










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkActa Comportamentalia
versão impressa ISSN 0188-8145
Acta comport. vol.18 no.3 Guadalajara 2010
ARTÍCULOS
Respostas de observação em pombos expostos a apresentações de comida independente de resposta1,2
Observing responses in pigeons exposed to response-independent food presentations
Gerson Yukio Tomanari3
Universidade de São Paulo
RESUMO
Pesquisas sobre respostas de observação convivem com divergências em relação ao estímulo discriminativo de menor probabilidade de reforçamento (S-), o qual, por vezes, pode ou não demonstrar funções reforçadoras condicionadas. Historicamente, estas divergências levaram autores a sugerir a existência de possíveis especificidades entre espécies quanto ao valor reforçador condicionado de S-, bem como a buscar modelos teóricos de reforçamento condicionado. Trata-se, sem dúvida, de uma área de pesquisa que acumula uma produção volumosa ao longo de mais de meio século, porém é marcadamente assistemática, especialmente em seus aspectos metodológicos. Isso dificulta, ou mesmo impede, comparações controladas entre resultados. Por essa razão, o presente trabalho reúne um conjunto de experimentos coletados em um processo sistemático de investigação em que pombos foram expostos ao procedimento em que comida era liberada independentemente do responder e respostas de observação produziam estímulos discriminativos de presença ou ausência de alimento. A partir dos resultados relatados, não se mantém a hipótese acerca das diferenças entre espécies. Além disso, os achados não sustentam os modelos teóricos vigentes, ao menos quando tomados isoladamente. Aparentemente, é importante que se faça uma avaliação mais ampla das variáveis envolvidas nas respostas de observação que, eventualmente, venha até mesmo a integrar diferentes aspectos de posições teóricas distintas.
Palavras chave: Resposta de observação; reforçamento condicionado; modelo da redução da incerteza; modelo da redução do atraso.
ABSTRACT
Research in observing behavior has faced divergences regarding the discriminative stimulus of lower probability of reinforcement (S-), which may or may not function as conditioned reinforcers. Historically, these divergences led some authors to suggest possible species-specific aspects of a conditioned reinforcement by S-, as well as to raise theoretical models of conditioned reinforcement. No doubt, it is a research area that has accumulated an expressive amount of data along more than half a century; however, such a production has been markedly non-systematic, especially in its methodological aspects. This fact makes it difficult, or even prevents, controlled comparisons among findings. For this reason, the present paper reviews a set of experiments carried out with pigeons exposed to a procedure in which food was delivered independently of responses and observing responses produced stimuli correlated with either the presence or the absence of food. Based on the results, the species-specific hypotheses could not be maintained. Besides, the findings do not support the current models of conditioned reinforcement, at least in their isolated forms. It seems important that variables related to the observing responses be analyzed in a broader assessment that could eventually integrate different aspects of distinct theoretical positions.
Key words: Observing response; conditioned reinforcement; uncertainty-reduction model; delay-reduction model.
Em 1952, Wyckoff propôs analisar como respostas que permitem a orientação do organismo a estímulos são adquiridas ao longo do estabelecimento de uma discriminação. O autor demonstrou originalmente que, dada a oportunidade, pombos emitem respostas cujas conseqüências são a produção de estímulos discriminativos. Essas respostas foram denominadas de respostas de observação (Wyckoff, 1952, 1969).
Desde o trabalho pioneiro de Wyckoff (1952, 1969), a necessidade da observação dos estímulos como requisito da discriminação tem-se consolidado à luz de um processo operante. Portanto, tanto o sucesso quanto o fracasso de um dado desempenho discriminado podem atrelar-se às respostas de observação controladas pelos estímulos envolvidos, identificados e compreendidos a partir da análise de seleção pelas conseqüências (Skinner, 1981). Portanto, uma análise mais precisa e eficiente de um operante discriminado poderia expandir-se para além dos termos da contingência relacionada com a produção do reforçador principal (estímulo discriminativo – resposta - estímulo reforçador) e incorporar os termos da contingência de observação (resposta de observação – estímulo discriminativo). A Figura 1 ilustra as duas contingências interligadas por meio do estímulo discriminativo, o qual atua como conseqüência das respostas de observação e como estímulo antecedente da resposta principal. Assim conduzida a análise, as possibilidades de compreensão da aquisição e da manutenção de operantes discriminados têm-se enriquecido enormemente (Dinsmoor, 1983; Shahan, 2002; Shahan, Magee, & Dobberstein, 2003; Shahan & Podlesnik, 2005; Tomanari, 2009).

Ao longo da história de investigação na área, esforços têm sido despendidos na compreensão acerca das funções possivelmente reforçadoras dos estímulos discriminativos que mantêm as respostas de observação. A literatura, por meio de uma variedade de métodos, acumula um conjunto substancial de resultados que atestam sistematicamente que um estímulo que acompanha uma maior probabilidade de reforçamento (S+) mantém respostas de observação. Com relação a esse achado, não há contestações. Entretanto, ao se tomar o estímulo que acompanha uma menor probabilidade de reforçamento (S-), dados experimentais divergem entre si. Por um lado, S- parece não apenas não contribuir para manter respostas que o produzem como, até mesmo, tendem a manter respostas que evitam sua apresentação (Allen & Lattal, 1989; Case & Fantino, 1981, 1989; Case, Fantino, & Wixted, 1985; Case, Ploog, & Fantino, 1990; Dinsmoor, Bowe, Green, & Hanson, 1988; Dinsmoor, Mulvaney, & Jwaideh, 1981; Fantino & Case, 1983; Fantino, Case, & Altus, 1983; Jenkins & Boakes, 1973; Jwaideh & Mulvaney, 1976; Mueller & Dinsmoor, 1984, 1986; Mulvaney, Dinsmoor, Jwaideh, & Hughes, 1974; Preston, 1985; Roper & Zentall, 1999; Schaal, Odun, & Shahan, 2000; Tomanari, 2001a,b, 2004; Tomanari, Machado, & Dube, 1998). Por outro, em contraposição a este conjunto majoritário de dados, foram verificadas condições em que S- pode, sim, manter respostas de observação (Lieberman, 1972; Lieberman, Cathro, Nichol, & Watson, 1997; Madden & Perone, 1999; Perone & Baron, 1980; Shrier, Thompson, & Spector, 1980; Tomanari, 1997, Experimento 2). Dados nesse sentido levam autores a sugerir a possibilidade de que um estímulo discriminativo de baixa ou nula probabilidade de reforçamento possa exercer funções reforçadoras, particularmente em primatas (Lieberman, 1972; Lieberman et al., 1997; Perone & Baron, 1980; Shrier et al., 1980).
