










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkCadernos de Pós-Graduação em Distúrbios do Desenvolvimento
versão impressa ISSN 1519-0307versão On-line ISSN 1809-4139
Cad. Pós-Grad. Distúrb. Desenvolv. vol.20 no.1 São Paulo jan./jun. 2020
https://doi.org/10.5935/cadernosdisturbios.v20n1p176-191
O ambiente materno influencia o desenvolvimento ponderal e a viabilidade da prole de ratos, porém não altera o cuidado maternal
The maternal environment influences the ponderal development and the viability of the offspring of rats but does not alter maternal care
El ambiente materno influye en el desarrollo ponderal y la viabilidad de la descendencia de las ratas, pero no altera el cuidado materno
Loren da Silva MedeirosI; Paula Da Silva RodriguesII; Ana Claudia Silva SampaioIII; Maria Martha BernardiIV
IUniversidade Paulista (Unip), São Paulo, SP, Brasil / E-mail: loren.medeiros.s@gmail.com
IIUniversidade Paulista (Unip), São Paulo, SP, Brasil / E-mail: paulaa.rodrigues@live.com
IIIUniversidade Paulista (Unip), São Paulo, SP, Brasil / E-mail: acssampaio.91@gmail.com
IVUniversidade Paulista (Unip), São Paulo, SP, Brasil / E-mail: maria.bernardi@docente.unip.br
RESUMO
O comportamento materno tem como objetivo garantir a sobrevivência e o desenvolvimento adequado da ninhada. Variações no ambiente ou no número de filhotes durante o período perinatal podem levar a alterações no desenvolvimento da prole. Este trabalho avaliou a influência de um ambiente materno desfavorável e da redução da ninhada no cuidado maternal, na viabilidade e no ganho de peso dos filhotes. Ratas lactantes foram divididas em três grupos: C - grupo controle mantido em gaiolas com 100% da maravalha e ninhadas padronizadas em oito filhotes; E1 - grupo experimental 1 em que as ninhadas foram padronizadas em quatro filhotes mantendo-se 100% da maravalha da gaiola; E2 - grupo experimental 2 mantido em gaiolas com redução de 60% na quantidade de maravalha e ninhadas padronizadas em oito filhotes. Entre os dias 5 e 7 da lactação, o comportamento materno foi observado. Avaliaram-se o ganho de peso das proles nos dias 4, 10 e 21 da lactação e a sobrevivência delas ao desmame. Em comparação ao grupo C, verificou-se o seguinte: 1. o cuidado maternal destinado aos filhotes não foi modificado; 2. as mães do grupo E1 apresentaram maior tempo de auto-grooming; 3. as mães do grupo E2 apresentaram maior frequência, tempo total e tempo máximo de auto-grooming; 4. a prole do grupo E1 apresentou maior ganho de peso; 5. a prole do grupo E2 não apresentou alterações no ganho de peso, porém houve menor sobrevivência. A redução da ninhada promoveu maior ganho de peso da prole, enquanto a menor disponibilidade de maravalha não influenciou o ganho de peso da prole. Em ambas as condições, o cuidado maternal não foi modificado. O aumento do auto-grooming materno no grupo E2 pode ser interpretado como um sinal de estresse materno e no grupo E1 como sinal de conforto.
Palavras-chave: Comportamento. Ganho de peso. Comportamento materno. Asseio animal. Atividade motora.
ABSTRACT
Maternal behavior aims to ensure the survival and development of the offspring and variations in the environment or the number of pups can lead to changes in the development of the offspring. This study evaluated the influence of an unfavorable maternal environment and the reduction of the litter in maternal care and in the offspring viability and weight gain of offspring. Lactating rats were divided into three groups: C - control group kept in cages with 100% shavings and standardized litters in 8 pups; E1 - experimental group 1 in which litters were standardized in 4 puppies keeping 100% of the shavings in the cage; E2 - experimental group 2 kept in cages with a 60% reduction in the amount of wood shavings and 8 hatchlings. Between days 5-7 of lactation, general activity and maternal behavior were observed. The weight gain of the offspring was evaluated on days 4, 10 and 21 of lactation. In relation to group C, it was found that: 1. the maternal care of the puppies was not modified; 2. mothers in group E1 had a longer auto-grooming time; 3. mothers in the E2 group had higher frequency, total time and maximum auto-grooming time; 4. the offspring of the E1 group showed greater weight gain; 5. the offspring of the E2 group showed no changes in weight gain but an increased mortality. The reduction of the litter promoted greater weight gain of the offspring while the lower availability of shavings did not influence the weight gain of the offspring. In both conditions, maternal care was not modified. The increase in maternal auto-grooming in the E2 group can be interpreted as a sign of maternal stress and only the increased time of the auto-grooming in the E1 group suggest a reduce response to stress.
