










Serviços Personalizados
Journal

artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista Latinoamericana de Psicología
versão impressa ISSN 0120-0534
Rev. Latinoam. Psicol. v.40 n.2 Bogotá ago. 2008
ARTÍCULOS
Modulación contextual de la tolerancia asociativa al etanol1
Ocassion setting of associative tolerance to ethanol
Ronald Betancourt Mainhard; Marion Inostroza Parodi; Mario A. Laborda Rojas2
Universidad de Chile, Chile
ABSTRACT
There is evidence that drug-paired cues not only become associated with the drug effects but also become occasion setters that modulate the association of other cues with the drug effects, contributing to the development of associative tolerance (Ramos, Siegel, & Bueno, 2002). Using a feature-positive discrimination training, we evaluated the contribution of contexts as occasion setters of the ataxic effect of ethanol in rats. The results suggest that the context acquire occasion setter properties of the drug tolerance, and that these properties can be extinguished.
Keywords: Pavlovian conditioning, Occasion setting, Extinction, Drug tolerance, Ethanol.
RESUMEN
Existe evidencia que señala que las claves pareadas con drogas no sólo son asociadas con los efectos de éstas, sino que también adquieren propiedades modulatorias de la asociación entre otras claves y los efectos de la droga, contribuyendo así al desarrollo de la tolerancia asociativa (Ramos, Siegel & Bueno, 2002). Utilizando un procedimiento de discriminación de rasgo positivo, en la presente investigación evaluamos la contribución de los contextos como moduladores del efecto atáxico del etanol en ratas. Los resultados sugieren que el contexto adquiere propiedades modulatorias de la tolerancia a las drogas y que estas propiedades pueden ser extinguidas.
Palabras clave: Condicionamiento pavloviano, Modulación, Extinción, Tolerancia a las drogas, Etanol.
INTRODUCCIÓN
Siegel y sus colaboradores (Larson & Siegel, 1998; Siegel, Baptista, Kim, McDonald & Weise-Kelly, 2000; Siegel & Larson, 1996;) han aportado evidencia acerca de la participación del condicionamiento pavloviano como responsable, al menos en parte, de la tolerancia a las drogas. La tolerancia es entendida como una disminución en la responsibidad a una droga producto de repetidas administraciones, en donde la asociación entre las claves pre-droga y el efecto de ésta juega un importante papel (Larson & Siegel, 1998; Ramsay & Woods, 1997; Siegel et al., 2000). Las claves presentes durante la administración de la droga actuarían como estímulos condicionados (ECs) provocando respuestas condicionadas compensatorias (RCCs) que contribuyen a la tolerancia. Este punto de vista acerca de la tolerancia a las drogas ha fomentado la realización de numerosas investigaciones en busca de establecer cuál es el rol del condicionamiento pavloviano en la tolerancia. Los resultados obtenidos muestran cómo los procedimientos propios del condicionamiento pavloviano alteran el curso de la tolerancia a las drogas.
En el condicionamiento clásico la administración de una droga se analiza como un ensayo de asociación entre eventos en donde el efecto de la droga es el evento significativo (EI), y los eventos que acompañan a la droga, es decir, aquellos eventos que ayudarían a predecir el EI, serían los estímulos condicionados (EC). Cuando se administra una droga el organismo se ve alterado por el efecto de ésta y se provocan respuestas incondicionadas (RI) de homeostasis para compensar dichos efectos. Los eventos que acompañan la administración de la droga se asocian con los efectos incondicionados de ésta, provocando respuestas condicionadas compensatorias que, al igual que las respuestas adaptativas incondicionadas (RI), equilibran la perturbación producida por el EI. Por ejemplo, se ha comprobado que las claves existentes en la administración de etanol provocan respuestas condicionadas compensatorias que contrarrestan sus efectos atáxicos (Betancourt, 2002; Larson & Siegel, 1998; White, Roberts & Best, 2002).
La disminución del efecto de una misma cantidad de droga tras repetidas administraciones, o bien la necesidad de aumentar la cantidad de droga para obtener el mismo efecto inicial, se denomina tolerancia. La tolerancia, como se señaló anteriormente, es atribuida a respuestas compensatorias opuestas al efecto de la droga, en donde intervienen mecanismos de retroalimentación y compensatorios para mantener la homeostasis. En la mantención de la homeostasis el aprendizaje tiene un rol esencial que permite al organismo predecir perturbaciones, anticipando sus efectos. La tolerancia es entonces el producto de los procesos compensatorios de la droga provocados por claves que en el pasado fueron pareadas con los efectos de la misma (Pinel, 2001).
Una de las características de la tolerancia asociativa es la especificidad situacional. Ésta es demostrada en experimentos en los cuales la droga (por ejemplo morfina) es administrada repetidas veces en presencia de claves ambientales específicas. Después de repetidas administraciones el organismo desarrolla tolerancia a la droga (efecto hipoanalgésico). Pero esta tolerancia es mucho más pronunciada cuando se está en presencia de las claves que fueron asociadas con el efecto de la droga (morfina), que en aquellas claves que no estuvieron presentes en la asociación. Este efecto de especificidad situacional se ha demostrado con diversas drogas y se ejemplifica dramáticamente por los efectos letales de "sobredosis" que ocurren cuando los adictos consumen droga en ausencia de las claves que fueron previamente pareadas con la administración de ésta (Siegel & Ramos, 2002).
La especificidad es una demostración del rol del condicionamiento en la tolerancia, ya que las claves asociadas a la droga provocan la respuesta compensatoria que atenúa el efecto de ésta. Esto se traduce en que el efecto de la tolerancia es manifiesto en presencia de las claves y no lo es cuando éstas están ausentes (Betancourt, 2002). Adicionalmente al hecho de que la especificidad situacional provee bases empíricas de la participación del condicionamiento clásico en la tolerancia, se han realizado manipulaciones no farmacológicas de los EC (las claves presentes en la administración de drogas). Los esfuerzos han seguido la lógica de que si estas manipulaciones afectan el curso del condicionamiento, debieran también afectar el curso del desarrollo de la tolerancia. Existe evidencia de que los estímulos condicionados a la droga se ven afectados por procedimientos de extinción (Betancourt, Corada, Dominichetti, Laborda, Martínez & Miguez, 2008; Brooks, 2005; Crombag & Shaham, 2002; MacRae & Siegel, 1997), inhibición externa (Siegel et al., 2000; Siegel & Ramos, 2002), pre-exposición del EC (Goodinson & Siegel, 1995), ensombrecimiento (Walter & Riccio, 1983), bloqueo (Dafters, Hetherington & McCartney, 1983) manipulaciones con glucosa (Siegel, 1999a) y modulación (Ramos, Siegel & Bueno, 2002), entre otros (Para una revisión, ver Siegel et al., 2000; Siegel, 2005).
De especial importancia para la presente investigación son los fenómenos de Modulación y Extinción. El primero de éstos se refiere, en general, al potencial de un estímulo para resolver el valor de una clave ambigua; esto es, cuando existen claves que no logran predecir un evento debido a un historial de reforzamientos ambiguos, cobran importancia otros eventos o estímulos que vienen a resolver las ambigüedades gracias a su asociación con la relación existente entre el estímulo condicionado y el estímulo incondicionado. En la literatura referente a este fenómeno encontramos que se le ha denominado de diversas maneras: "Facilitation" (Rescorla, 1986a), "Occasion Setting" (Holland, 1992) y "Modulation" (Swartzentruber, 1995). En la presente investigación nos referiremos a este fenómeno como "Modulación" (para una acabada revisión del fenómeno de modulación, ver Schmajuk & Holland, 1998).