Atualmente, a área conta com uma variedade metodológica que, acompanhada por um amplo espectro de resultados, dificulta a síntese de seu estado atual frente a suas questões fundamentais. Tipicamente, os estudos em resposta de observação empregam procedimentos cuja apresentação do reforçador principal (água, comida, pontos ou fichas, por exemplo) depende da emissão de respostas (respostas principais) que concorrem com as respostas de observação e, portanto, competem entre si (DeFulio & Hackenberg, 2008). Assim, por um lado, conseguem-se medidas de discriminação a partir da análise do responder na presença dos estímulos discriminativos. Por outro, na medida em que as respostas mantidas pelo reforçador principal concorrem com as respostas de observação, a análise dessas últimas deve, necessariamente, incorporar a simultaneidade das contingências relacionadas com a produção do reforçador principal. Nesse caso, a complexidade da situação tende a dificultar a análise dos fatores que contribuem (ou dificultam) a manutenção de respostas de observação.
Uma forma de minimizar as interações entre as respostas conseqüenciadas por comida e as respostas conseqüenciadas pela apresentação dos estímulos discriminativos se dá pelo uso de um procedimento cuja liberação de água, comida, pontos, por exemplo, independe da emissão de resposta. A Figura 2 esquematiza esse procedimento.
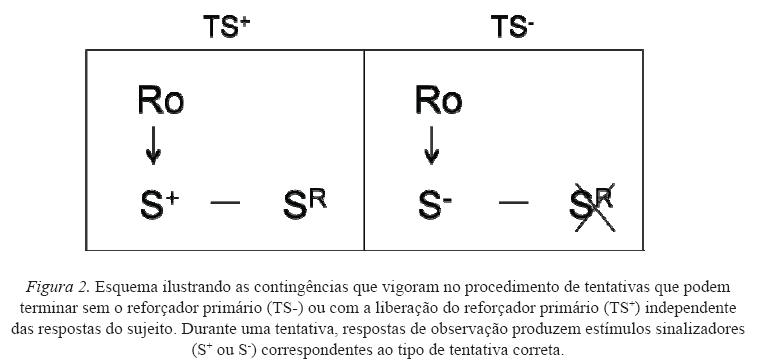
Basicamente, o procedimento é formado por tentativas discretas. As tentativas que terminam com e sem a liberação de comida (tentativas TS+ e TS-, respectivamente) são acompanhadas por estímulos que podem ser produzidos emitindo-se respostas ao único manipulando disponível no ambiente experimental. Em linha de base, a produção de ambos os estímulos discriminativos se dá sob um mesmo esquema, por exemplo, de intervalo variável. Posteriormente, em duas condições experimentais distintas, o esquema empregado para a produção de cada estímulo discriminativo é modificado e passa a incorporar exigências adicionais. A comparação do desempenho dos sujeitos frente a estas mudanças nos esquemas fornece parâmetros do valor reforçador de cada estímulo a partir da freqüência de respostas de observação nos dois tipos de tentativas e/ou da produção de cada um dos estímulos. Um procedimento como esse possui características favoráveis à sua utilização:
1. Fornece uma situação experimental simples, reduzida nos números de fatores que possam interferir nas respostas de observação e dificultar sua análise.
2. Já foi replicado com macacos (Schrier et al., 1980), pombos (Benvenuti & Tomanari, 2010; Blanchard, 1975; Tomanari, 2001b; 2008) e humanos (Tomanari, 2004), fornecendo, assim, parâmetros sistemáticos para a discussão acerca de possíveis diferenças entre espécies.
3. Ainda que as respostas na presença dos estímulos discriminativos não contenham conseqüências programadas, a análise delas poderá fornecer evidências de discriminação.
Tomanari et al. (1998), Tomanari (1997, Experimento 2) e Tomanari (2001b) constituíram uma seqüência de trabalhos com pombos que empregou o procedimento de tentativas cuja liberação de comida independia de resposta do sujeito e manipularam sistematicamente o esquema ao qual as respostas de observação foram expostas.
Tomanari et al. (1998) submeteram pombos privados de comida a sessões compostas por tentativas discretas separadas por intervalos entre tentativas. Com igual probabilidade de ocorrência e independentemente de qualquer resposta do sujeito, metade das tentativas encerrava-se com a apresentação de comida e metade encerrava-se sem apresentação de comida. Em ambos os tipos de tentativa, o único disco de resposta presente na caixa experimental permanecia iluminado por luz branca a menos que ocorressem respostas de observação. Em caso positivo, a cor do disco tornava-se verde (S+) ou vermelha (S-), de acordo com o tipo de tentativa em vigor.
Em condições de linha de base, VI 15 s era o esquema utilizado para as respostas de observação, tanto para a produção de S+ quanto de S-. Em duas condições experimentais, visando à avaliação da função reforçadora condicionada de S+ e de S-, as tentativas TS+ e TS- foram diferenciadas quanto aos esquemas em vigor. Em uma das condições, foi superposto ao VI o esquema DRL 6 s para a produção de S+, enquanto a produção de S- foi mantida em VI. Na outra condição, ao contrário, o esquema DRL foi introduzido para a produção de S-, enquanto a produção de S+ foi mantida em VI.