Keywords: Behavior. Weight gain. Maternal behavior. Animal cleanliness. Motor activity.
RESUMEN
El comportamiento materno tiene como objetivo garantizar la supervivencia y el desarrollo de la descendencia y las variaciones en el entorno o el número de crías pueden conducir a cambios en el desarrollo de la descendencia. Este trabajo evaluó la influencia de un entorno materno desfavorable y la reducción de la prole en el cuidado materno, en la viabilidad y el aumento de peso de la descendencia. Las ratas lactantes se dividieron en tres grupos: C- grupo de control mantenido en jaulas con aserrín de madera 100% y proles estandarizadas en 8 crías; E1 - grupo experimental 1 en el que las proles se estandarizaron en 4 crías manteniendo el 100% de las aserrín de madera en la jaula; E2 - grupo experimental 2 mantenido en jaulas con una reducción del 60% en la cantidad de aserrín de madera y 8 neonatos. Entre los días 5-7 de lactancia, se observó actividad general y comportamiento materno. El aumento de peso de la descendencia se evaluó los días 4, 10 y 21 de lactancia y su viabilidad al destete. En relación con el grupo C, se encontró que: 1. el cuidado materno de las proles no fue modificados; 2. las madres en el grupo E1 tuvieron un tiempo de aseo más largo; 3. las madres en el grupo E2 tuvieron mayor frecuencia, tiempo total y tiempo máximo de preparación; 4. la descendencia del grupo E1 mostró mayor aumento de peso; 5. la descendencia del grupo E2 no mostró cambios en el aumento de peso pero presentó reducción en su viabilidad. La reducción de la camada promovió un mayor aumento de peso de la descendencia, mientras que la menor disponibilidad de virutas no influyó en el aumento de peso de la descendencia. En ambas condiciones, la atención materna no fue modificada. El aumento de la preparación materna en el grupo E2 puede interpretarse como un signo de estrés materno y en el grupo E1 sugiere una respuesta reducida al estrés.
Palabras clave: Comportamiento. Aumento de peso. Comportamiento maternal. Limpieza animal. Actividad motora.
INTRODUÇÃO
O comportamento materno (CM) consiste em uma série de cuidados que os membros adultos de uma determinada espécie realizam em torno dos indivíduos reprodutivamente imaturos, para auxiliar na propagação de sua espécie. O controle do CM envolve fatores neuroendócrinos e sensoriais (CAMERON et al., 2008). Durante esse período, a meta principal da mãe é garantir a sobrevivência dela e de seus filhotes. Assim, os hormônios gestacionais preparam o animal para agir de forma maternal para com os filhotes, enquanto os neurotransmissores regulam o CM durante a fase de manutenção e lactação (NUMAN, 1988; KINSLEY; AMORY-MEYER, 2011; ROBINSON; ZITZMAN; WILLIAMS, 2011; TEODOROV; FELÍCIO; BERNARDI, 2010).
Em ratas, antes do nascimento dos filhotes, as mães constroem ninhos para promover a termorregulação de sua prole, já que ao nascerem, os filhotes são incapazes de controlar sua temperatura corpórea (HENNESSY et al., 1980). No ninho, as fêmeas lactantes podem ficar sobre os filhotes numa postura denominada crouching ou cifose fisiológica, na qual sua coluna fica arqueada, facilitando a amamentação, além de passarem um tempo significativo lambendo a cria para limpá-la, já que a limpeza da área anogenital estimula a defecação e micção dos filhotes, assim como favorece a diferenciação sexual do cérebro da prole masculina (DÖHLER, 1991).
Diversos fatores ambientais, psicológicos, biológicos e neurais interagem para promover e manter o CM. Assim como mudanças hormonais internas geram respostas fisiológicas e comportamentais compensatórias, mudanças ambientais também levam a alterações hormonais (SCHULKIN, 1999; TEODOROV; FELÍCIO; BERNARDI, 2010).