Como ejemplo podemos considerar que un estímulo A (estímulo condicionado) sea seguido por el EI (A+), pero solamente si A es precedido por el estímulo X (Modulador); esto es lo que se llama un entrenamiento de discriminación de rasgo positivo (X:A+/A-). La situación inversa es aquella en que un estímulo A es seguido por el EI (A+), sólo en aquellas situaciones en que A no es precedido por X; a esta relación se le denomina entrenamiento de discriminación de rasgo negativo (X:A-/A+). Ambos procedimientos pueden ser presentados simultánea o serialmente, dependiendo del arreglo temporal entre X y A.
La evidencia sugiere que al llevar a cabo procedimientos simultáneos se favorece la adquisición de propiedades excitatorias por parte de X, mientras que los procedimientos seriales favorecen la adquisición de propiedades moduladoras. Lo anterior se hace evidente mediante el procedimiento de extinción; esto es, el exponer reiteradamente "A" por si mismo no debilita las respuestas condicionadas si el procedimiento de adquisición fue serial, mientras que sí lo hace si éste fue simultáneo (Holland, 1992; Ross & Holland 1981). Las investigaciones que han intentado extinguir un modulador presentándolo reiteradamente sin ser seguido del estímulo incondicionado no han logrado influenciar sus propiedades de modulación de la asociación entre los estímulos condicionado e incondicionado, sino que tan sólo eliminan la relación directa entre el modulador y el estímulo incondicionado, sugiriendo así, cierta independencia entre las propiedades excitatorias y moduladoras que posee el modulador (Holland, 1992; Ross & Holland, 1981).
En los estudios de modulación se han realizado grandes esfuerzos para aclarar si los contextos adquieren o no cualidades de modulador tal como lo hacen otras claves medio ambientales (Para una revisión ver Hall & Mondragón, 1998). Bouton y Swartzentruber (1986) demostraron que los contextos adquieren propiedades moduladoras en experimentos de discriminación. En sus experimentos concluyen que los contextos adquieren la propiedad de modular las respuestas condicionadas a un estímulo condicionado (EC). Sus resultados sugieren que el contexto A, en el cual se reforzó la presencia del EC, se comportó como un modulador positivo; mientras que el contexto B, en cual no se reforzó el EC, adquirió una débil habilidad para modular negativamente. Ellos sugieren que posiblemente el aprendizaje acerca del contexto B fue incompleto, ya que investigaciones precedentes encontraron que la modulación negativa requiere de un entrenamiento mucho más extenso, y que una fuerte modulación negativa se adquiere solo lentamente (Jenkins & Sainsburry, 1970, citado en Bouton & Swartzentruber, 1986)
El fenómeno de modulación estuvo fuera del estudio con drogas hasta que Anagnóstaras y Robinson (1996) lo incluyeron como posible solución a los hallazgos que encontraron en estudios de sensibilización a los efectos psicomotores de la anfetamina. Los autores propusieron que el contexto en el cual se realizó su investigación pudo haber adquirido simples propiedades excitatorias al mismo tiempo que propiedades de modulación de la respuesta sensibilizada, lo cual sería confirmado en sus resultados posextinción donde se demostró que el procedimiento convencional de extinción( EC sin EI) no debilitó las propiedades moduladoras del contexto, sino que eliminó tan sólo las excitatorias (Holland, 1992; Ross & Holland 1981).
Ramos et al. (2002) recientemente han aportado evidencia que demuestra el desarrollo de relaciones moduladoras en la tolerancia al etanol. En su investigación utilizaron entrenamientos de discriminación de rasgo positivo tanto seriales como simultáneos para evaluar la contribución de un modulador a la tolerancia del efecto hipotérmico del etanol en ratas. Los datos son concordantes con el análisis asociativo de la tolerancia; con ambos procedimientos la tolerancia fue evidente y la presentación no reforzada del estímulo condicionado no afectó las propiedades de modulación del estímulo modulador de la tolerancia en un entrenamiento de discriminación de rasgo positivo serial, mientras sí lo hizo en el entrenamiento de discriminación de rasgo positivo simultáneo.
En conjunto los hallazgos de Bouton y Swartzentruber (1986), Anagnóstaras y Robinson (1996) y Ramos et al. (2002) sugieren que los contextos pueden tener gran importancia como moduladores de la tolerancia asociativa a las drogas.
Como ya se ha dicho, otro procedimiento relevante para la presente investigación es la extinción. Este procedimiento permite que las respuestas condicionadas adquiridas previamente disminuyan. El mecanismo responsable de esta disminución ha sido objeto de estudio desde que Pavlov (1927) hipotetizó que durante la presentación no pareada del EC se formaba una nueva asociación entre el EC y el EI, esta vez inhibitoria, y que actuaba en sentido contrario de la asociación excitatoria previamente adquirida. Los fenómenos de desinhibición y de recuperación espontánea dieron a Pavlov evidencia para sustentar que durante la extinción no se destruía la asociación excitatoria entre el EC y el EI, sino que se agregaba una nueva asociación. Este análisis de la extinción se ha visto reafirmado por modernas investigaciones en donde la extinción ha resultado ser, consistentemente, un nuevo aprendizaje más que un desaprendizaje (Denniston, Chang & Miller, 2003; Denniston & Miller, 2003; Pearce, 1987).
La investigación contemporánea sugiere que un estímulo (EC) extinguido podría tener las propiedades de un estímulo condicionado inhibitorio, como originalmente planteó Pavlov (ver Blaisdell & Miller, 2001; Bouton, 1993; Calton, Mitchell & Schachtman, 1996; Denniston et al., 2003; Denniston & Miller, 2003; Gewirtz, Brandon & Wagner, 1998; Gewirtz y Falls, 1997; Harris, Jones, Bailey & Westbrook, 2000; Konorski, 1967; Rescorla, 1999; Schachtman, Threlkeld & Meyer, 2000), aunque éstas disten de ser idénticas a las obtenidas por los procedimientos estándar de condicionamiento inhibitorio. Incluso hay autores que han sugerido la posible distinción entre tipos de inhibiciones (Williams, 1995).
En conclusión, si bien aún existe controversia acerca de qué aprendizaje está involucrado en la extinción, la evidencia apunta a que en ella se desarrolla un nuevo aprendizaje en vez de un desaprendizaje y se adquieren al menos algunas propiedades inhibitorias, las cuales se acrecientan a medida que aumentan los ensayos de extinción, sin eliminar o "desaprender" la antigua asociación excitatoria. El contexto juega un factor importante durante la extinción, ya que funcionaría como una señal para que se recupere la asociación excitatoria o la nueva asociación inhibitoria, dependiendo del contexto en que se encuentren los sujetos (Bouton, 1993; Bouton, 1994; Bouton & Nelson, 1994; Bouton & Nelson, 1998; Nelson & Bouton, 1997; Nelson, 2002). El contexto actúa hipotéticamente para resolver la ambigüedad del EC. El EC y el contexto son necesarios para activar la conexión inhibitoria que produce la extinción (Para recientes revisiones, ver Bouton, 2004; Delamater, 2004).