No esquema tand VI DRL, a apresentação do estímulo discriminativo era contingente ao cumprimento das exigências do VI e, a seguir, sem qualquer sinalização programada, das exigências do DRL. No esquema DRL, a resposta que produzia a apresentação do estímulo discriminativo deveria manter um intervalo mínimo de 6 s da resposta imediatamente anterior. Respostas que não satisfizessem tal intervalo mínimo não eram acompanhadas pela apresentação do estímulo discriminativo e reiniciavam um novo intervalo. Por isso, em esquema DRL, taxas baixas de resposta resultavam na apresentação do estímulo discriminativo. Ao contrário, taxas altas ou nulas de respostas resultavam na sua ausência.
Como se compararam as freqüências de apresentação de S+ e de S- quando, em condições distintas, a apresentação de cada uma delas encontrava-se sob as exigências do esquema tand VI DRL? Os resultados de Tomanari et al. (1998) mostraram que, em esquema tand VI DRL, S+ e S- controlaram de maneira distinta o desempenho dos sujeitos no que se refere à produção de tais estímulos. Quando este esquema foi aplicado à produção de S+, os intervalos entre as respostas foram, majoritariamente, iguais ou maiores que 6 s, resultando na produção desse estímulo em praticamente todas as tentativas apresentadas. Quando associado à produção de S-, ao contrário, os intervalos entre as respostas foram, em sua maioria, menores do que 6 s, resultando na baixa produção desse estímulo relativamente ao número de tentativas apresentadas. Aparentemente, portanto, S+ demonstrou propriedades reforçadoras condicionadas, dado que o responder cumpriu as exigências do esquema tand VI DRL e manteve a apresentação do estímulo. Em contrapartida, S- teria demonstrado propriedades aversivas condicionadas, dado que o responder normalmente não cumpria as exigências do DRL e, portanto, S- foi, em relação ao total de oportunidades, pouco apresentado.
Um dado especialmente interessante nos resultados de Tomanari et al. (1998) tratase da constatação de que, sob as mesmas condições do esquema tand VI DRL, a freqüência de respostas nas tentativas TS- foi claramente maior do que nas tentativas TS+. Este dado, por um lado, reforça a possibilidade de as respostas em TS- terem se mantido pela esquiva de S-. Entretanto, as mesmas respostas produziam outra conseqüência, o prolongamento da duração das tentativas TS-. No procedimento empregado, o encerramento das tentativas dava-se somente na ausência de resposta em seus 3 s finais. Caso houvesse uma resposta nesse período, a tentativa era prolongada por 3 s contados a partir da emissão da resposta. Os resultados descritos por Tomanari et al. (1998) revelam, portanto, uma aparente contradição. Ao mesmo tempo em que as respostas de observação evitavam a apresentação de S-, a duração das tentativas com as quais este estímulo estava associado era marcadamente prolongada. Diante dessa constatação, a interpretação dos resultados baseada na função aversiva de S- precisou ser reavaliada.
Tomanari (2001b) deu continuidade a Tomanari et al. (1998) e buscou investigar, ainda em pombos, a possível função aversiva de S-. Nesse estudo, o esquema tand VI DRL foi estrategicamente substituído por tand VI FR. Sob tand VI FR, ao contrário do esquema tand VI DRL, a função aversiva de S- seria confi rmada pela ocorrência de baixas taxas de respostas de observação, e não por taxas elevadas. Replicando os dados de Tomanari et al. (1998), os pombos produziram S+ e S- em praticamente todas as tentativas de linha de base sob VI 15 s. A introdução do tand VI FR à produção, ora de S+, ora de S-, permitiu comparar as funções reforçadoras exercidas por esses estímulos na manutenção das respostas de observação. Quando o tand VI FR (inicialmente em FR 15 e, posteriormente, em FR 24) esteve em vigor nas tentativas TS-, a freqüência e a distribuição de produção de S- assemelharam-se àquelas em VI. Na medida em que foram aumentadas as exigências do FR para a produção de S-, observou-se um aumento progressivo na freqüência de respostas, resultando na produção de S- em praticamente todas as tentativas TS- na sessão. Quando a apresentação de S+ esteve sob as exigências do tand VI FR, resultados distintos foram verificados. Em tand VI FR 15, observou-se uma pequena diminuição na freqüência de produção de S+, em relação à linha de base, para a maior parte dos sujeitos. O aumento no valor do FR foi acompanhado por uma diminuição ainda mais acentuada na produção de S+.
Frente à literatura na área, os resultados de Tomanari (2001b) são inusitados em diversos aspectos. Primeiro, mostram respostas de observação que acompanham o aumento do valor do FR progressivamente e levam à produção de S-. Esse dado é compatível com a interpretação de que S- exercera funções reforçadoras e, portanto, diverge das demonstrações de suas funções aversivas, especialmente em Blanchard (1975) e Tomanari et al. (1998), os quais utilizaram a mesma base metodológica. Segundo, o fato de os sujeitos não acompanharem o aumento da razão para a produção de S+ confronta dados muito bem estabelecidos na literatura e, aparentemente, revela a existência de fatores aparentemente associados a esse procedimento em particular que precisam ser identificados.
Em Tomanari (2008, Experimento 1), o autor buscou identificar possíveis variáveis que possam ter gerado a alta freqüência de bicadas que resultou, ora na produção de S- (em tand VI FR), ora em sua esquiva (em tand VI DRL), bem como o relativo enfraquecimento do responder em FR quando S+ era a conseqüência. Uma característica potencialmente relevante para esses resultados pode derivar do fato de os esquemas tand VI FR e tand VI DRL controlarem distintamente o desempenho dos sujeitos a partir de aspectos exclusivamente temporais (VI e DRL) ou de aspectos temporais associados a número de respostas (VI e FR). Em esquema tandem, o cumprimento do VI e o início do componente seguinte ocorre sem qualquer sinalização. Sendo DRL, mantém-se uma exigência baseada em características temporais do responder. Sendo FR, a exigência passa a se basear no número de respostas. Em alguma medida, essa alteração no controle do responder pode dificultar a comparação do desempenho em tand VI DRL e tand VI FR.