Variações no CM causadas por diversas condições relacionadas à mãe ou ao ambiente materno são associadas a distúrbios neurocomportamentais tardios, que podem ser observados no início e ao longo da vida de um determinado indivíduo. Modelos como o estresse materno e a separação maternal mostraram que maus-tratos na primeira infância aumentam o risco para deficiências comportamentais (HEYSER, 2003), imunes (NGUYEN; ARMSTRONG; YAGER, 2017) e distúrbios neurológicos (KINSLEY et al., 1999; OLAZÁBAL et al., 2013) em longo prazo, podendo resultar em comportamento social alterado (FORD; FRALEIGH; CONNOR, 2010) e distúrbios na saúde (BIFULCO et al., 2002). Estima-se que, nas sociedades ocidentais, as deficiências provocadas pela redução do cuidado maternal na infância variam de 10% a 15% (GUSTAFSON; SARWER, 2004). Em humanos, podem-se citar, como deficiências do cuidado na infância, os abusos físico, sexual e psicológico, a negligência e a existência de ambientes não apropriados para o desenvolvimento. Devem-se considerar ainda as instituições para crianças abandonadas ou em risco (FORD; FRALEIGH; CONNOR, 2010).
Modelos animais podem ser úteis para entender a influência do cuidado materno no desenvolvimento da prole. Nesse sentido, um paradigma naturalístico para induzir estresse materno e redução no cuidado maternal é a limitação da maravalha da gaiola, como descrito por Yan et al. (2017). Nessa condição, pode ocorrer redução da capacidade das mães em construir o ninho, necessidade de construção de ninhos mais frequentemente, menor tempo de contato com os filhotes, manuseio brusco e pisoteamento dos filhotes, reduzindo assim o cuidado maternal.
Contudo, o modelo em que se reduz o tamanho da ninhada é classicamente empregado como modelo de obesidade infantil. Nesse caso, Carvalho et al. (2016)litters were adjusted to a small litter (SL mostraram que fêmeas lactantes de ninhadas reduzidas cuidaram melhor de sua prole que aquelas do grupo controle. Portanto, este trabalho teve como objetivo comparar o CM de ratas nessas duas condições, bem como a influência desses fatores na viabilidade e no desenvolvimento ponderal de sua prole.
MÉTODOS
Animais
Empregaram-se 16 ratas e 15 ratos Wistar no início dos experimentos, entre 15 e 17 semanas de idade, pesando 220-275 g, oriundos do Departamento de Imunologia, Instituto de Ciências Biomédicas da Universidade de São Paulo. Esses animais foram trazidos ao Biotério de Experimentação da Universidade Paulista, em veículo com ar-condicionado, certificados quanto à saúde pelo respectivo biotério e alojados em gaiolas de polipropileno (45,5 x 34,5 x 20 cm; máximo dois animais/gaiola), com sistema de microisoladores (Tecniplast, Buguggiate, Italy), em temperatura controlada (23 ºC ± 2 ºC) e umidade (55%-65%), com iluminação artificial (12 horas luz/12 horas escuro, luz ligada às 7 horas). Os animais tiveram acesso, sem nenhum tipo de limitação, à ração para roedores irradiada (Bio Base, Águas Frias, Brazil) e água filtrada. Maravalha esterilizada e livre de resíduos foi usada como cama para os animais. Após dez dias de habituação, as fêmeas foram cruzadas com ratos de mesma linhagem, experientes sexualmente, para obtenção de suas proles. Após 18 dias da confirmação da gestação pela observação da presença de espermatozoides no lavado vaginal, as ratas foram alojadas individualmente, com comida e água ad libitum. Permitiu-se que a gestação fosse a termo. O parto foi considerado como dia 1 pós-natal (DPN 1), e não se realizou nenhum manuseio nesse dia.
Formação dos grupos
No DPN 2, pesaram-se e padronizaram-se as ninhadas, e as 16 ratas foram divididas em três grupos: E1 (n = 4), E2 (n = 6) e controle (n = 6).
• Grupo C: as ratas do grupo controle ficaram em gaiolas com 100% da maravalha, e as ninhadas foram padronizadas em quatro machos/quatro fêmeas com a mãe.
• Grupo E1: as ratas do grupo experimental 1 tiveram as ninhadas padronizadas em três filhotes machos e uma fêmea, mantendo-se 100% da maravalha da gaiola.
• Grupo E2: as ratas do grupo experimental 2 tiveram redução de 60% na quantidade de maravalha de sua gaiola, mantendo-se quatro filhotes fêmeas e quatro filhotes machos com a mãe.
