input to medial prefrontal cortex in rhesus monkey. Experimental Brain Research, 115, 430-444. [ Links ]
Barbas, H. (1988). Anatomic organization of basoventral and medio-dorsal visual recipient prefrontal regions in the rhesus monkey. The Journal of Comparative Neurology, 276, 313-342. [ Links ]
Barbas, H. (1992). Architecture and cortical connections of the prefrontal cortex in the Rhesus monkey. Advances in Neurology, 57, 91-115. [ Links ]
Barbas, H. (1995). Anatomic basis of cognitive-emotional interactions in the primate prefrontal cortex. Neuroscience and Biobehavioral Reviews, 19, 3, 499510. [ Links ]
Barbas, H. & De Olmos, J. (1990). Projections from the amygdala to basoventral and medio-dorsal prefrontal regions in the rhesus monkey. The Journal of Comparative Neurology, 300, 549-571. [ Links ]
Barbas, H. & Pandya, D.N. (1989). Architecture intrinsic connections of the prefrontal cortex in the rhesus monkey. The Journal of Comparative Neurology, 286, 353-375. [ Links ]
Barbas, H. & Rempel-Clower, N. (1997). Cortical structure predicts the pattern of corticocortical connections. Cerebral Cortex 7, 635-646. [ Links ]
Brodmann, K. (1909). Vergleichende lokalisationslehre der grosshirnrinde in ihren prinzipien dargestellt auf grund des zellenbaues. Barth: J.A. [ Links ]
Burgess, P.W., Simons, J.S., Dumontheil, I. & Gilbert, S.J. (2005). The Gateway Hypothesis of rostral prefrontal cortex (area 10) function. En J. Duncan, L. Phillips y P. Mcleod (Eds.), Measuring the mind: Speed, control, and age. Oxford: Oxford University Press. [ Links ]
Cavada, C., Compañy, T., Tejedor, J., Cruz-Rizzolo, R.J. & Reinoso-Suárez, F. (2000). The anatomical connections of the macaque monkey orbitofrontal cortex: A review. Cerebral Cortex, 10, 220-242. [ Links ]
Christoff, K. & Gabrieli, J.D. (2000). The frontopolar cortex and human cognition: Evidence for a rostro-caudal hierarchical organization within the human prefrontal cortex. Psychobiology, 28, 2, 168-186. [ Links ]
Christoff, K., Prabhakaran, V, Dorfman, J., Zhao, Z., Kroger, J.K., Holyoak, K.J. & Gabrieli, J.D. (2001). Rostrolateral prefrontal cortex involvement in relational integration during reasoning. Neuroimage, 14, 1136-1149. [ Links ]
Christoff, K., Ream, J.M & Gabrieli, J.D. (2004). Neural basis of spontaneous thought processes. Cortex, 40, 623-630. [ Links ]
DeFelipe, J. (1997). Types of neurons, synaptic connections and chemical characteristics of cells immunoreactive for Calbindin-D28k, Parvalbumin and Calretinin in the neocortex. Journal of Chemical Neuroanatomy, 14, 1-19. [ Links ]
Dombrowski, S.M., Hilgetag, C.C. & Barbas, H. (2001). Quantitative architecture distinguishes prefrontal cortical systems in the rhesus monkey. Cerebral Cortex, 11, 975-988. [ Links ]
Elston, G.N. (2000). Piramidal cells of the frontal lobe: All the more spinous to think with. The Journal of Neuroscience, 20, 1-4. [ Links ]
Erickson, S.L. & Lewis, D.A. (2004). Cortical connections of the lateral medio-dorsal thalamus in Cynomolgus monkeys. The Journal of Comparative Neurology, 473, 107-127. [ Links ]
Ferry, A.T., Öngür, D., An, X. & Price, J.L. (2000). Prefrontal cortical projections to the striatum in macaque monkey: Evidence for an organization related to prefrontal networks. The Journal of Comparative Neurology, 425, 447-470. [ Links ]
Fletcher, P.C. & Henson, R.N. (2001). Frontal lobes and human memory. Insights from functional neuroimaging. Brain, 124, 849-881. [ Links ]
Fuster, J.M. (2000). Executive Frontal Functions. Experimental Brain Research, 133, 66-70. [ Links ]
Ghashghaei, H.T. & Barbas, H. (2001). Neural interaction between the basal forebrain and functionally distinct prefrontal cortices in the rhesus monkey. Neuroscience, 103, 3, 593-614. [ Links ]