A partir dessa análise, Tomanari (2008, Experimento 1) avaliou o papel reforçador de S+ e de S- sob esquema tand VI DRH, o qual gera um responder sob controle dos intervalos entre respostas (à semelhança do tand VI DRL), e não sob controle do número de respostas (diferentemente do tand VI FR). Nessa investigação, os valores de DRH foram manipulados (0,8; 0,6; 0,4; 0,2 s) de modo a identificar eventuais funções paramétricas. Entretanto, os resultados mostraram claras distinções funcionais entre S+ e S- apenas sob as exigências de DRH 0,2 s, as quais tenderam a ser cumpridas quando a apresentação de S-, mas não a de S+, era a conseqüência das respostas de observação.
Na medida em que a produção de S- foi alta sob as condições de tand VI DRH (Tomanari, 2008, Experimento 1), esse resultado diverge daqueles obtidos por Blanchard (1975) e Tomanari et al. (1998) no que estes mostraram uma baixa produção de S- sob tand VI DRL. Entretanto, analisada à luz do número de respostas, houve um responder elevado sob tand VI DRL nas tentativas TS- que terminaram por estender marcadamente a duração total da tentativa (Tomanari et al., 1998). Um elevado número de respostas foi igualmente registrado sob tand VI DRH (Tomanari, 2008, Experimento 1), bem como sob tand VI FR (Tomanari, 2001b). Ao que parece, portanto, a freqüência de respostas em TS- sob tand VI DRL, tand VI FR e tand VI DRH tendeu a ser elevada, em todos os três esquemas, independentemente da conseqüência a que estiveram associados. Frente a esse dado, pode-se suspeitar que fatores alheios aos esquemas e conseqüências vigentes devam ter controlado as respostas de observação.
Na tentativa de identificar tais fatores é necessário inicialmente destacar a importância da medida utilizada na avaliação do valor reforçador dos estímulos discriminativos. Blanchard (1975) e Schier et al. (1980) relataram apenas as freqüências de produção de S+ e S- sob linha de base e, fundamentalmente, sob tand VI DRL. Por sua vez, Tomanari et al. (1998), Tomanari (2001b) e Tomanari (2008), entre outras medidas, analisaram também a freqüência de respostas de observação nas tentativas TS+ e TS-, bem como o momento da tentativa em que cada estímulo era produzido. A ampliação dessas medidas mostrou-se extremamente relevante para a compreensão dos resultados. Em Tomanari et al. (1998), a descrição das respostas de observação ao longo das tentativas foi essencial para a constatação do seu aumento surpreendente apenas nas tentativas TS-. Sem tal medida, a contribuição do trabalho iria se restringir a uma replicação bem-sucedida de Blanchard (1975), corroborando a interpretação mais provável, a partir desse dado isolado, de que S- exercera funções aversivas. Em Tomanari (2001b), sob tand VI FR, a exibição de S+ tendia a ocorrer mais freqüentemente no início das tentativas, enquanto a de S- tendia a ocorrer mais ao final delas. Articulado ao fato de que S+ era menos freqüentemente produzido, esse dado compatibiliza a possibilidade de que S+ controlasse discriminativamente outras respostas. Em Tomanari (2008, Experimento 1), dados opostos foram obtidos, na medida em que, sob tand VI DRH, a produção de S- tendeu a ocorrer em momentos anteriores ao de S+. A sensibilidade dessa medida foi capaz de identificar os efeitos dos valores de DRH (0,8 s, 0,6 s e 0,4 s) quando a própria freqüência de produção de S+ e S- ainda não se diferenciava. Portanto, esses dados ratificam a especial importância de analisar as respostas de observação sob a perspectiva de diferentes medidas, as quais podem levar, isoladamente, a interpretações distintas, por vezes incompatíveis ou incompletas.
O momento de produção dos estímulos discriminativos nas tentativas pode contribuir para a compreensão do comportamento dos sujeitos experimentais diante das contingências programadas. Pelas próprias exigências do esquema tandem, seja tand VI DRL, tand VI FR ou tand VI DRH, a produção do estímulo ao qual se encontrava associado deveria necessariamente ocorrer mais tardiamente na tentativa do que o estímulo alternativo, sob VI. Portanto, mediante a exposição repetida às contingências de uma mesma fase experimental, os animais poderiam discriminar entre as tentativas cujo estímulo era exibido rapidamente (sob VI) e aquelas em que o estímulo demorava a aparecer (sob tandem). Portanto, em uma situação como essa, os animais tiveram acesso a fatores temporais discriminativos das tentativas antes mesmo de qualquer mudança na cor do disco. Esses fatores podem ter controlado respostas diversas sob o controle discriminativo, incluindo as próprias respostas de observação. Em tentativas TS+, por exemplo, uma sistemática demora para a mudança na cor do disco pode ter controlado respostas dirigidas ao comedouro, as chamadas respostas preparatórias mantidas pela otimização no consumo do reforço (por exemplo, a salivação facilitando o comer) ou pela minimização da exposição a estímulos aversivos (por exemplo, enrijecimento muscular quando o sujeito é exposto ao choque) (Perkins, 1968). A esse respeito, com base nas observações assistemáticas de um pombo durante algumas sessões experimentais, Tomanari (2001b) relatou a ocorrência de respostas de aproximação ao comedouro durante a apresentação de S+, mas não de S-. Apenas na presença de S+, os sujeitos abaixavam freqüentemente a cabeça, próximo à abertura do comedouro localizada logo abaixo do disco de respostas. Durante a ocorrência de tais respostas, os pombos não podiam emitir respostas ao disco, o que acabou por interferir na emissão de respostas de observação.
Ao considerar possíveis interações entre as respostas de observação e outras respostas, abre-se um caminho para se compreender a freqüência menor de respostas em TS+ do que em TS-. No entanto, não contribui para a compreensão do dado fundamental de Tomanari (2001b) e de Tomanari (2008, Experimento 1), ou seja, a manutenção em si de respostas de observação por S-. Por essa razão, os trabalhos descritos a seguir, Tomanari (1997, Experimento 2) e Tomanari (2008, Experimentos 2 e 3), serão analisados às luz das contribuições que podem trazer para a avaliação das funções dos estímulos discriminativos sob condições na qual o esquema de produção de cada um deles permanecia inalterado.