Comportamento materno
Para o teste de CM, as mães foram isoladas de seus filhotes em uma gaiola com maravalha limpa, em uma sala diferente dos filhotes, por 30 minutos. Sua prole foi mantida aquecida em outra gaiola por uma manta térmica. Antes do início das observações, a caixa moradia teve seu ninho desfeito. A seguir, os filhotes foram distribuídos aleatoriamente pela gaiola moradia e a fêmea lactante foi introduzida em seguida. As observações do CM foram feitas por 30 minutos. Avaliaram-se os seguintes parâmetros: 1. latência para recolhimento dos filhotes em segundos, 2. tempo total e máximo da mãe com os filhotes em segundos, 3. frequência de auto-grooming materno, 4. tempo total e máximo de auto-grooming materno em segundos, 5. frequência de grooming nos filhotes; 6. tempo total e máximo de grooming nos filhotes em segundos.
Define-se auto-grooming como o ato de a mãe lamber o próprio corpo e grooming como o ato de lamber o filhote.
Avaliação do desenvolvimento ponderal da prole
Os filhotes machos foram pesados individualmente nos DPN 4, DPN 10 e DPN 21. Esse procedimento permitiu, ao final dos experimentos, calcular o ganho de peso.
Viabilidade da prole ao desmame
A sobrevivência da prole foi avaliada no DPN 21 (dia do desmame).
Análise estatística
A análise de variância de uma via (ANOVA), seguida pelo teste de comparações múltiplas de Dunnett, foi empregada para comparação dos dados de três grupos. A ANOVA de duas vias foi empregada, seguida pelo teste de Tuckey para análise de dois fatores. A probabilidade de 5% foi considerada capaz de indicar diferenças significantes entre os grupos.
RESULTADOS
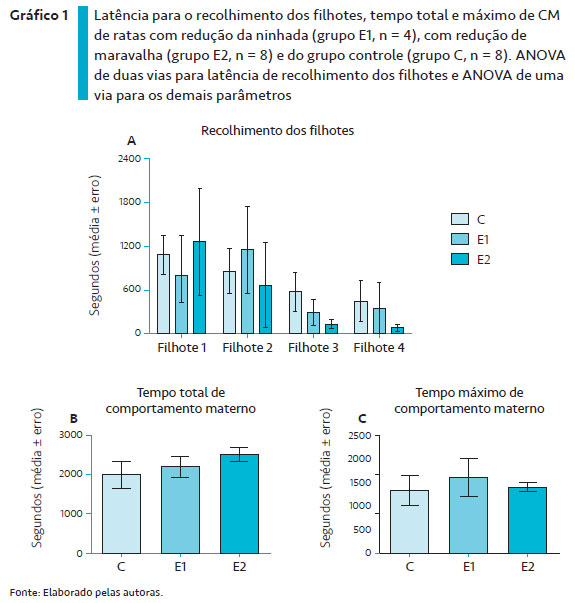
Os parâmetros do CM estão ilustrados no Gráfico 1. A latência para recolhimento de quatro filhotes (Gráfico 1A) não foi diferente com relação ao tipo de ambiente [F 2,52 = 0,31, p = 0,74] e entre a sequência de recolhimento dos filhotes [F3,52 = 2,72, p = 0,54] sem interação entre os fatores [F3,52 = 0,32, p = 0,93].
Quanto ao tempo total de CM (Gráfico 1B), a ANOVA de uma via não revelou diferenças entre os grupos em relação ao grupo controle [F (2, 12) = 0,8884; p = 0,4367]. Resultados similares foram observados para o tempo máximo de CM (Gráfico 1C) [F (2, 12) = 0,2105; p = 0,8131].
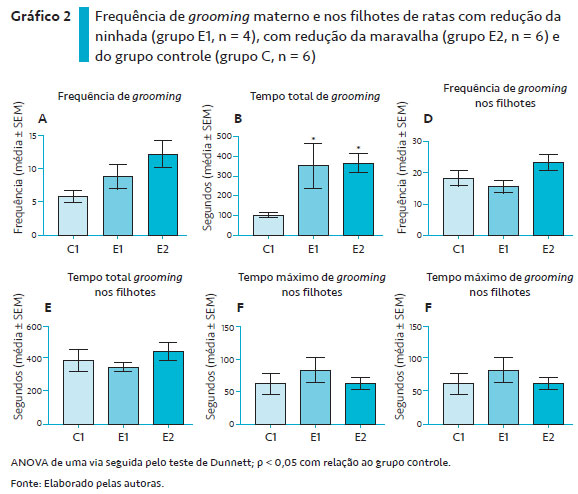
O Gráfico 2 ilustra os parâmetros de auto-grooming materno e dirigido aos filhotes das ratas dos grupos E1 e E2.
Com relação à frequência de auto-grooming materno (Gráfico 2A), a ANOVA de uma via não revelou diferenças entre os grupos E1 e E2 em relação ao grupo controle [F (2, 12) = 3,745; p = 0,0545].