La importancia del contexto durante la extinción y su rol modulador de la nueva asociación inhibitoria nos introduce en el análisis de la relación existente entre la extinción y la modulación. Como ya se expuso, durante la modulación se adquieren propiedades distintas de las excitatorias. Si se somete a extinción un estímulo modulador de la relación de otro estímulo con el EI, sólo se logran eliminar sus propiedades excitatorias, mientras su habilidad para modular dicha asociación permanece intacta (Holland, 1992; Ross & Holland 1981). Rescorla (1986b) examinó distintas formas de extinguir las propiedades modulatorias de un estímulo. En sus experimentos encontró que la modulación no es susceptible de ser extinguida por el procedimiento convencional Pavloviano de presentaciones aisladas del modulador. Rescorla concluyó que la modulación podía ser extinguida exponiendo a los sujetos a las contingencias opuestas al entrenamiento. Esto es, si en la adquisición de la modulación se reforzó el EC sólo si era precedido por el modulador (discriminación de rasgo positivo), en la fase de extinción no se debe reforzar el EC cuando es precedido por el modulador, sino que en su ausencia. Más aún, demostró que incluso omitiendo los ensayos de reforzamiento del estímulo condicionado (sólo con los ensayos no reforzados del EC precedido por el modulador), la extinción resultaba efectiva (Rescorla, 1986b; Miller & Oberling, 1998). Con respecto a la tolerancia, se ha demostrado que ésta disminuye ante la exposición a claves asociadas a los efectos de la droga. De hecho la tolerancia a los analgésicos y los efectos sedantes de la morfina son atenuados por la presentación repetida de las claves pre-droga, lo cual ha sido demostrado también en la tolerancia al etanol (Betancourt, 2002; Siegel, 1999b).
La presente investigación intenta esclarecer el rol que cumplen los contextos en la adquisición de la tolerancia asociativa al etanol, mediante la aplicación de un procedimiento de discriminación de rasgo positivo utilizando el contexto como modulador de la asociación entre las claves pre-droga y el efecto de ésta. Con base en el análisis pavloviano de la tolerancia, y tomando en cuenta la evidencia en torno a la modulación, se aporta evidencia acerca de la adquisición y extinción de la modulación contextual de la tolerancia al etanol.
La presente investigación se centró en verificar la especificidad contextual de la tolerancia al etanol y evaluar si el contexto actúa como modulador de la tolerancia asociativa al efecto atáxico del etanol en ratas en un procedimiento de modulación.
MÉTODO
Sujetos
Veinte ratas albinas (Rattus norvegicus) sin experiencia previa en laboratorio, de alrededor de 100 días y de aproximadamente 300 g al inicio del experimento fueron utilizadas en la presente investigación. Todos los sujetos constituyeron un solo grupo y permanecieron en el Bioterio del Departamento de Ecología perteneciente a la Facultad de Ciencias de la Universidad de Chile.
Aparatos
Plano de deslizamiento. Para medir la respuesta atáxica en ratas se utilizó un plano de deslizamiento diseñado especialmente para esta tarea (Larson & Siegel, 1998). El plano de deslizamiento consistió en un callejón construido con plexiglas de 60 cm de largo, 18 cm de ancho y 30 cm de alto. La parte superior del callejón se encontraba abierta, mientras la parte inferior estaba adherida al suelo en uno de sus extremos por medio de bisagras. Una manivela y un sistema de polea fueron operados para alzar el lado sin bisagras del callejón. La inclinación aumentaba aproximadamente ocho grados por cada vuelta de la manivela y la elevación ocurrió aproximadamente a cuatro grados por segundo. Un transportador situado en el lado con bisagra del callejón indicaba el ángulo de inclinación del plano.
Contextos. Para las sesiones reforzadas se utilizó un contexto A definido como: la sala de experimentación; el plano de deslizamiento, adicionando a este un aromatizador (aromatizante Glade ® con fragancia a potpurrí de jardín campestre, código JOH477); una luz indirecta de 75 vatios ubicada a nivel del suelo de la sala de experimentación; una luz directa de 25 vatios ubicada a un metro sobre el plano de deslizamiento. Para crear el contexto B (sesiones no reforzadas) se eliminó el aroma, se utilizaron paredes azul marino en todo el perímetro inferior del plano de deslizamiento y se excluyó la luz directa.
Estímulos. El estímulo incondicionado (EI), fue el efecto fisiológico de una inyección intraperitoneal de etanol al 99% en una solución al 20% en suero fisiológico. La dosis por sujeto fue de 2 gramos de etanol por kilogramo de peso (esta dosis es similar a la utilizada por Larson & Siegel, 1998; Siegel & Larson, 1996, en sus estudios de etanol con ratas). Los estímulos que configuraron el compuesto fueron una inyección y una luz intermitente producida por una foto estimuladora halógena.
Procedimiento
El diseño del experimento se encuentra ilustrado en la Tabla 1 .
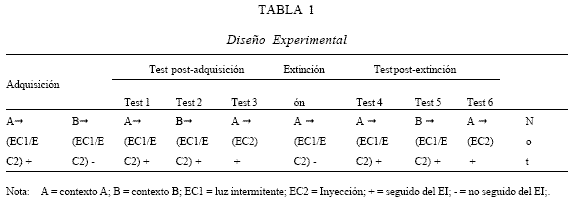
Línea Base. Se establecieron dos líneas base para todos los sujetos. La ataxia fue obtenida a través de la medición (en los minutos 4 y 6 luego de recibir la inyección correspondiente) del grado de deslizamiento. El plano de deslizamiento fue elevado a una velocidad de 4 grados por segundo hasta que los sujetos comenzaban a deslizarse hacia la base del callejón. La línea base 1 (LB1) consistió en medir el grado de deslizamiento de cada sujeto sin la presentación de los estímulos condicionados ni del estímulo incondicionado. Para evitar un efecto de pre-exposición al contexto A, la duración de esta medicion fue solo de 6 minutos, La línea base 2 (LB2) consistió en medir el grado de deslizamientos de cada sujeto durante el primer ensayo reforzado de adquisición [Contx A (EC1/EC2)-> EI], por lo que no puede ser considerado un ensayo de pre-exposición al contexto A o al EI.
Fase de adquisición. La fase de adquisición consistió en la ejecución de los ensayos reforzados [Contx A (EC1/EC2)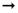 EI], es decir, seguidos de la administración de etanol, y no reforzados [Contx B (EC1/EC2)â Sin EI], seguidos por la administración de suero fisiológico, los cuales fueron realizados en días pares e impares respectivamente, con el objeto de lograr la ambigüedad del compuesto. Los sujetos fueron expuestos al contexto por cinco minutos e inmediatamente después fueron inyectados con la solución de etanol (días reforzados) o con una solución salina (días no reforzados). Una vez terminado este procedimiento se realizaba la medición de la ataxia en los minutos 4 y 6 a partir de la inyección.
EI], es decir, seguidos de la administración de etanol, y no reforzados [Contx B (EC1/EC2)â Sin EI], seguidos por la administración de suero fisiológico, los cuales fueron realizados en días pares e impares respectivamente, con el objeto de lograr la ambigüedad del compuesto. Los sujetos fueron expuestos al contexto por cinco minutos e inmediatamente después fueron inyectados con la solución de etanol (días reforzados) o con una solución salina (días no reforzados). Una vez terminado este procedimiento se realizaba la medición de la ataxia en los minutos 4 y 6 a partir de la inyección.