Ghashghaei, H.T. & Barbas, H. (2002). Pathways for emotion: Interactions of prefrontal and anterior temporal pathways in the amygdala of the rhesus monkey. Neuroscience, 115, 1261-1279. [ Links ]
Gilbert, S.J., Frith, C.D. & Burgess, P.W. (2005). Involvement of rostral prefrontal cortex in selection between stimulus-oriented and stimulus-independent thought. European Journal of Neuroscience, 21, 1423-1431. [ Links ]
Hof, P.R., Mufson, E.J. & Morrison, J.H. (1995). Human orbitofrontal cortex: cytoarchitecture and quantitative immunohistochemical parcellation. The Journal of Comparative Neurology, 359, 48-68. [ Links ]
Jacobs, B., Schall, M., Prather, M., Kapler, E., Driscoll, L., Baca, S., Jacobs, J., Ford, K., Wainwright, M. & Treml, M. (2001). Regional dendritic and spine variation in human cerebral cortex: A quantitative Golgi study. Cerebral Cortex, 11, 558-571. [ Links ]
Johnson, S.C., Baxter, L.C., Wilder, L.S., Pipe, J.G., Heiserman, J.E. & Prigatano, G.P. (2002). Neural correlates of self-reflection. Brain, 125, 1808-1814. [ Links ]
Kaas, H.J. (1995). Principles of organization of sensory perceptual systems in mammals. Progress in Psychobiology and Physiological Psychology, 9, 1-41. [ Links ]
Koechlin, E., Basso, G., Pietrini, P., Panzer, S. & Grafman, J. (1999). The role of the anterior prefrontal cortex in cognition. Nature, 399, 148-151. [ Links ]
Koechlin, E., Corrado, G., Pietrini, P. & Grafman, J. (2000). Dissociating the role of the medial and lateral anterior prefrontal cortex in human planning. Proceedings of the National Academy of Sciences, USA, 97, 13, 7651-7656. [ Links ]
Koechlin, E., Ody, C. & Kouneiher, F. (2003). The architecture of cognitive control in the human prefrontal cortex. Science, 302, 1181-1185. [ Links ]
Kondo, H., Saleem, K.S. & Price, J.L. (2003). Differential connections of the temporal pole with the orbital and medial prefrontal networks in macaque monkey. The Journal of Comparative Neurology, 465, 499-523. [ Links ]
Kupfermann, I. S. (1991). Localization of higher cognitive and affective functions: The association cortices. En E. Kandel, J. Schwartz y T. Jessell (Eds), Principles of Neural Science (pp. 823 - 883). Nueva York: Appleton & Lange. [ Links ]
Markram, H., ToledoRodríguez, M., Wang, Y., Gupta, A., Silberberg, G. & Wu, C. (2004). Interneurons of the neocortical inhibitory system. Nature Reviews Neuroscience, 5, 793-807. [ Links ]
Middleton, F.A. & Strick, P.L. (2001). Cerebellar projections to the prefrontal cortex of the primate. The Journal of Neuroscience, 21, 700-712. [ Links ]
Morecraft, R.J. & Van Hoesen, G.W. (1993). Frontal granular cortex input to the cingulate (M3), supplementary (M2) and primary (M1) motor cortices in the rhesus monkey. The Journal of Comparative Neurology, 337, 669-689. [ Links ]
Örgür, D., Ferry, A.T. & Price, J.L. (2003). Architectonic subdivision of the human orbital and medial prefrontal cortex. The Journal of Comparative Neurology, 460, 425-449. [ Links ]
Örgür, D. & Price, J.L. (2000). The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cerebral Cortex, 10, 206-219. [ Links ]
Pandya, D. & Yeterian, E. (1990). Prefrontal cortex in relation to other cortical areas in Rhesus monkey: Architecture and connections. Progress in Brain Research, 85, 63-94. [ Links ]
Perlaza, O.I., Buriticá, E., Umbarila, J., Arteaga, G. & Pimienta, H.J. (2005). Corteza prefrontal humana: Áreas 10m, 11o y 46d. Citoarquitectura e implicaciones funcionales. Revista Salud UIS, 37, 6-14. [ Links ]
Petrides, M. & Pandya, D.N. (1999). Dorsolateral prefrontal cortex: Comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. European Journal of Neuroscience, 16, 291-310. [ Links ]
