










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTIIndicadores
Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.4 n.1-2 São Paulo 1993
ARTIGOS ORIGINAIS
Neuropsicologia da memória humana
Neuropsychology of human memory
Maria Gabriela Menezes Oliveira; Orlando F. A. Bueno
Departamento de Psicobiologia - Escola Paulista de Medicina
RESUMO
A síndrome amnésica do lobo temporal medial tem por característica básica uma severa amnésia anterógrada, eventualmente acompanhada de amnésia retrógrada. Seus efeitos não se restringem a uma modalidade sensorial ou qualidade do material. Outras capacidades cognitivas, memória imediata, assim como a capacidade de aprender e reter tarefas que não exijam a lembrança de eventos específicos, permanecem intactas. Uma das estruturas críticas envolvidas é o hipocampo. Na amnésia de Korsakoff há lesões talâmicas e dos corpos mamilares, mas a destruição de conexões destas estruturas pode ser também importante. Tanto na amnésia do lobo temporal medial, como em Korsakoff é afetada a memória declarativa (ou explícita) mas não a de procedimento (ou implícita). Estruturas cerebrais que embasam a memória implícita ainda não foram identificadas de todo. O estriado pode estar envolvido; o efeito de pré-ativação possivelmente ocorre no neocórtex; o papel do cerebelo no condicionamento clássico de respostas motoras foi demonstrado.
Descritores: Memória. Neuropsicologia. Cérebro. Amnésia. Distúrbios cerebrais.
ABSTRACT
The basic feature of medial temporal lobe amnesia is a severe anterograde amnesia, eventually followed by retrograde amnesia. It is not restricted to any specific sensory modality nor to verbal or non-verbal aspects. Other cognitive capacities (speech, etc.), short-term memory, and the ability to learn and remember tasks which do not require recollection of specific experiences are preserved. One of the critical structures involved is the hippocampus. Korsakoff 's syndrome is caracterized by damage of the thalamic medial dorsal nucleus and the mammilary bodies. The destruction of fiber connections running through the mammillothalamic tract and the internal medullary lamina of thalamus may be even more important than nuclear mass loss . Both in Korsakoff as in medial temporal lobe amnesia, there is an impairment of declarative (or explicit) memory but not of procedural (or implicit) memory. Brain structures involved in implicit memory are not yet well identified. The striatum may be a substrate for procedural memory; priming possibly occurs at the posterior cerebral cortex; animal studies have shown the role of cerebellum in the classical conditioning of motor responses.
Index terms: Memory. Neuropsychology. Brain. Amnesia. Brain disorders.
O termo neuropsicologia refere-se, originalmente, a investigações de desordens de percepção o, memória, linguagem, pensamento, emoção o e ação em pacientes neurológicos (Shallice, 1988), ampliado para abarcar o resultado de numerosos estudos complementares em animais de laboratório, como se depreende do uso que dele fazem Squire e Butters (1984). No seu uso original ele designava o trabalho conjunto de psicólogos, neurologistas e neurocirurgiões, e no seu uso mais generalizado -e este é o aspecto fundamental que necessita ser destacado - ele continua a refletir o trabalho em cooperação o de psicólogos, neurologistas, neurocirurgiões, neuroanatomistas, neurofísiologistas e outros neurocientistas. é nossa crença que somente este trabalho cooperativo pode fazer avançar o conhecimento a respeito do cérebro e do comportamento. O que se segue é uma tentativa de resumo de resultados obtidos nas últimas duas ou três décadas num campo particular da neuropsicologia, a neuropsicologia da memória humana, que neste período conheceu um crescimento espetacular.
Amnésia do lobo temporal medial
A década de 50 foi um marco na história da neuropsicologia da memória. Em 1954 o neurocirurgião Scoville descreveu o caso do paciente H.M., por ele operado em 1953, o qual começou a apresentar uma grave e inesperada deficiência de memória logo após a ablacão bilateral do lobo temporal medial. Brenda Milner e colaboradores publicaram vários trabalhos relatando os resultados obtidos em testes formais de memória e inteligência executados neste paciente (Milner, Corkin & Teuber, 1968; Scoville & Milner, 1957). Estes resultados confirmaram amplamente os relatos informais de seus familiares, de médicos e de pesquisadores, os quais constataram que H.M. não conseguia recordar nenhum evento pessoal ou público ocorrido após a cirurgia (amnésia anterógrada), além de sempre subestimar a própria idade e não reconhecer amigos aos quais havia sido apresentado após o ano de 1953.