Test post-adquisición. Se realizaron tres tests post adquisición que consistieron en medir el grado de deslizamiento de los sujetos en la caja estándar en tres situaciones experimentales (este procedimiento es idéntico al realizado en las mediciones de línea base 2):
Test 1: Contx A (EC1/EC2)EI
Test 2: Contx B (EC1/EC2)EI
Test 3: Contx A (EC2)EI
Fase de extinción. Consistió en extinguir el contexto como modulador de la asociación entre las claves pre-droga y el efecto atáxico del etanol en ratas. La fase de extinción se realizó en 8 sesiones, realizadas diariamente, en las que se expuso a los sujetos experimentales al mismo procedimiento realizado en los ensayos reforzados de modulación, pero remplazando la inyección de etanol por una de solución salina.
Test post-extinción. Finalmente, se realizaron tres test post extinción que consistieron en medir el grado de deslizamiento de los sujetos en la caja estándar luego del procedimiento de extinción (este procedimiento es idéntico al realizado en el test post-adquisición):
Test 4: Contx A (EC1/EC2)EI
Test 5: Contx B (EC1/EC2)EI
Test 6: Contx A (EC2)EI
RESULTADOS
Los análisis de los resultados se realizaron a partir de los promedios de desempeño del grupo experimental a través de un diseño antes - después, el cual consistió en comparar las ejecuciones del grupo experimental con sus respectivas líneas bases y los efectos de intervención experimental en las subsecuentes etapas del diseño. Para el análisis estadístico de los datos se utilizó la prueba T-Student para grupos relacionados o pareados.
La Figura 1 nos indica las líneas bases del grupo experimental que se obtuvieron a través de mediciones sin tratamiento (sin inyección de ningún tipo). Los resultados de LB2 fueron obtenidos a partir de la evaluación de su ejecución en la caja de deslizamiento en forma posterior a la primera administración de etanol (EI). El promedio de deslizamiento obtenido en LB1 fue de 42,7º, mientras que el promedio de deslizamiento de LB2 fue de 22,5º. Se presenta una clara diferencia en los resultados, lo que demuestra el efecto atáxico producto de la administración de etanol. El análisis estadístico entre ambas líneas bases arroja una diferencia significativa, t(19) = 7.796, p < 0.05.
Finalizada la fase de adquisición se evaluó la ejecución del grupo experimental en ambos contextos frente al compuesto y bajo los efectos del etanol, lo cual corresponde a la primera administración de etanol de los sujetos en el contexto B.
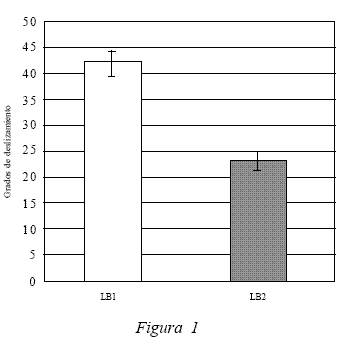
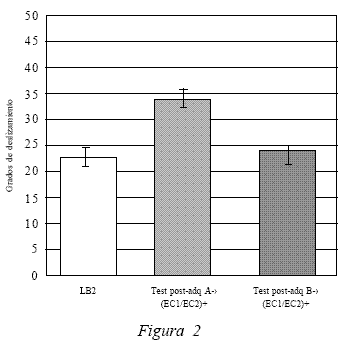
En la Figura 2 se comparan los datos obtenidos en LB2 (22,5º) con el Test 1 (34,2º) (test post-adquisición del compuesto ante el contexto A), en donde se expresa claramente la disminución del efecto atáxico del etanol -evidenciado en la mejora de los puntajes promedios obtenidos con una diferencia significativa, t(19) = 4.441, p < 0.05, producto del tratamiento realizado en la fase de adquisición. Lo anterior permite sostener que los sujetos experimentales desarrollaron un aprendizaje asociativo de respuestas compensatorias de tolerancia a los efectos atáxicos del etanol frente al contexto A. Además, se puede observar que los puntajes de LB2 (22,5º) y el Test 2 (23,4) (test post-adquisición ante el contexto B) no presentan diferencias significativas, según el análisis estadístico, t(19) = -0.326, p > 0.05. Lo anterior confirma que el aprendizaje de respuestas de tolerancia solo se produjo ante el contexto A. Esto permite concluir acerca de la modulación contextual de la tolerancia. La ataxia disminuye sólo frente al compuesto en el contexto A, pero no frente al contexto B, ya que producto de la fase de adquisición el contexto A adquirió propiedades modulatorias de la asociación entre el compuesto (EC1/EC2) y el efecto incondicionado del etanol, lo cual se evidencia en la respuesta condicionada compensatoria de tolerancia.
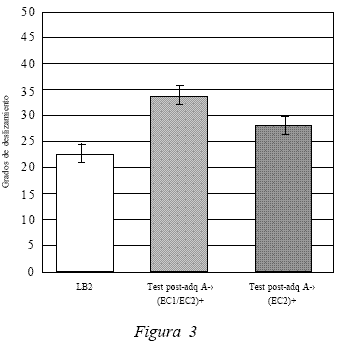
Además de los test post adquisición en el contexto A y B, se evaluó el desempeño post asociación de los sujetos en el contexto A frente a uno de los estímulos del compuesto, la inyección (EC2). Los resultados de este test, el Test 3, se muestran en la Figura 3 . El promedio del desempeño de los sujetos frente al EC2 en el contexto A post asociación fue de 28,9º, al compararlo con el desempeño en LB2 (22,5º) se evidencia una diferencia significativa, t(19) = 2.208, p < 0.05, lo que indica la existencia de respuestas compensatorias de tolerancia provocadas por la inyección (EC2) frente al contexto A. Al comparar la ejecución de los sujetos ante el compuesto y el EC2 en el contexto A también se observa una diferencia significativa, t(19) = 2.255, p < 0.05 (entre el Test 1 y el Test 3).
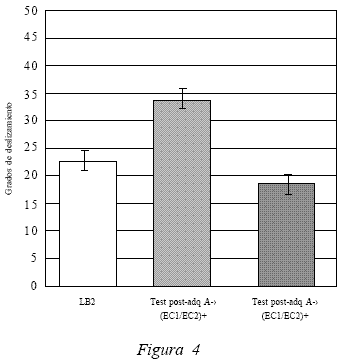
Luego de finalizada la fase de extinción, el grupo experimental fue nuevamente evaluado en la caja de deslizamiento. Los resultados presentados en la Figura 4 indican que la medición de la ejecución post extinción del compuesto frente al contexto A (Test 4) da como resultado un promedio de 18,6º. Al comparar este promedio con el obtenido por los sujetos en el test post adquisición del compuesto frente al contexto A (34,2º) (Test 1), los datos estadísticos señalan una diferencia significativa, t(19) = 6.009, p < 0.05. Lo anterior permite sostener que los sujetos experimentales tras el procedimiento de extinción dejan de presentar respuestas condicionadas compensatorias de tolerancia frente a los efectos atáxicos del etanol, evidenciado a partir de la disminución del desempeño en el plano de deslizamiento. Lo anterior también se sustenta al comparar la ejecución post extinción del compuesto ante el contexto A (Test 4) con LB2, los cuales no presentan diferencias significativas entre ellos t(19) = 1.373, p > 0.05. Es decir, los sujetos experimentales que presentaban respuestas de tolerancia frente a la modulación contextual del contexto A adquirida en la fase de adquisición, dejan de presentar tales respuestas cuando son evaluados en el test post extinción.