O tempo total de auto-grooming materno (Gráfico 2B) foi diferente entre os grupos [F (2, 12) = 5,767; p = 0,0176]. O teste post hoc de Dunnett revelou que os grupos E1 e E2 apresentaram um tempo maior de auto-grooming materno em relação ao grupo controle (*p < 0,5).
No tempo máximo de auto-grooming (Gráfico 2C), a ANOVA de uma via revelou diferenças significantes entre os grupos [F (2, 11) = 4,145; p = 0,0455]. O teste post hoc de Dunnett revelou que o grupo E2 apresentou maior tempo máximo de auto-grooming que o grupo controle (*p < 0,5). Não houve diferença entre os grupos dos animais do grupo E1 e do grupo controle.
Com relação ao grooming dos filhotes, a ANOVA de uma via não indicou a existência de diferenças significantes entre os grupos na frequência de grooming dos filhotes (Gráfico 2D) [F (2, 13) = 2,598; p = 0,1124], no tempo total de grooming nos filhotes (Gráfico 2E) [F (2, 13) = 0,6464; p = 0,5400] e no tempo máximo de grooming nos filhotes (Gráfico 2F) [F (2, 13) = 0,5337; p = 0,5988].
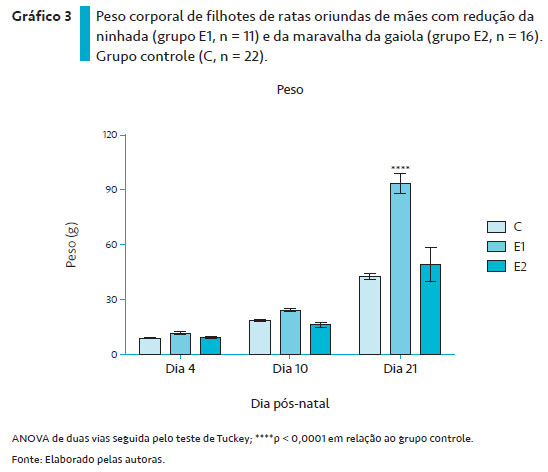
O Gráfico 3 ilustra o desenvolvimento ponderal de filhotes das ratas dos grupos controle, E1 e E2. A ANOVA de duas vias mostrou diferenças significantes entre os dias de observação (F2,78 = 195,69, p < 0,0001) e tipos de manutenção das gaiolas dos animais (F 2,78 = 28,70, p < 0,0001), com interação entre os fatores (F4,78 = 16,65, p < 0,0001). O teste de Tuckey de comparações múltiplas indicou que no dia 21 os animais do grupo E1 apresentaram aumento significante em relação aos grupos controle (p < 0,0001) e E2, enquanto aqueles do grupo E2 não mostraram diferenças com relação ao grupo controle.
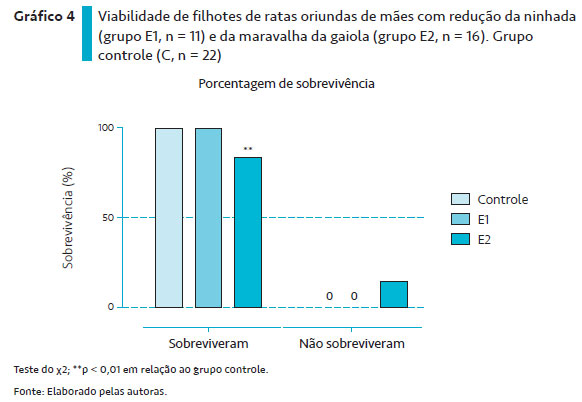
O Gráfico 4 ilustra o índice de sobrevivência da prole de ratos dos grupos C, E1 e E2. Nota-se que todos os animais das proles dos grupos C e E1 sobreviveram até o desmame. No entanto, as proles do grupo E2 apresentaram 85,4% de sobrevivência, o que diferiu estatisticamente daquelas do grupo C e E1 (p < 0,001).
DISCUSSÃO
Os resultados deste estudo demonstraram que variações no ambiente neonatal foram capazes de influenciar o desenvolvimento ponderal da prole de ratos, em particular aumentando o ganho de peso nas ninhadas com menor número de filhotes. Por sua vez, o índice de sobrevivência das proles com redução da maravalha do ninho foi menor que nos demais grupos.