Apesar deste profundo e persistente quadro amnésico, o paciente H.M. tinha uma inteligência superior, era capaz de estabelecer diálogos normalmente, desde que não fosse distraído, e tinha preservada sua memória para eventos remotos, sendo, porém, incapaz de lembrar com acuidade eventos ocorridos até cerca de três anos antes da cirurgia (amnésia retrógrada). Em testes de percepção o seu desempenho era normal, indicando que o prejuízo observado não era devido à incapacidade de registrar novas informações (Milner et al., 1968; Scoville & Milner, 1957). A memória de curto prazo de H.M. achava-se preservada (Milner et al., 1968), assim como a sua capacidade de adquirir novas habilidades motoras e cognitivas (Corkin, 1968). Além disso, ele também conseguia aprender, e reter na memória por algum tempo, o trajeto de labirintos visuais e tácteis muito simples (onde o número de pontos de escolha a serem lembrados não excedesse a capacidade da memória de curto prazo), embora a aquisição o destas tarefas de labirinto fosse extremamente lenta (Milner et al., 1968). Foi demonstrado também que H.M. era capaz de se beneficiar do efeito de pré-ativação (Corkin, 1984), efeito no qual a performance do indivíduo pode ser facilitada ou enviesada por informações apresentadas previamente, sem que o sujeito se dê conta disso. O paradigma básico dos estudos de pré-ativação (priming) é aquele imaginado por Warrington e Weiskrantz (1970), que consiste em apresentar uma lista de palavras e subseqüentemente testar os sujeitos através de dois testes. Um dos testes é de reconhecimento, no qual a tarefa é identificar no meio de várias palavras quais foram apresentadas anteriormente. O outro teste é de completamente de palavras, no qual são dadas as letras iniciais para que se forme a palavra completa. H.M., assim como outros amnésicos, tipicamente completava as palavras de acordo com a lista apresentada previamente, mas não conseguia reconhecer as palavras que constavam desta lista. O paciente H.M. foi exaustivamente estudado ao longo dos anos e muito se sabe sobre as características de sua amnésia, mas o seu não é, de modo algum, o único caso descrito na literatura. Em 1958 Penfield e Milner descreveram vários casos de cirurgia de ablação unilateral do lobo temporal medial, que incluía a amígdala e boa parte do hipocampo. Destes pacientes, dois desenvolveram um quadro amnésico semelhante ao observado em H.M.. Estudos post-mortem mostraram que o hipocampo do hemisfério cerebral não operado se apresentava necrótico e provavelmente inoperante (Penfield & Mathieson, 1974). Portanto, a síndrome amnésica só aparecia quando havia comprometimento bilateral desta estrutura. Estes achados mostraram um claro envolvimento do hipocampo no processamento de informações mnemônicas, o que já havia sido sugerido em 1900 por Bechterew, que descreveu alterações neuropatológicas no lobo temporal medial de uma pessoa que havia apresentado problema de memória durante a sua vida (citado por Zola-Morgan, Squire & Amaral, 1986). Síndrome amnésica semelhante pode ser observada em pacientes pós-encefalíticos, nos quais há lesões extensas do lobo temporal medial, incluindo hipocampo, amígdala e uncus (Damasio, Eslinger, Damasio, Van Hoesen & Cornell, 1985). O hipocampo também tem sido responsabilizado pela amnésia que se manifesta após oclusão da artéria cerebral posterior (Benson, Marsden & Meadows, 1974) ou após isquemia cerebral (Volpe & Hirst, 1983).
Síndrome amnésica, amnésia global ou amnésia orgânica
A síndrome amnésica é definida por algumas características positivas - das quais a mais importante é uma severa amnésia anterógrada -concomitantes é ausência de outros déficits cognitivos (linguagem, raciocínio, capacidade intelectual, habilidades sociais, percepção, etc). Amnésia retrógrada pode aparecer, porém em graus variados e restrita aos anos, meses ou dias que antecederam o agente amnésico. A memória para eventos remotos se conserva intacta, todavia. Pacientes com amnésia do lobo temporal medial apresentam capacidade preservada de aprender tarefas que envolvam apenas habilidades motoras, perceptuais ou cognitivas (como as tarefas de perseguição o de um ponto giratório, de traçado bimanual, e de escrever ou ler palavras invertidas por um espelho, ou como solucionar o jogo da Torre de Hanói e congêneres), ou seja, que não exijam a lembrança de eventos específicos. Um bom exemplo para ilustrar esta dissociação o apresentada por pacientes amnésicos encontra-se no experimento realizado por Cohen e Squire (1980), no qual os sujeitos liam tríades de palavras invertidas como num espelho, durante três dias. Com o treino, o tempo de leitura, tanto de sujeitos normais como de amnésicos, caía igualmente, revelando que a habilidade de ler palavras invertidas estava sendo bem aprendida. Esta habilidade persistiu por três meses, pelo menos. Porém, testes de reconhecimento revelaram que os amnésicos se esqueciam muito mais rapidamente das palavras específicas que foram apresentadas durante o treino. A esta dissociação o os autores chamaram sugestivamente de " saber como" e " saber o quê".