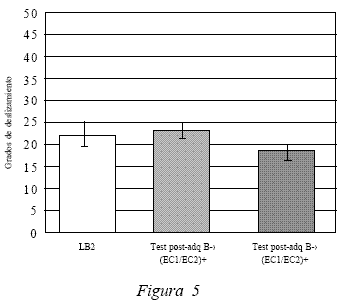
Además de evaluar la ejecución del compuesto frente al contexto A, se realizó una medición del desempeño obtenido por los sujetos ante el compuesto frente al contexto B, Test 5 (ver Figura 5 ). Dicha medición dio como resultado un promedio de deslizamiento de 18,6º, el cual según el análisis de los datos estadísticos presenta una diferencia significativa, t(19) = 2.857, p < 0.05, en comparación con el test post asociación del compuesto frente al contexto B (Test 2) y no presenta diferencias significativas con LB2, t(19) = 1.401, p > 0.05. Tales datos permiten afirmar que a pesar de que los sujetos cuando fueron evaluados post adquisición frente al contexto B no presentaban respuestas condicionadas compensatorias frente a los efectos atáxicos del etanol, luego de realizar el procedimiento de extinción y ser nuevamente evaluados en dicho contexto, los sujetos presentan una disminución significativa de su desempeño en la caja de deslizamiento (ver discusión).
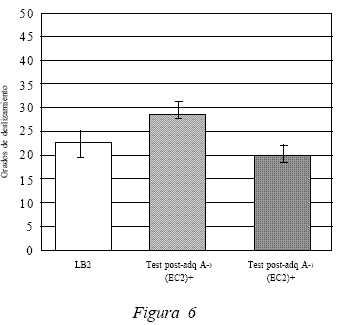
Finalmente, la Figura 6 presenta los datos obtenidos a partir de la medición de uno de los estímulos presentes en el compuesto ante el contexto A, la inyección (EC2). La ejecución de los sujetos obtuvo como resultado un promedio de 20,3º. Al comparar los promedios del test post adquisición del EC2 frente al contexto A (Test 3) con el test post extinción del EC2 frente al contexto A (Test 6), los datos estadísticos arrojan una diferencia significativa, t(19) = 4.234, p < 0.05. Entonces, se puede sostener que tras el procedimiento de extinción, los sujetos ya no ejecutan respuestas condicionadas compensatorias de tolerancia frente a los efectos del etanol ante el EC2 en el contexto A. Lo anterior también se sustenta al comparar la ejecución post extinción del EC2 frente al contexto A (Test 6) con LB2, donde los datos estadísticos establecen que no hay diferencias significativas entre ambos, t(19) = 0.787, p > 0.05.
DISCUSIÓN
La presente investigación se basó en el paradigma del condicionamiento Pavloviano y demostró cómo ratas expuestas a claves ambiguas pareadas con etanol desarrollaron tolerancia a los efectos atáxicos de éste, solamente cuando estas claves fueron presentadas en un contexto que señalizaba la probabilidad de ser seguido por el EI. Dicha evidencia se encontró a través de un procedimiento de discriminación de rasgo positivo, en el cual un contexto A señalizaba la ocurrencia del EI luego de la presentación de un estímulo compuesto (EC1/EC2). Estos resultados se suman a los ya expuestos por Anagnóstaras y Robinson (1996), y por Ramos et al. (2002), quienes indican que algunas claves pareadas con drogas pueden adquirir propiedades moduladoras.
La modulación contextual de los efectos atáxicos del etanol se evidencia a partir de los resultados obtenidos en el test post adquisición, en donde el análisis estadístico permite afirmar que los sujetos después de sucesivos ensayos de pareación desarrollaron respuestas condicionadas compensatorias mejorando así significativamente su ejecución en comparación con LB2. De importancia, estas respuestas compensatorias condicionadas sólo se presentaron cuando los sujetos fueron expuestos al compuesto en el contexto A, no así frente al contexto B. Es decir, el desarrollo de respuestas compensatorias opuestas a los efectos atáxicos del etanol fue modulado por el contexto A producto de que el compuesto tiene una significación ambigua como resultado de haber recibido la misma cantidad de ensayos pareados y no pareados con el EI, tal conclusión es concordante con la evidencia experimental (Boston & Swartzentruber, 1986). Además, los datos obtenidos a partir de los test realizados post-adquisición permiten sugerir que las propiedades que adquirió el contexto no fueron por una asociación directa con el EI, sino que una relación de modulación entre el contexto y el compuesto como provocador de la RC. Esta posibilidad se establece a partir del desempeño que obtuvieron los sujetos al realizar el test post-adquisición frente a sólo uno de los estímulos del compuesto (EC2) en el contexto A. El resultado obtenido indica una diferencia significativa al comparar el EC2 en el contexto A (28,9º) con el compuesto (EC1/EC2) en el contexto A (34,2º), lo que sugiere que si bien ambos presentan tolerancia en este contexto, esta tolerancia es mayor frente al compuesto, lo que a su vez da evidencia de que el contexto no fue asociado directamente con el EI, ya que si ese fuese el caso, la expresión del desempeño debería haber sido mantenida ante la presentación del EC2 y no mostrar una diferencia significativa con la presentación del compuesto como arrojó el análisis estadístico de los resultados.
La segunda fase de esta investigación consistió en someter a extinción la modulación adquirida en la fase de adquisición. Los resultados de los tests post extinción del compuesto (EC1/EC2) en el contexto A y B, y del EC2 en el contexto A, tienen importantes implicancias para el análisis asociativo de la tolerancia al etanol. Lo primero que se observa es que las respuestas condicionadas compensatorias provocadas frente al compuesto en el contexto A disminuyeron luego de la extinción, lo cual se demuestra en la disminución de los grados de deslizamiento al comparar el desempeño post-adquisición y post-extinción (34.2º y 18.6º, respectivamente). Estos hallazgos son concordantes con los encontrados por Rescorla (1986b) al examinar distintas formas de extinguir la modulación. Rescorla encontró que una forma de extinguir la modulación de un estímulo es exponerlo a las contingencias opuestas al entrenamiento (en este caso reforzar el compuesto en el contexto B y no hacerlo en el contexto A, CtxA? EC1/EC2 ? Sin EI // CtxB? EC1/EC2? EI). Incluso omitiendo los ensayos reforzados, como se realizó en esta investigación, la modulación es extinguida (en este caso solo CtxA? EC1/EC2 ? Sin EI). La modulación contextual del contexto A fue extinguida durante la fase de extinción, lo cual se observa en la pérdida de las respuestas condicionadas compensatorias de tolerancia frente al compuesto en dicho contexto.
Paralelamente con la extinción de la modulación contextual del compuesto frente al contexto A, el desempeño de los sujetos frente al EC2 en el contexto A también disminuyó. De acuerdo al procedimiento de la investigación, el contexto A al someterse a extinción perdió sus propiedades modulatorias, lo cual no sólo se evidenció frente al compuesto, sino que también frente a una de sus partes (EC2, inyección).