O CM foi observado entre os dias 6 e 7 da lactação. Nesses dias, os filhotes mantiveram o CM pela estimulação da liberação de hormônios maternos que é feita no ato da amamentação (CHAMPAGNE; CURLEY, 2009; CHAMPAGNE et al., 2003). Os mamíferos não humanos geralmente apresentam um conjunto estereotipado de respostas comportamentais aos seus recém-nascidos. Eles exibem maneiras espécie-específicas de transporte, exploração, alimentação e higiene de seus filhotes, e de protegê-los de predadores e outros perigos (KINSLEY; AMORY-MEYER, 2011). Esses comportamentos promovem a resiliência fisiológica e imunológica, a maturação física e o desenvolvimento social e emocional típico das espécies nos jovens. Nos primeiros dez dias após o parto, as mães ficam muito tempo no ninho, e, à medida que os filhotes crescem e se tornam mais independentes, os cuidados maternos tendem a decrescer gradativamente. Como o CM é mantido em boa parte pela estimulação oriunda do filhote, a mãe se torna menos responsiva em relação a ele. Em ratos, o processo de desmame da prole inicia-se a partir do dia em que os filhotes começam a andar e entram em contato com a ração, o que ocorre a partir do dia 10 da lactação. Em torno do 20º dia da lactação, os filhotes se alimentam preferencialmente de ração (NUMAN; FLEMING; LEVY, 2006; NUMAN, 2007; TEODOROV; FELÍCIO; BERNARDI, 2010). No entanto, não se observaram diferenças no CM, evidenciando que tanto as alterações ambientais como a redução da ninhada não foram capazes de modificar o cuidado maternal.
Contudo, a mães dos dois grupos apresentaram aumento na expressão do comportamento de auto-grooming, também denominado autolimpeza.
O comportamento de grooming é um padrão fixo de ação que ocorre em várias espécies animais, incluindo artrópodes e aves (SPRUIJT; VAN HOOFF; GISPEN, 1992), vários mamíferos roedores (BERRIDGE, 1990) e felídeos (ECKSTEIN; HART, 2000). Sua função primária em roedores é ligada à higienização e a outros processos fisiológicos, como termorregulação, comunicação social e estimulação do organismo (KALUEFF et al., 2015).
Além disso, o grooming responde às variáveis ambientais estressoras. Spruijt, Van Hooff e Gispen (1992) estudaram o curso temporal do grooming em resposta a alguns estressores ambientais e observaram que a severidade do estressor pode retardar ou incrementar a ocorrência de grooming. O aumento observado no grooming materno pode então indicar que as alterações no ambiente materno e a redução da ninhada promoveram nas fêmeas aumento da ansiedade.
No entanto, essas alterações no auto-grooming materno não modificaram o grooming nos filhotes. Como parte do cuidado maternal, o grooming materno tem funções não só de higiene, mas também de estímulo da prole, em particular, a regulação do desenvolvimento dos sistemas neurais envolvidos com o estresse e a determinação sexual (MCCARTHY et al., 1997). Dessa forma, embora as variações no ambiente maternal tenham modificado o comportamento da mãe, os cuidados maternos não foram alterados.
Com relação ao desenvolvimento ponderal, nas proles do grupo E1 observou-se maior ganho de peso aos 21 dias de idade, enquanto aqueles do grupo E2 não mostraram diferenças nesse parâmetro com relação ao grupo controle.
O aumento do peso corporal da prole do grupo E1 pode ser atribuído ao fato de a ninhada ter sido reduzida, permitindo ao filhote maior aporte ao leite materno e, em consequência, maior ganho de peso. Resultados similares foram observados por Oliveira de Lira et al. (2014), em que a ninhada foi reduzida para dois filhotes. Nesse trabalho, os autores evidenciaram um efeito precoce da hiperalimentação nos períodos iniciais da vida sobre os parâmetros de crescimento somático e desenvolvimento sensório-motor em ratos. Estes autores propuseram que o excesso de alimentação nos períodos iniciais da vida pode ter promovido alterações metabólicas na regulação serotoninérgica do comportamento alimentar. Além disso, segundo Xavier et al. (2019). A redução da ninhada não só permite maior aporte do leite materno ao filhote, mas também altera qualitativamente a sua composição, levando a um leite com alto valor energético. Xavier et al. (2019) analisaram a composição do leite de ratas com redução da ninhada e verificaram que, em relação ao leite de ratas sem redução da ninhada, houve aumento no conteúdo de gordura e maiores níveis de triglicérides, sem modificações naqueles de glicose e proteínas. Dessa forma, embora o cuidado maternal não tenha sido modificado pela redução da ninhada, a interação materno-filhote por meio das modificações qualitativas e quantitativas do leite favoreceu o ganho de peso da prole.