Tomanari (1997, Experimento 2) manteve o procedimento básico cujas respostas de observação, no esquema de VI 15 s, produziam estímulos discriminativos de tentativas que terminavam com e sem o acionamento do comedouro. Em linha de base, uma vez produzido, o estímulo era exibido até o final da tentativa. Na condição experimental, os estímulos discriminativos eram exibidos por, no máximo, 10 s. Ao final desse período, ainda na mesma tentativa, a situação inicial era restabelecida e os sujeitos podiam voltar a emitir respostas de observação e a reapresentar o estímulo discriminativo corrente.
As comparações entre as freqüências de reapresentações de S+ e S- permitiram avaliar as funções reforçadoras desses estímulos após os sujeitos terem sido expostos a uma apresentação prévia deles. Os resultados de Tomanari (1997, Experimento 2) mostraram que, no que diz respeito à primeira exposição, a produção de S+ era semelhante à de S-. Entretanto, com bastante sistematicidade e regularidade entre os sujeitos, as freqüências de reapresentações de S- foram claramente maiores do que as de S+. Esses dados são compatíveis com a hipótese de que, produzido S+, os animais emitiam respostas dirigidas ao comedouro. Por outro lado, tendo produzido S-, esse estímulo parece ter estabelecido a ocasião para a emissão de reiteradas respostas ao disco branco que produziam as suas reapresentações. Nesse sentido, os dados fortalecem aqueles obtidos em Tomanari (2008, Experimento 1), bem como por Tomanari (2001b), no sentido de haver condições nas quais se demonstra a função reforçadora condicionada de S- .
A fim de especificar cada vez mais essas condições, Tomanari (2008, Experimento 2) analisou a emissão de respostas de observação em função da manipulação das probabilidades de ocorrência de tentativas que terminavam com e sem a apresentação do comedouro. Respostas de observação produziam o estímulo discriminativo do tipo de tentativas em vigor sob esquema de VI 15 s. A partir de uma situação basal de igual probabilidade, passou-se a alterar gradualmente as probabilidades de TS+ e TS-, aumentando- as e diminuindo-as complementarmente até atingir a condição em que apenas TS+ ou apenas TS- compunha a totalidade das tentativas na sessão. Os resultados mostraram que, sob probabilidades intermediárias (0,1 a 0,9 de TS+ ou de TS-), poucos efeitos foram ressaltados às respostas de observação comparativamente à linha de base. O fato de as respostas de observação não variarem significativamente em função das probabilidades intermediárias dos tipos de tentativas sugere, lançando mão de uma interpretação livre, um frágil controle do responder dos sujeitos pela previsibilidade das condições vigentes. Essa interpretação alinha-se aos trabalhos de Eckerman (1973) e Bower, McLean e Meacham (1966), cujos resultados indicavam que uma preferência por uma situação informativa não varia linearmente com a probabilidade de reforçamento. Em uma situação na qual a probabilidade de que respostas de observação produzam S- é de 0,9, por exemplo, é bastante previsível que uma tentativa corrente seja TS- e que as respostas ao disco leve à apresentação de S-. Em uma situação a probabilidade é 0,5, as chances de produzir S+ ou S- são equiprováveis e, portanto, haveria um grau menor de previsibilidade. Nas duas situações, de maior e menor previsibilidade acerca das condições vigentes, os pombos emitiam respostas de observação e produziam os estímulos discriminativos em freqüências semelhantes. As respostas de observação tenderam a se reduzir apenas quando a previsibilidade tornou-se absoluta. Tal redução teria decorrido de uma eventual perda de funções discriminativas?
Analisando-se a emissão de respostas, Tomanari (2008) relata uma clara e sistemática diminuição no responder relativamente à linha de base, tanto na vigência exclusiva de TS+ quanto de TS-. Entretanto, essa diminuição era sempre menos acentuada quando todas as tentativas eram TS+. Quando todas as tentativas eram TS-, o número de respostas ao disco diminuía mais acentuadamente, praticamente extinguindo-se.
Analisando-se a produção dos estímulos, a produção de S+ tendeu a ser mantida em praticamente todas as oportunidades quando todas as tentativas encerravam-se com a apresentação do comedouro. Ao contrário, quando todas as tentativas encerravam-se na ausência do comedouro, houve uma drástica redução, para todos os sujeitos, no número de estímulos produzidos. De maneira distinta aos demais experimentos relatados anteriormente, portanto, esses dados demonstraram condições nas quais as respostas de observação eram mais fortemente mantidas nas condições em que produziam S+ do que S-. Possivelmente, pareamentos S-S possam ter sido gerados a partir das apresentações sistemáticas entre a cor do disco e o acionamento do comedouro. Caso tal fato tenha ocorrido, as funções reforçadoras condicionadas da cor talvez possam ter contribuído para a queda menos acentuada do estímulo associado à TS+ relativamente a TS-. Entretanto, essa sugestão requer cautela, pois, nesse caso, o aumento crescente no número de pareamentos que acompanhou o número de tentativas TS+ deveria ter fortalecido as respostas de observação, e não enfraquecido, como mostraram os dados.
Em prosseguimento à análise do papel das funções discriminativas dos estímulos para a manutenção das respostas de observação, Tomanari (2008, Experimento 3) propôs uma situação experimental cujos estímulos deixavam de exercer funções discriminativas à medida que uma cor era tomada para acompanhar ambos os tipos de tentativas. Todavia, ainda que não fosse discriminativa, a cor não deixava de ser pareada com o acionamento do comedouro metade das vezes em que era apresentada.