Si bien en la fase de adquisición los resultados no demuestran que el contexto B haya adquirido alguna asociación, en el test post extinción esto parece menos claro. El desempeño de los sujetos en LB2 fue de 22.5 grados y no tuvo diferencias significativas con el desempeño post adquisición frente al compuesto en el contexto B, 23.4º, de lo cual se concluye que el compuesto no provocó respuestas compensatorias condicionadas en el contexto B, sino que estas respuestas fueron moduladas por el contexto A. Sin embargo, en el test post extinción del compuesto en el contexto B se observan diferencias significativas al compararlo con el test post adquisición (18.6º y 23.4º, respectivamente), evidenciándose respuestas de hipertaxia. Cabe recordar que durante la fase de extinción el contexto B no fue manipulado, lo que hace suponer que algún fenómeno ocurrió tanto en la fase de asociación como en la de extinción, para que en el desempeño post extinción se evidenciara hipertaxia.
Una posible explicación es que durante la fase de asociación, el compuesto estaba siendo pre-expuesto en el contexto B, lo cual lleva a postular el desarrollo de un aprendizaje latente. Existe suficiente evidencia para sustentar que dicho aprendizaje es modulado por los contextos en los cuales se desarrolla (Hall, 1991; Hall & Honey, 1989; Hall & Mondragón, 1998). Si la pre exposición de un estímulo se lleva acabo en un contexto, este adquiere la propiedad para modular la inhibición latente; si se intenta condicionar el estímulo en un nuevo contexto, la inhibición se atenúa o incluso elimina (Para una revisión ver Hall & Mondragón, 1998).
Tomando en cuenta el aprendizaje latente posiblemente adquirido en los ensayos de adquisición en que se expuso el compuesto en el contexto B sin ser seguido del EI, cabe aclarar qué sucedió durante la fase de extinción para fomentar la expresión de dicha inhibición. Al analizar en conjunto los hallazgos acerca de la extinción, y las propiedades inhibitorias que se adquieren mediante ésta, podemos sugerir que en la fase de extinción de la investigación el compuesto adquiere propiedades inhibitorias en relación al EI. Se ha comprobado que dichas propiedades pueden ser adquiridas mediante procedimientos de extinción (Bouton, 1993; Calton et al., 1996; Denniston et al., 2003; Denniston & Miller, 2003; Schachtman et al., 2000; Williams, 1995).
Entonces, una posible explicación de las respuestas de hiperataxia presentes en el test post extinción frente al compuesto en el contexto B, es que durante la fase de adquisición se produjo una inhibición latente de la modulación contextual negativa del contexto B (modulación de la asociación EC1/EC2 ? Sin EI), modulación que se hizo evidente solamente cuando el compuesto adquirió propiedades inhibitorias mediante el procedimiento de extinción. Existe evidencia que sustenta que un modulador negativo no será eficaz si no hay una relación inhibitoria entre el estímulo y el EI, ya que no habría qué modular, señalar o recuperar de la memoria (van Wijk, Maes & Vossen, 1997). Además hay investigaciones que señalan que los procesos involucrados en la inhibición latente, la extinción y el condicionamiento inhibitorio mantienen algunas propiedades comunes (Bouton, 1993; Kraemer & Spear, 1992; Miller, Kasprow, & Schachtman, 1986; Schachtman et al., 2000), lo cual sustentaría nuestra proposición de que el compuesto al adquirir propiedades inhibitorias por medio de la extinción determine la expresión de propiedades modulatorias negativas del contexto B sobre el compuesto, haciéndose explícita la inhibición hasta el momento latente.
Una explicación alternativa de las respuestas de hipertaxia frente al compuesto en el contexto B post extinción, se basa en las propiedades inhibitorias adquiridas por el compuesto mediante la extinción. Sugerimos que el procedimiento discriminativo utilizado en el presente experimento, y que desarrolló modulación positiva del contexto A frente al compuesto, pudo haber también desarrollado modulación negativa del contexto B frente al compuesto pero de forma incompleta. Bouton y Swartzentruber (1986) sugieren esta posible explicación para sus hallazgos en experimentos en los cuales utilizaron discriminaciones contextuales muy similares a las utilizadas en esta investigación en experimentos de supresión condicionada. En su trabajo, Bouton y Swartzentruber encontraron que los contextos no adquirieron relaciones excitatorias ni inhibitorias con el EI. Además, demostraron que el contexto A adquirió propiedades modulatorias positivas, mientras el contexto B adquirió tan solo una débil habilidad modulatoria, posiblemente, debido a los pocos ensayos de entrenamiento. En nuestra investigación encontramos resultados similares basándonos en las respuestas dadas por los sujetos post extinción en el contexto B frente al compuesto. Luego de que el compuesto dejó de ser ambiguo y (posiblemente) adquirió propiedades inhibitorias mediante la extinción, se transformó en un mejor estímulo para la modulación negativa del contexto B (ver van Wijk et al., 1997), y por ello, el desempeño post extinción de los sujetos frente al compuesto en el contexto B disminuyó por debajo de la LB2 evidenciando respuestas hiperatáxicas, demostrando así la posible asociación modulatoria del contexto B y el compuesto.
Un punto importante para el análisis de los resultados generales presentados en la presente investigación es el hecho de que nuestro diseño experimental no permite excluir completamente un análisis configuracional de los datos. Puede ser que los sujetos experimentales adquirieron tan sólo una discriminación entre los compuesto [ContxA(EC1/EC2)+] y [ContxB(EC1/EC2)-], En cuyo caso los contextos jugarían el rol de claves condicionadas y no de moduladores. En futuras investigación resultará fundamental someter esta hipótesis a prueba, posiblemente integrando tests de transferencia de las posibles propiedades modulatorias del contexto a otros compuestos que hayan sido modulados anteriormente.
Finalmente, un aspecto importante acerca de los resultados de la presente investigación es su relación con la problemática de las conductas adictivas. Betancourt (2002) sugirió la estrecha relación entre los hallazgos de laboratorio y la práctica clínica. En esta investigación se ha encontrado evidencia que podría ser de ayuda al intentar explicar los datos provenientes de las adicciones a las drogas. Como es sabido, algunos tratamientos intentan extinguir las asociaciones entre las claves pre-droga y el efecto de éstas. De particular importancia son los tratamientos de "exposición de claves", que intentan extinguir las respuestas de abstinencia con la repetitiva exposición de las claves pre-droga. Como Ramos et al. (2002) y Siegel y Ramos (2002) sugieren, es posible que los resultados mixtos de estas terapias sean producto de que el clínico esté presentando repetidamente un modulador en vez de un estímulo condicionado con lo cual no conseguirían resultados promisorios en la terapia. Una posible sugerencia para las intervenciones clínicas en adicciones, y que se deriva de los presentes resultados, sería señalar la necesidad de extinguir cadenas conductuales completas, integrando posibles moduladores, en vez de la mera presentación repetida de posibles estímulos condicionados de forma aislada. Esta recomendación se sumaría a recientes investigaciones que sugieren que para conducir una extinción efectiva (disminuyendo posibles recaídas) se debería ampliar el número de contextos utilizados en los ensayos de extinción (Chelonis, Calton, Hart & Schachtman, 1999; Gunther, Denniston & Miller, 1998; Rodríguez, Craske, Mineka & Hladek, 1999, pero ver Betancourt et al., 2008; Bouton, García-Gutiérrez, Zilski, & Moody, 2006). Además, Denniston et al. (2003) han sugerido que para minimizar las recaídas se debe: exponer explícitamente las claves de forma no pareada, extinguir en variados contextos y sobreentrenar la extinción en cada uno de ellos.