Pimienta, H.J., Escobar, M.I., Palomino, J.C. & Quijano, M.C. (1999). Corteza prefrontal: un mosaico evolutivo, estructural, funcional y clínico. En J. Muñoz y C. Serrano (Eds), Primates, evolución e identidad humana (pp. 73 - 82). México: Publicaciones del Instituto Mexicano de Psiquiatría. [ Links ]
Ramnani, N. & Owen, A.M. (2004). Anterior prefrontal cortex: Insights into function from anatomy and neuroimaging. Nature, 5, 184-194. [ Links ]
Ranganath, C., Johnson, M.K. & D'esposito, M. (2003). Prefrontal activity associated with working memory and episodic longterm memory. Neuropsychologia, 41, 378-389. [ Links ]
Rempel-Clower, N.L. & Barbas, H. (1998). Topographic organization of connections between the hypothalamus and prefrontal cortex in the rhesus monkey. The Journal of Comparative Neurology, 398, 393-419. [ Links ]
Rempel-Clower, N.L. & Barbas, H. (2000). The laminar pattern of connections between prefrontal and anterior temporal cortices in the rhesus monkey is related to cortical structure and function. Cerebral Cortex, 10, 851-865. [ Links ]
Romanski, L.M.; Giguere, M.; Bates, J.F. & Goldman-Rakic, P.S. (1997). Topographic organization of medial pulvinar connections with the prefrontal cortex in the rhesus monkey. The Journal of Comparative Neurology, 379, 313-332. [ Links ]
Sanides, F. (1969). Comparative architectonics of the neocortex of mammals and their evolutionary interpretation. En M.I. Escobar, J.C. Palomino, M. Arévalo y H.J. Pimienta (1998). Dorsolateral prefrontal cortex of the Owl monkey: A qualitative and quantitative Nissl and GABA immunostaining study. Alzheimer's Disease Review, 3, 57-62. [ Links ]
Schmahmann, J.D. & Pandya, D.N. (1997). Anatomic organization of the basilar pontine projections from prefrontal cortices in rhesus monkey.The Journal of Neuroscience, 17, 438-458. [ Links ]
Seltzer, B. & Pandya, D.N. (1978). Afferent cortical connections and architectonics of the superior temporal sulcus and surrounding cortex in the rhesus monkey. Brain Research, 149, 1-24. [ Links ]
Semendeferi, K., Armstrong, E., Schleicher, A., Zilles, K. & Van Hoesen, G.W. (2001). Prefrontal cortex in humans and apes: A comparative study of area 10. American Journal of Physical Anthropology, 114, 224-241. [ Links ]
Van Eden, C.G. & Buijs R.M. (2000). Functional Neuroanatomy of the Prefrontal Cortex: autonomic Interactions. Progress in Brain Research, 126, 49-62. [ Links ]
Walker, Ae. (1940). A cytoarchitectural study of the prefrontal area of the macaque monkey. The Journal of Comparative Neurology, 73, 59-86. [ Links ]
Xiao, D. & Barbas, H. (2002). Pathways for emotions and memory I. Input and output zones linking the anterior thalamic nuclei with prefrontal cortices in the rhesus monkey.Thalamus & Related Systems, 2, 21-32. [ Links ]
Yeterian, E. & Pandya, D. (1991). Prefrontostriatal connections in relation to cortical arquitectonic oraganization in rhesus monkey. Journal of Comparative Neurology, 312, 43-67. [ Links ]
Recepción: Diciembre de 2005
Aceptación final: Agosto de 2006
1 Correspondencia: Hernán José Pimienta-Jiménez, Centro de Estudios Cerebrales, Facultad de Salud, Universidad del Valle, Cali, Colombia. Correo electrónico: hernpim@telesat.com.co.
2 Los autores desean expresar su agradecimiento al Instituto Nacional de Medicina Legal y Ciencias Forenses Regional Bogotá por el suministro de los especímenes. A la Vicerrectoría de Investigaciones de la Universidad del Valle por el apoyo financiero. A Colciencias por el apoyo a través del programa de Jóvenes Investigadores. A la Dra. Martha Isabel Escobar por el apoyo en los protocolos de inmunohistoquímica y por la revisión crítica del manuscrito. A los Dres. Liliana Villamil, John Umbarila, Gabriel Arteaga y Óscar Perlaza por su apoyo técnico.