O condicionamento clássico da resposta de piscar o olho é outro exemplo de aprendizagem e retenção o de longo prazo preservados em pacientes amnésicos (Weiskrantz & Warrington, 1979).
A memória de curto prazo, avaliada através do desempenho em testes de capacidade (span) de memória ou através do efeito de recência em testes de recordação livre de palavras, também não é afetada nestes pacientes (Baddeley & Warrington, 1970; Brooks & Baddeley, 1976).
A síndrome amnésica não se restringe a uma modalidade sensorial específica, nem é qualidade do material apresentado (verbal, seja escrito ou falado, e não-verbal), sendo, por isso mesmo, também chamada de amnésia global (Cohen, 1984; Rozin, 1976; Squire, 1982b). No entanto, lesões focais do lobo temporal direito (não-dominante) e do esquerdo revelaram, respectivamente, que o hipocampo direito desempenha papel crítico na memorização o de material difícil de verbalizar (como rostos e sons não-familiares, padrões visuais abstratos) e de material disposto espacialmente, e que o hipocampo esquerdo está mais relacionado com a memória de material verbal (Jones-Gotman, 1986; Jones-Gotman & Milner, 1978; Smith & Milner, 1981).
A síndrome amnésica ou amnésia global é também conhecida como amnésia orgânica. Este termo refere-se ao fato da amnésia provir de uma alteração o orgânica resultante de cirurgia, ruptura de aneurismas, anóxia, tumores cerebrais, infecções, lesões cerebrais por penetração, traumatismo craniano, etc.
A estrutura crítica: o hipocampo?
Desde o trabalho pioneiro de Scoville e Milner (1957) o hipocampo tem sido considerado a estrutura crítica no desencadeamento da síndrome amnésica associada ao lobo temporal medial, pois observa-se uma correlação entre o grau de prejuízo e a magnitude do comprometimento do hipocampo. Alguns autores, no entanto, questionaram a conclusão de que a ausência do hipocampo seria necessária e suficiente para causar o quadro amnésico e lançaram algumas explicações alternativas. Horel (1978) propôs que a estrutura crítica para as funções mnemônicas seria o pedúnculo temporal (um sistema de fibras que conectam a amígdala e o córtex temporal a outras estruturas subcorticais). Segundo o autor, esta região necessariamente seria atingida no procedimento cirúrgico utilizado para remover o lobo temporal medial. Estudos com primatas não-humanos comprovaram posteriormente que a lesão do pedúnculo temporal não provoca prejuízo em testes de memória. Aparentemente este tipo de lesão leva a um déficit de aprendizagem em tarefas onde se exige a discriminação entre padrões visuais diferentes (Zola-Morgan & Squire, 1984; Zola-Morgan, Squire & Mishkin, 1982). Este déficit provavelmente se deve a um problema no processamento de informações visuais e não de memória propriamente (Zola-Morgan & Squire, 1984).
Mishkin (1978, 1982), por sua vez, não nega a importância do hipocampo, mas sustenta que o extraordinário déficit de memória observado em pacientes como H.M. é devido a lesões conjuntas de hipocampo e amígdala. Mishkin verificou, em macacos, que a lesão bilateral conjunta de hipocampo e amígdala produz um prejuízo maior que lesões bilaterais de qualquer uma destas duas estruturas feitas isoladamente. Macacos com este tipo de lesão conjunta apresentavam prejuízos em tarefas nas quais pacientes amnésicos também mostravam dificuldades (Squire, Zola-Morgan & Chen, 1988; Zola-Morgan & Squire, 1985).