Os resultados de Tomanari (2008, Experimento 3) mostraram que o início das condições nas quais apenas uma cor era produzida em ambos os tipos de tentativas foi normalmente acompanhado por um imediato declínio no número de respostas, tendendo a diminuir ainda mais ao longo das sessões, normalmente de forma gradual. Essa redução no responder foi acompanhada por efeitos diversos sobre a produção da cor no disco que, em diferentes fases, para diferentes sujeitos, manteve-se ora como em linha de base, ora reduziu-se. Esses resultados destacam a importância das funções discriminativas dos estímulos para a manutenção das respostas de observação (Rand, 1977). O fato de uma dada cor se correlacionar parcialmente ao acionamento do comedouro não se mostrou suficiente para manter as respostas de observação como em linha de base, ou seja, suficiente para gerar sua apresentação em praticamente todas as tentativas que compunham uma sessão. Ainda que se correlacionasse algumas vezes à apresentação de comida, a cor se correlacionava igualmente à ausência dela. Em termos livres, tratava-se de uma cor pouco confi ável, absolutamente irrelevante na previsibilidade do tipo de tentativa. Na vigência de um mesmo tipo de tentativa (Tomanari, 2008, Experimento 2), a cor era confi ável, mas igualmente irrelevante. Em ambos os casos, eram irrelevantes porque perderam suas funções discriminativas e, portanto, reforçadoras condicionadas para as respostas de observação.
QUESTÕES DE DEBATE NA ÁREA
Adicionalmente à identificação das funções reforçadoras de S- para as respostas de observação, a literatura na área conta com dois grandes pontos de debate, (a) a existência de possíveis especificidades entre espécies quanto ao valor reforçador condicionado de S- e (b) o modelo teórico mais apropriado para se descrever os fatores críticos para a manutenção de respostas de observação, o modelo da redução do atraso e o modelo da redução da incerteza.
Diferenças entre espécies
Segundo alguns autores, diferentes espécies, mais especificamente primatas versus nãoprimatas, seriam distintamente susceptíveis ao reforçamento por S- (Lieberman, 1972; Perone & Baron, 1980; Schrier et al., 1980). Esta possibilidade era levantada com base no fato de que as evidências de que S- manteriam respostas de observação haviam sido encontradas em humanos (Perone & Baron, 1980) e macacos (Lieberman, 1972; Schrier et al., 1980). Entretanto, resultados recentes da literatura enfraquecem as possibilidades de existirem diferenças entre espécies em relação ao reforçamento por S- (Carvalho e Machado, 1992; Roper & Baldwin, 2004, com ratos; Tomanari, 1997; 2001b; 2008, com pombos).
Na mesma direção, Allen e Lattal (1989) e Kaminski e Moore (1990) realizaram uma análise experimental de procedimentos que poderiam gerar um controle fi dedigno de respostas de observação pela produção de S- apenas. Kaminski e Moore (1990) realizaram um experimento com pombos no qual procederam à retirada de S+ por meio de um desvanecimento gradual. Com este recurso, as respostas de observação foram mantidas quando produziam apenas S-. Allen e Lattal (1989) usaram um procedimento engenhoso cujas respostas em extinção diminuíam a densidade de reforços no componente alternativo. Nesta situação específica, as respostas de observação que produziam S- foram mantidas.
O fato de não haver diferenças espécies-específicas com relação ao controle das respostas de observação por S- não significa que primatas e não primatas compartilhem contextos de análise diretamente comparáveis. Por exemplo, em estudos sobre as respostas de observação em humanos têm-se verificado que, além das variáveis identificadas nos procedimentos com não-humanos, outras coexistem e aumentam a complexidade da análise. Por exemplo, as instruções, sejam elas fornecidas pelo experimentador antes da sessão (Case et al., 1995; Fantino & Case, 1983) ou formuladas pelos próprios sujeitos (Perone & Kaminski, 1992), parecem desempenhar um papel importante nas funções exercidas pelos estímulos discriminativos. Papel igualmente importante também parece ter a história passada dos sujeitos com instruções (Lieberman et al., 1997; Perone & Kaminski, 1992). No estudo de Perone e Kaminski (1992), por exemplo, quando S- era a frase “o status do programa agora é ausência de pontos”, a freqüência de respostas de observação dos sujeitos humanos diminuía drasticamente; quando era a frase “o status do programa agora é B” (previamente definido como ausência de pontos), as respostas de observação não se reduziram. Segundo Escobar e Bruner (2009), nessa condição em que as contingências não são descritas ao participante, há possivelmente estabelecimento de uma relação acidental entre o S- e o reforçador.
Modelo da redução do atraso e modelo da redução da incerteza
O modelo da redução do atraso prevê que um estímulo inicialmente neutro se torne um reforçador condicionado em função do quanto sinaliza uma redução temporal entre sua apresentação e a liberação do reforçador incondicionado (Fantino, 1977; Fantino & Logan, 1979; Fantino, Preston, & Dunn,1993). A partir de uma base epistemológica distinta, o modelo da redução da incerteza considera as características preditivas dos estímulos como os determinantes de suas funções reforçadoras (Berlyne, 1957; Hendry, 1969; 1983). Ambos os modelos prevêem, portanto, que um estímulo correlacionado à apresentação do reforçador primário deva se tornar um reforçador condicionado. Entretanto, em relação ao estímulo correlacionado a uma baixa ou nula probabilidade de apresentação de comida, os dois modelos prevêem resultados opostos.
No procedimento de respostas de observação, segundo o modelo da redução do atraso, S- não deveria adquirir função reforçadora condicionada, ou deveria adquirir função punitiva, porque sinaliza que o reforçador está longe temporalmente, isto é, que irá demorar a ser liberado. Por outro lado, segundo o modelo da redução da incerteza, S- seria reforçador por diminuir a incerteza acerca das contingências em vigor, haja vista que a incerteza seria aversiva, reforçadora negativa de respostas que a elimine ou a evite.