Los resultados que surgen de esta investigación refuerzan la idea de que si bien el condicionamiento crea un potencial para que un EC provoque la RC, la fuerza de una RC va a depender también de otros estímulos presentes en la situación. Es así como las claves presentes en el contexto A de esta investigación que estuvieron presentes durante la presentación del EC, van a tener efecto sobre la RC e incluso van a determinar la ocurrencia de las respuestas condicionadas compensatorias. Por tanto, si los tratamientos para sujetos que presentan alguna adicción se limitan solamente a aquellos eventos presentes en el tratamiento, otras claves (contextuales) no expuestas tendrán plena capacidad de provocar la respuesta de tolerancia.
Tomando en cuenta nuestros resultados, podemos concluir que en la tolerancia a las drogas se encuentran implicados procesos modulatorios (Ramos et al., 2002) que enriquecen aún más la visión de la tolerancia asociativa. Ramos y colaboradores concluyen que las claves presentes en la administración de droga pueden adquirir propiedades modulatorias de la tolerancia, lo cual en esta investigación es ampliado al comprobar que en este caso el contexto de administración funcionó como modulador de la tolerancia asociativa a las drogas.
REFERENCIAS
Anagnostaras, S. & Robinson, T. (1996). Sensibilization to the psychomotor stimulant effect of amphetamine: Modulation by associative learning. Behavioral Neuroscience, 110 (6), 1397-1414. [ Links ]
Betancourt, R. (2002). Condicionamiento clásico y drogas: Modulación de los procedimientos de extinción a la tolerancia y síntomas de abstinencia al etanol en ratas. Tesis para optar al grado de Doctor en Psicología, Departamento de Psicología, Universidad de Chile, Santiago, Chile. [ Links ]
Betancourt, R., Corada, L., Dominichetti, J., Laborda, M., Martínez, G. & Miguez, G. (2008). Efecto de la extinción en múltiples contextos sobre la renovación de la tolerancia a las drogas. Psicothema, 20 (2), 279-283. [ Links ]
Blaisdell, A. P. & Miller, R. R. (2001). Conditioned inhibition produced by extinction-mediated recovery from the relative stimulus validity effect. A test of acquisition and performance models of empirical retrospective revaluation. Journal of Experimental Psychology: Animal Behavior Processes, 27, 48-58. [ Links ]
Bouton, M. E. (1993). Context, time, and memory retrieval in the interference paradigms of Pavlovian conditioning. Psychological Bulletin, 114, 80-99. [ Links ]
Bouton, M. E. (1994). Conditioning, remembering, and forgetting. Journal of Experimental Psychology: Animal Behavior Processes, 20, 219-231. [ Links ]
Bouton, M. E. (2004). Context and behavioral processes in extinction. Learning & Memory, 11, 485-494. [ Links ]
Bouton, M. E., García-Gutiérrez, A., Zilski, J. & Moody, E. W. (2006). Extinction in multiple contexts does not necessarily make extinction less vulnerable to relapse. Behaviour Research and Therapy, 44, 983-994. [ Links ]
Bouton, M. E. & Nelson, J. B. (1994). Context-specificity of target versus feature inhibition in a feature-negative discrimination. Journal of Experimental Psychology: Animal Behavior Processes, 20, 51-65. [ Links ]
Bouton, M. E. & Nelson, J. B. (1998). Mechanisms of feature-positive and feature-negative discrimination learning in an appetitive conditioning paradigm. En N.A. Schmajuk & P. C. Holland, (Eds), Occasion Setting. Associative Learning and Cognition in Animals. Pp. 383-424. Washington, DC: American Psychological Association. [ Links ]
Bouton, M. E. & Swartzentruber, D. (1986). Analysis of the associative and occasion-setting properties of context participating in a Pavlovian discrimination. Journal of Experimental Psychology: Animal Behavior Processes, 12, 333-350. [ Links ]
Brooks, D. (2005). Alcohol ataxia tolerance: Extinction cues, spontaneous recovery and relapse. International Journal of Comparative Psychology, 18, 141-153. [ Links ]
Calton, J. L., Mitchell, K. G., & Schachtman, T. R. (1996). Conditioned inhibition produced by extinction of a conditioned stimulus. Learning and Motivation, 27, 335-361. [ Links ]
Chelonis, J. J., Calton, J. L., Hart, J.A. & Schachtman, T. R. (1999). The renewal effect in conditioned taste aversion. Learning and Motivation, 30, 1-14. [ Links ]
Crombag, H. S. & Shaham, Y. (2002). Renewal of drug seeking by contextual cues after prolonged extinction in rats. Behavioral Neuroscience, 116,169-173. [ Links ]
Dafters, R., Hetherington, M. & McCartney, H. (1983). Blocking and sensory preconditioning effects in morphine analgesic tolerance: support for a Pavlovian conditioning model of drug tolerance. The Quarterly Journal of Experimental Psychology, 35B:1-11. [ Links ]
Delamater, A. R. (2004). Experimental extinction in pavlovian conditioning: Behavioural and neuroscience perspectives.The Quarterly Journal of Experimental Psychology, 57B, 97-132. [ Links ]
Denniston, J., Chang, R. & Miller, R. R. (2003). Massive extinction treatment attenuates the renewal effect. Learning and Motivation, 34, 68-86. [ Links ]
Denniston, J. & Miller, R. R. (2003). The role of temporal variables in inhibition produced through extinction. Learning & Behavior, 31 (1),35-48. [ Links ]
Gewirtz, J. C., Brandon, S. E. & Wagner, A. R. (1998). Modulation of the acquisition of the rabbit eyeblink conditioned response by conditioned contextual stimuli. Journal of Experimental Psychology: Animal Behavior Processes, 24, 106-117. [ Links ]
Gewirtz, J. C. & Falls, W. A. (1997). Normal conditioned inhibition and extinction of freezing and fear-potentiated startle following electrolytic lesions of medial prefrontal cortex in rats. Behavioral Neuroscience, 111, 712-726. [ Links ]
Goodison, T. & Siegel, S. (1995). Tolerance to Naloxone-Induced Suppression of Intake Learning and Cross-Tolerance to Cholecystokinin in Rats. Behavioral Neuroscience, 109, 455-465. [ Links ]
Gunther, L. M., Denniston, J. C. & Miller, R. R. (1998). Conducting exposure treatment in multiple contexts can prevent relapse. Behaviour Research and Therapy, 36, 75-91. [ Links ]
Hall, G. (1991). Perceptual and associative learning. Oxford, England: Clarendon Press. [ Links ]
Hall, G. & Honey, R. (1989) Perceptual and associative learning. En S. B. Klein & R. R. Mowrer (Eds.), Contemporary Learning Theories. Hillsdale, N. J.: LEA. [ Links ]
Hall, G. & Mondragón, E. (1998). Contextual control as ocassion setting. En N. A. Schmajuk & P. C. Holland, (Eds.)Occasion Setting. Associative Learning and Cognition in Animals. pp.199-221. Washington, DC: American Psychological Association. [ Links ]