Por sua vez, nas proles dos animais do grupo E2, não foram detectadas diferenças no ganho de peso corporal quando comparadas àquele do grupo controle, fato interessante, pois, nesse grupo, as condições ambientais não eram as ideais para o desenvolvimento da prole. No entanto, nota-se menor viabilidade da prole ao desmame, sugerindo que as condições do ninho tenham influenciado a sobrevida de alguns filhotes. Apesar de, na avaliação do CM, não haver diferenças entre os grupos controle e E2, verificou-se, da observação direta dos animais, que as mães com redução da maravalha cobriam os filhotes com toda a maravalha disponível, indicando uma tentativa de manter a temperatura da prole pela dificuldade de construir um ninho. Esse dado, associado ao aumento da frequência e tempo de auto-grooming, pode ser indicativo de estresse materno. Embora a avaliação do ganho de peso da prole dos animais do grupo E2 não tenha mostrado alterações, é fato conhecido que o estresse materno pode levar a efeitos deletérios na prole, em médio e longo prazo (LEVINE, 1967; TRACTENBERG et al., 2016). Nesse sentido, Moussaoui et al. (2016) mostraram que a redução das condições do ninho promove estresse materno por ativação do eixo hipófise-hipotálamo-adrenal e aumento dos níveis de glicocorticoides maternos.
CONCLUSÕES
Embora a avaliação do CM em condições de redução da ninhada e do ninho não tenha diferido, o ganho de peso das ninhadas foi diferente nas duas condições de manutenção da prole. Atribuiu-se o maior ganho de peso dos animais com redução da ninhada à melhor qualidade e disponibilidade de leite materno. No caso da redução do ninho, a prole mostrou ganho de peso similar ao do grupo controle, porém menor viabilidade, apesar de o cuidado maternal não diferir daquele do grupo controle.
REFERÊNCIAS
BERRIDGE, K. C. Comparative fine structure of action: rules of form and sequence in the grooming patterns of six rodent species. Behaviour, v. 113, n. 1-2, p. 21-56, 1990. [ Links ]
BIFULCO, A. et al. Exploring psychological abuse in childhood: II. Association with other abuse and adult clinical depression. Bulletin of the Menninger Clinic, v. 66, n. 3, p. 241-258, 2002. [ Links ]
CAMERON, N. M. et al. Epigenetic programming of phenotypic variations in reproductive strategies in the rat through maternal care. Journal of Neuroendocrinology, v. 20, n. 6, p. 795-801, 2008. [ Links ]
CARVALHO, A. L. O. et al. Early life overnutrition induced by litter size manipulation decreases social play behavior in adolescent male rats. International Journal of Developmental Neuroscience, v. 53, p. 75-82, 2016. [ Links ]
CHAMPAGNE, F. A.; CURLEY, J. P. Epigenetic mechanisms mediating the long-term effects of maternal care on development. Neuroscience & Biobehavioral Reviews, v. 33, n. 4, p. 593-600, 2009. [ Links ]
CHAMPAGNE, F. A. et al. Variations in maternal care in the rat as a mediating influence for the effects of environment on development. Physiology & Behavior, v. 79, n. 3, p. 359-371, 2003. [ Links ]
DÖHLER, K. D. The Pre- and postnatal influence of hormones and neurotransmitters on sexual differentiation of the mammalian hypothalamus. International Review of Cytology, v. 131, n. C, p. 1-57, 1991. [ Links ]
ECKSTEIN, R. A.; HART, B. L. Grooming and control of fleas in cats. Applied Animal Behaviour Science, v. 68, n. 2, p. 141-150, 2000. [ Links ]
FORD, J. D.; FRALEIGH, L. A.; CONNOR, D. F. Child abuse and aggression among seriously emotionally disturbed children. Journal of Clinical Child and Adolescent Psychology, v. 39, n. 1, p. 25-34, 2010. [ Links ]
GUSTAFSON, T. B.; SARWER, D. B. Childhood sexual abuse and obesity. Obesity Reviews, v. 5, n. 3, p. 129-135, 2004. [ Links ]
HENNESSY, M. B. et al. Maternal behavior, pup vocalizations, and pup temperature changes following handling in mice of 2 inbred strains. Developmental Psychobiology, v. 13, n. 6, p. 573-584, 1980. [ Links ]