Os resultados de Tomanari et al. (1998), Tomanari (2001b), Tomanari (2008, Experimento 1) e Tomanari (1997, Experimento 2) fornecem suporte tanto ao modelo da redução da incerteza quanto ao modelo da redução do atraso, a depender de quais aspectos são considerados na análise. Tomanari et al. (1998) mostraram um elevado responder em TS-. Tomanari (2001b, 2008) mostrou a produção de S- nos esquemas tand VI FR e tand VI DRH, respectivamente. Tomanari (1997) mostrou uma elevada freqüência de reapresentações de S-. Em termos da freqüência de respostas ou produção do estímulo, os resultados aparentemente fortaleceriam o modelo da redução da incerteza. Entretanto, S- foi produzido em baixas freqüências (Tomanari et al., 1998) e normalmente ao final das tentativas (Tomanari, 2001b), indicativos de funções reforçadoras pouco preponderantes, tal como prevê o modelo da redução do atraso. Em Tomanari (1997, Experimento 2), as reapresentações de S- não poderiam ser mantidas pela eliminação da incerteza, pois esta teria sido anulada na primeira exposição do estímulo. Tampouco seria tal desempenho oriundo de qualquer redução do atraso ao reforçador.
As divergências e contradições entre os diferentes aspectos dos dados, que parecem dar ou não suporte aos modelos teóricos de reforçamento condicionado, acabam por revelar as próprias limitações de ambos modelos quando tomados isoladamente. A compreensão do processo de reforçamento condicionado, ao menos no procedimento de respostas de observação, parece exigir uma avaliação mais ampla das variáveis envolvidas que, eventualmente, busque integrar aspectos de posições teóricas que, aparentemente, têm sido tomadas como divergentes ou excludentes (Fantino & Silberberg, 2010).
REFERÊNCIAS
Allen, K. D., & Lattal, K. A. (1989). On conditional reinforcing effects of negative discriminative stimuli. Journal of the Experimental Analysis of Behavior, 52, 335-339. [ Links ]
Berlyne, D. E. (1957). Uncertainty and conflict: A point of contact between information theory and behavior concepts. Psychological Review, 64, 329-333. [ Links ]
Benvenuti, M. F. L., & Tomanari, G. Y. (2010). Respostas de observação mantidas por estímulos compostos em pombos. Acta Comportamentalia, 18, 9-33. [ Links ]
Blanchard, R. (1975). The effect of S- on observing behavior. Learning and Motivation, 6, 1-10. [ Links ]
Bower, G., McLean, J., & Meacham, J. (1966). Value of knowing when reinforcement is due. Journal of Comparative and Physiological Psychology, 62, 184-192. [ Links ]
Carvalho, S. G., & Machado, L. M. C. M. (1992). Esquemas mistos e múltiplos concorrentes: uma reavaliação da resposta de observação. Acta Comportamentalia, 0, 109-144. [ Links ]
Case, D. A., & Fantino, E. (1981). The delay-reduction hypothesis of conditioned reinforcement and punishment: Observing behavior. Journal of the Experimental Analysis of Behavior, 35, 93-108. [ Links ]
Case, D. A., & Fantino, E. (1989). Instructions and reinforcement in the observing behavior of adults and children. Learning and Motivation, 20, 373-412. [ Links ]
Case, D. A., Fantino, E., & Wixted, J. (1985). Human observing maintained by negative informative stimuli only if correlated with improvement in response efficiency. Journal of the Experimental Analysis of Behavior, 43, 289-300. [ Links ]
Case, D. A., Ploog, B. O., & Fantino, E. (1990). Observing behavior in a computer game. Journal of the Experimental Analysis of Behavior, 54, 185-199. [ Links ]
DeFulio, A., & Hackenberg, T. D. (2008). Combinations of response-dependent and response-independent schedule-correlated stimulus presentation in an observing procedure. Journal of the Experimental Analysis of Behavior, 89, 299–309.
Dinsmoor, J. A. (1983). Observing response and conditioned reinforcement. Behavioral and Brain Sciences, 6, 693-704. [ Links ]
Dinsmoor, J. A., Bowe, C. A., Green, L., & Hanson, J. (1988). Information on response requirements compared with information on food density as a reinforcer of observing in pigeons. Journal of the Experimental Analysis of Behavior, 49, 229-237. [ Links ]
Dinsmoor, J. A., Mulvaney, D. E., & Jwaideh, A. R. (1981). Conditioned reinforcement as a function of duration of stimulus. Journal of the Experimental Analysis of Behavior, 36, 41-49. [ Links ]
Eckerman, D. A. (1973). Uncertainty reduction and conditioned reinforcement. The Psychological Record, 23, 39-47. [ Links ]
Escobar, R., & Bruner, C. A. (2009). Observing responses and serial stimuli: Searching for the reinforcing properties of the S-. Journal of the Experimental Analysis of Behavior, 92, 215–231.
Fantino, E. (1977). Conditioned reinforcement: Choice and information. Em W. K Honig & J. E. R. Staddon (orgs.), Handbook of operant behavior. New York: Prentice Hall. [ Links ]
Fantino, E., & Case, D. A. (1983). Human observing: maintained by stimuli correlated with reinforcement but not extinction. Journal of the Experimental Analysis of Behavior, 40, 193-210. [ Links ]
Fantino, E., Case, D. A., & Altus, D. (1983). Observing reward-informative and uninformative stimuli by normal children of different ages. Journal of the Experimental Analysis of Behavior, 36, 437-452. [ Links ]
Fantino, E., & Logan, C. A. (1979). The experimental analysis of behavior: A biological perspective. San Francisco: Freeman. [ Links ]
Fantino, E., Preston, R. A., & Dunn, R. (1993). Delay reduction: Current status. Journal of the Experimental Analysis of Behavior, 40, 193-210. [ Links ]
Fantino, E., & Silberberg, A. (2010). Revisiting the role of bad news in maintaining human observing. Journal of the Experimental Analysis of Behavior, 93, 157-170. [ Links ]
Hendry, D. P. (1969). Introduction. Em D. P. Hendry (Org.), Conditioned reinforcement. Homewood, IL: Dorsey. [ Links ]
Hendry, D. P. (1983). Uncertainty, information, observing. Behavioral and Brain Sciences, 6, 708-709. [ Links ]
Kaminski, B. J., & Moore, J. (1990). A fading procedure facilitates the maintenance of observing responses when the more valued stimulus is not produced. Animal Learning and Behavior, 18, 423-433. [ Links ]
Lieberman, D. A, Cathro, S. S., Nichol, K., & Watson, E. (1997). The role of S- in human observing behavior: Bad news is sometimes better than no news. Learning and Motivation, 28, 20-42. [ Links ]
Lieberman, D. A. (1972). Secondary reinforcement and information as determinants of observing behavior in monkeys (Macaca mulatta). Learning and Motivation, 3, 341-358. [ Links ]
Madden, G. J., & Perone, M. (1999). Human sensitivity to concurrent schedules of reinforcement: Effects of observing schedule-correlated stimuli. Journal of the Experimental Analysis of Behavior, 71, 303- 318. [ Links ]
Mueller, K. L., & Dinsmoor, J. A. (1984). Testing the reinforcement properties of S-: A replication of Lieberman’s procedure. Journal of the Experimental Analysis of Behavior, 41, 17-25.