Harris, J. A., Jones, M. L., Bailey, G. K. & Westbrook, R. F. (2000). Contextual control over conditioned responding in an extinction paradigm. Journal of Experimental Psychology: Animal Behavior Processes, 26, 174-185. [ Links ]
Holland, P. C. (1992). Ocassion setting in pavlovian conditioning. En D. L. Medin (Ed). The Psychology of Learning and Motivation, 28 (pp. 69-125). San Diego, CA: Academic press. [ Links ]
Kraemer, P. J. & Spear, N. (1992). The effect of nonreinforced stimulus exposure on the strength of a conditioned taste aversion as a function of retention interval: Do latent inhibition and extinction involve a shared process?. Animal Learning & Behavior, 20 (1), 1-7. [ Links ]
Konorski, J. (1967). Integrative activity of the brain: An interdisciplinary approach. Chicago: University of Chicago Press. [ Links ]
Larson, S. & Siegel, S. (1998). Learning and tolerance to the ataxic effect of ethanol. Pharmacology Biochemistry and Behavior, 61, 131-142. [ Links ]
MacRae, J. R. & Siegel, S. (1997). The role of self-administration in morphine withdrawal in rats. Psychobiology, 25, 77-82 [ Links ]
Miller, R. R., Kasprow, W. J. & Schachtman, T. R. (1986). Retrieval variability: Psychobiological sources and consequences. American Journal of Psychology, 99, 145-218. [ Links ]
Miller, R. R. & Oberling, P. (1998). Analogies between occasion setting and pavlovian conditioning. En N. A. Schmajuk & P. C. Holland (Eds.) Occasion Setting: Associative Learning and Cognitions in Animals. pp. 343-382. Washington, DC: American Psychological Association. [ Links ]
Nelson, J. B. (2002). Context specifity of excitation and inhibition in ambiguous stimuli. Learning and Motivation, 33, 284-310. [ Links ]
Nelson, J. B. & Bouton, M. E. (1997). The effects of a context switch following serial and simultaneous feature-negative discriminations. Learning and Motivation, 28, 56-84. [ Links ]
Pavlov, I. P. (1927) Lectures on conditioned reflexes: Twenty-five years of objective study of the higher nervous activity (behavior) of animals. (W. H. Gantt, Traducción.) New York: International Publishers. [ Links ]
Pearce, J. M. (1987). A model for stimulus generalization in pavlovian conditioning. Psychological Review, 94, 1, 61-73. [ Links ]
Pinel, J. P. J. (2001). Biopsicología. España: Prentice-Hall. [ Links ]
Ramos, B., Siegel, S. & Bueno, J. L. (2002). Occasion setting and drug tolerance. Integrative Physiological & Behavioral Science, 37, 165-177. [ Links ]
Ramsay, D. S. & Woods, S. C. (1997). Biological consequences of drug administration: Implications for acute and chronic tolerance. Psychological Review, 104, 170-193. [ Links ]
Rescorla, R. A. (1986a). Facilitation and excitation. Journal of Experimental Psychology: Animal Behavior Processes, 12, 325-332. [ Links ]
Rescorla, R. A. (1986b). Extinction of facilitation. Journal of Experimental Psychology: Animal Behavior Processes, 12, 16-24. [ Links ]
Rescorla, R. A. (1999). Associative changes in elements and compounds when the other is reinforced. Journal of Experimental Psychology: Animal Behavior Processes, 25, 247-255. [ Links ]
Rodríguez B. I., Craske M. G., Mineka S. & Hladek D. (1999). Context-specificity of relapse: effects of therapist and environmental context on return of fear. Behaviour Research and Therapy, 37, 845-62. [ Links ]
Ross, R. T. & Holland, P. C. (1981). Conditioning of simultaneous and serial feature positive discrimination. Animal, Learning & Behavior, 9, 293-303. [ Links ]
Schachtman, T., Threlkeld, R. & Meyer, K. (2000). Retention of conditioned inhbition produced by extinction.Learning and Motivation, 31, 283-300. [ Links ]
Schmajuk, N. A. & Holland, P. C. (1998), Occasion Setting. Associative Learning and Cognition in Animals. Washington, DC: American Psychological Association. [ Links ]
Siegel, S. (1999a). Glucose enhancement of tolerance to morphine and ethanol in rats. Psychobiology, 27, 372-376. [ Links ]
Siegel, S. (1999b). Drug anticipation and drug addiction: The 1998 H. David Archibald lecture. Addiction, 94, 1113-1124. [ Links ]
Siegel, S. (2005). Drug tolerante, drug addiction and drug anticipation.Current Directions in Psychological Science, 14 (6),296-300. [ Links ]
Siegel, S., Baptista, M., Kim, J., McDonald, R. & Weise-Kelly, L. (2000). Pavlovian psychopharmacology: the associative basis of tolerance. Experimental & Clinical Psychopharmacology, 8, 276-293. [ Links ]
Siegel, S. & Larson, S. J. (1996). Disruption of tolerance to the ataxic effect of etanol by a novel stimulus. Pharmacology, Biochemistry & Behavior, 55, 125-130. [ Links ]
Siegel, S. & Ramos, B. M. (2002). Applying laboratory research. Drug anticipation and the treatment of drug addiction.Experimental and Clinical Psychopharmacology, 10, 162-183. [ Links ]
Swartzentruber, D. (1995). Modulatory mechanisms in pavlovian conditioning. Animal, Learning & Behavior, 23, 123-143. [ Links ]
van Wijk, E. P. A., Maes, J. H. R. & Vossen, J. M. H. (1997). Competition between a simultaneous and a serial feature in appetitive feature-positive and feature-negative discriminations. The Quarterly Journal of Experimental Psychology, 50B(1), 17-39. [ Links ]
Walter, T. A. & Riccio, D. C. (1983). Overshadowing effects in the stimulus control of morphine analgesic tolerance. Behavioral Neuroscience, 97, 658-662. [ Links ]
White, A. M., Roberts, D. C. & Best, P. J. (2002). Context-specific tolerance to the ataxic effects of alcohol. Pharmacology, Biochemistry and Behavior, 72(1-2), 107-110. [ Links ]
Williams, D. (1995). Forms of inhibition in animal and human learning. Journal of Experimental Psychology: Animal Behavior Processes, 21, 129-142. [ Links ]
Recepción: Diciembre de 2006
Aceptación final: Enero de 2008
1 Los autores agradecen a la Dra. MARÍA DE LOS ÁNGELES SAAVEDRA y al Dr. JORGE LUZORO G. por sus comentarios a versiones anteriores del presente artículo. El orden de autoría del presente artículo ha sido determinado por el orden alfabético de los apellidos de los autores, sin guardar relación alguna con el trabajo realizado por cada uno de ellos. Parte de los resultados presentados en este artículo fueron previamente expuestos en la 77th Annual Meeting of the Eastern Psychological Association (Baltimore, 2006).
2 Correspondencia: RONALD BETANCOURT, MARION INOSTROZAO MARIO A. LABORDA, Departamento de Psicología, Universidad de Chile, Avenida Capitán Ignacio Carrera Pinto 1045, 4to piso, Ñuñoa, Santiago, Chile. Correos electrónicos: rbetancourt@progranja.cl, mainostr@uchile.cl, mlaborda@uchile.cl