HEYSER, C. J. Assessment of developmental milestones in rodents. Current Protocols in Neuroscience, v. 25, n. 1, p. 8.18- 8.18.15, 2003. [ Links ]
KALUEFF, A. V. et al. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nature Reviews Neuroscience, v. 17, n. 1, p. 45-59, 2015. [ Links ]
KINSLEY, C. H.; AMORY-MEYER, E. Why the maternal brain? Journal of Neuroendocrinology, v. 23, n. 11, p. 974-983, 2011. [ Links ]
KINSLEY, C. H. et al. Motherhood improves learning and memory. Nature, v. 402, n. 6758, p. 137-138, 1999. [ Links ]
LAPORTA, J. et al. Serotonin (5-HT) affects expression of liver metabolic enzymes and mammary gland glucose transporters during the transition from pregnancy to lactation. PLoS ONE, v. 8, n. 2, p. e57847, 2013. [ Links ]
LEVINE, S. Maternal and environmental influences on the adrenocortical response to stress in weanling rats. Science, v. 156, n. 3772, p. 258-260, 1967. [ Links ]
MCCARTHY, M. M. et al. Influence of maternal grooming, sex and age on fos immunoreactivity in the preoptic area of neonatal rats: implications for sexual differentiation. Developmental Neuroscience, v. 19, n. 6, p. 488-496, 1997. [ Links ]
MOUSSAOUI, N. et al. Limited nesting stress alters maternal behavior and in vivo intestinal permeability in male Wistar pup rats. PLoS ONE, v. 11, n. 5, p. e0155037, 2016. [ Links ]
NGUYEN, A. T.; ARMSTRONG, E. A.; YAGER, J. Y. Neurodevelopmental reflex testing in neonatal rat pups. Journal of Visualized Experiments, n. 122, p. 1-7, 2017. [ Links ]
NUMAN, M. Neural basis of maternal behavior in the rat. Psychoneuroendocrinology, v. 13, n. 1-2, p. 47-62, 1988. [ Links ]
NUMAN, M. Motivational systems and the neural circuitry of maternal behavior in the rat. Developmental Psychobiology, v. 49, n. 1, p. 12-21, 2007. [ Links ]
NUMAN, M.; FLEMING, A.; LEVY, F. Maternal behavior. In: Knobill, E. (ed.). Physiology of reproduction. New York: Raven Press, 2006. p. 1921-1993. [ Links ]
OLAZÁBAL, D. E. et al. New theoretical and experimental approaches on maternal motivation in mammals. Neuroscience & Biobehavioral Reviews, v. 37, n. 8, p. 1860-1874, 2013. [ Links ]
OLIVEIRA DE LIRA, L. et al. Overnutrition in lactation: effects on somatic and sensorimotor development in rats. Neurobiologia, v. 77, p. 1-16, 2014. [ Links ]
ROBINSON, D. L.; ZITZMAN, D. L.; WILLIAMS, S. K. Mesolimbic dopamine transients in motivated behaviors: focus on maternal behavior. Frontiers in Psychiatry, v. 2, p. 23, 2011. [ Links ]
SCHULKIN, J. Corticotropin-releasing hormone signals adversity in both the placenta and the brain: regulation by glucocorticoids and allostatic overload. Journal of Endocrinology, v. 161, n. 3, p. 349-356, 1999. [ Links ]
SPRUIJT, B. M.; VAN HOOFF, J. A. R. A. M.; GISPEN, W. H. Ethology and neurobiology of grooming behavior. Physiological Review, v. 72, n. 3, p. 825-852, 1992. [ Links ]
TEODOROV, E.; FELÍCIO, L. F.; BERNARDI, M. M. Maternal behavior. In: ANDERSEN, M.; TUFICK, S. (ed.). Animal models as ethical tools in biomedical research. São Paulo: CLR Balieiro Editores, 2010. v. I, p. 149-162. [ Links ]
TRACTENBERG, S. G. et al. An overview of maternal separation effects on behavioural outcomes in mice: evidence from a four-stage methodological systematic review. Neuroscience & Biobehavioral Reviews, v. 68, p. 489-503, 2016. [ Links ]
XAVIER, J. L. P. et al. Litter size reduction induces metabolic and histological adjustments in dams throughout lactation with early effects on offspring. Anais da Academia Brasileira de Ciências, v. 91, n. 1, p. e20170971, 2019. [ Links ]
YAN, C.-G. et al. Aberrant development of intrinsic brain activity in a rat model of caregiver maltreatment of offspring. Translational Psychiatry, v. 7, n. 1, p. e1005, 2017. [ Links ]
Recebido em: 10.2.2020
Aprovado em: 23.4.2020


{kind=link}
{kind=link}
{kind=link}
{kind=link}