Mueller, K. L., & Dinsmoor, J. A. (1986). The effect of negative stimulus presentations on observing-response rates. Journal of the Experimental Analysis of Behavior, 46, 281-291. [ Links ]
Mulvaney, D. E., Dinsmoor, J. A., Jwaideh, A. R., & Hughes, L. H. (1974). Punishment of observing by the negative discriminative stimulus. Journal of the Experimental Analysis of Behavior, 21, 37-44. [ Links ]
Perkins, C. C. Jr. (1968). An analysis of the concept of reinforcement. Psychological Review, 75, 155-172. [ Links ]
Perone, M., & Baron, A. (1980). Reinforcement of human observing behavior by stimulus correlated with extinction or increased effort. Journal of the Experimental Analysis of Behavior, 34, 239-261. [ Links ]
Perone, M., & Kaminski, B. J. (1992). Conditioned reinforcement of human observing behavior by descriptive and arbitrary verbal stimuli. Journal of Experimental Analysis of Behavior, 58, 557-575. [ Links ]
Preston, G. C. (1985). Observing responses in rats: Support for the secondary reinforcement hypothesis. The Quarterly Journal of Experimental Psychology, 37B, 23-31. [ Links ]
Rand, J. F. (1977). Behaviors observed during S- in a simple discrimination learning task. Journal of the Experimental Analysis of Behavior, 27, 103-117. [ Links ]
Roper, K. L., & Baldwin, E. R. (2004). The two-alternative observing response procedure in rats: Preference for non-discriminative stimuli and the effect of delay. Learning and Motivation, 35, 275-302. [ Links ]
Roper, K. L., & Zentall, T. R. (1999). Observing in pigeons: The effect of reinforcement probability and response cost using a symmetrical choice procedure. Learning and Motivation, 30, 201-220. [ Links ]
Schaal, D. W., Odum, A. L., & Shahan, T. A. (2000). Pigeons may not remember the stimuli that reinforced their recent behavior. Journal of the Experimental Analysis of Behavior, 73, 125-139. [ Links ]
Schrier, A. M., Thompson, C. R., & Spector, N. R. (1980). Observing behavior in monkeys (Macaca arctoides): Support for the information hypothesis. Learning and Motivation, 11, 355-365. [ Links ]
Shahan, T. A. (2002). The observing-response procedure: A novel method to study drug-associated conditioned reinforcement. Experimental and Clinical Psychology, 10, 3-9. [ Links ]
Shahan, T. A., & Podlesnik, C. A. (2005). Rate of conditioned reinforcement affects observing but not resistance to change. Journal of the Experimental Analysis of Behavior, 84, 1-17. [ Links ]
Shahan, T. A., Magee, A., & Dobberstein, A. (2003). The resistance to change observing. Journal of the Experimental Analysis of Behavior, 80, 273-293. [ Links ]
Skinner, B. F. (1981). Selection by consequences. Science, 213, 501-504. [ Links ]
Tomanari, G. Y. (1997). Parâmetros da resposta de observação. Tese de doutoramento parcialmente publicada, Instituto de Psicologia, Universidade de São Paulo, São Paulo, Brasil. [ Links ]
Tomanari, G. Y. (2001a). Reforçamento condicionado. Revista Brasileira de Terapia Comportamental e Cognitiva, 1, 61-77. [ Links ]
Tomanari, G. Y. (2001b). Respostas de observação controladas por estímulos sinalizadores de reforçamento e extinção. Acta Comportamentalia, 2, 119-143. [ Links ]
Tomanari, G. Y. (2004). Human observing behavior maintained by S+ and S-: Preliminary data. International Journal of Psychology and Psychological Therapy, 4, 155-163. [ Links ]
Tomanari, G. Y. (2008). Resposta de observação: as principais questões na área analisadas a partir de três experimentos com pombos. Tese de Livre-Docência parcialmente publicada, Instituto de Psicologia, Universidade de São Paulo, São Paulo, Brasil. [ Links ]
Tomanari, G. Y. (2009). Respostas de observação: uma reavaliação. Acta Comportamentalia, 17, 259-277. [ Links ]
Tomanari, G. Y., Machado, L. M. C., & Dube, W. V. (1998). Pigeons’ observing responses and responseindependent food presentations. Learning and Motivation, 29, 249-260.
Wyckoff, L. B. (1952). The role of observing responses in discrimination learning - Part I. Psychological Review, 59, 431-442. [ Links ]
Wyckoff, L. B. (1969). The role of observing responses in discrimination learning. Em D. P. Hendry (Org.). Conditioned Reinforcement. Homewood, IL: Dorsey Press. [ Links ]
Received: February 27, 2010
Accepted: July 12, 2010
1 Texto integrante da tese de Livre-Docência do autor (Tomanari, 2008).
2 A realização desse trabalho teve o apoio da Fapesp e do CNPq na forma de auxílios (Fapesp e CNPq) e bolsa produtividade em pesquisa (CNPq).
3 Endereço para correspondência: Instituto de Psicologia, USP. Av. Prof. Mello Moraes, 1721 - São Paulo, SP, 05508-030. Email: tomanari@usp.br.