Zola-Morgan, Squire e Amaral (1989), entretanto, puseram em dúvida as conclusões tiradas por Mishkin em relação à contribuição da amígdala na gênese da síndrome amnésica. Segundo estes autores, lesões controladas em macacos, que realmente só atinjam a amígdala, poupando o cortéx adjacente (conseguidas através de técnicas estereotóxicas), não apenas não produzem efeito em várias tarefas de memória, como também não potencializam os efeitos da lesão hipocampal. Eles sugeriram, então, que o prejuízo observado por Mishkin deve-se ao fato de que as lesões produzidas por ele atingiram, além da amígdala, áreas corticais adjacentes (córtex perirrinal, giro para-hipocampal), e apontam uma provável contribuição destas áreas no prejuízo de memória. E realmente a lesão conjunta bilateral do hipocampo e destas áreas corticais (poupando a amígdala, porém) produziu um efeito deletério maior em testes de memória do que a lesão apenas do hipocampo (Squire & Zola-Morgan, 1991).
Revendo as evidências disponíveis na literatura sobre casos de lesões na amígdala, além de estudos de registro e estimulação desta estrutura em seres humanos, Sarter e Markowitsch (1985) propuseram um papel mais restrito para a amígdala: ela seria responsável pelos eventos mnemônicos que têm um significado emocional para o sujeito.
O caso do paciente R.B. foi o primeiro relatado no qual uma lesão restrita ao hipocampo produziu amnésia anterógrada, sem sinais de outros déficits cognitivos ou de amnésia retrógrada significativa. Este paciente ficou amnésico devido a um episódio isquêmico, e após sua morte o exame histológico do cérebro mostrou que havia lesão bilateral restrita ao campo CA1 do hipocampo (Zola-Morgan, Squire & Amaral, 1986). É importante notar, entretanto, que o déficit observado no paciente R.B. não era tão severo quanto o observado no paciente H.M.
Estes achados indicam que a formação hipocampal (entendida como o conjunto que inclui o giro denteado, o corno de Amon, o subículo e o córtex entorrinal) e mais o córtex perirrinal e para-hipocampal desempenham papel de fundamental importância na memória, presumivelmente devido às conexões recíprocas entre estas estruturas e o neocórtex associativo posterior. Em consonância com esta idéia está o relato de perda de memória em primatas devida à desconexão entre o córtex inferotemporal (área TE) e o sistema límbico (Mishkin, 1982).
Amnésia diencefálica
A síndrome descrita primeiramente por Korsakoff no século passado (citações em Lhermitte & Signoret, 1972; Victor, Adams & Collins, 1971), é outra patologia que apresenta a perda de memória como um de seus sinais mais marcantes. Esta síndrome é causada por deficiência de tiamina, geralmente secundária a desnutrição associada com alcoolismo crônico, com possível contribuição dos efeitos tóxicos do álcool per se (Butters, 1985; Lishman, 1981; Victor et al., 1971). Em pacientes portadores desta doença constatou-se, através de autópsia, que o lobo temporal medial se apresenta intacto. A lesão cerebral centra-se principalmente em estruturas do diencéfalo, sendo que as mais atingidas são o núcleo medial dorsal do tálamo e os corpos mamilares no hipotálamo (Mair, Warrington & Weiskrantz, 1979; Victor et al., 1971).
Pacientes de Korsakoff apresentam amnésia global anterógrada persistente, semelhante à encontrada em pacientes com lesão bilateral do lobo temporal medial (Butters, 1985; Mair et al., 1979; Parkin, 1984). Amnésia retrógrada também é uma característica marcante, sendo mais difícil para os pacientes se lembrarem de eventos que ocorreram logo antes do início da doença do que de eventos mais remotos, como por exemplo da infância (Butters, 1985; Shimamura & Squire, 1986). Testes formais de amnésia retrógrada, como o teste de personalidades famosas de Albert, comprovaram a existência deste gradiente de dificuldade (Albert, Butters & Levin, 1979). Além da amnésia anterógrada e retrógrada, os pacientes com Korsakoff apresentam alguns outros déficits cognitivos, principalmente em tarefas que envolvam resolução de problemas, atribuídos geralmente a atrofia de regiões neocorticais (Butters, 1985). Assim como a amnésia do lobo temporal, a amnésia diencefálica também é de natureza seletiva, pois estes pacientes são capazes de se beneficiarem do efeito de pré-exposição (Shimamura, 1986) e de aprenderem habilidades percepto-motoras (Brooks & Baddeley, 1976), apesar da severa amnésia anterógrada.
Não existe consenso na literatura a respeito do comprometimento da memória de curto prazo na síndrome de Korsakoff. Baddeley e Warrington (1970) e Brooks e Baddeley (1976) notaram que, em vários testes de memória de curto prazo, estes pacientes não apresentaram alteração. Entre estes testes estava a tarefa de memória de curto prazo de Brown-Peterson, na qual Baddeley e Warrington (1970) observaram um esquecimento normal em seus pacientes amnésicos. Cermak, Butters e Good-glass (1971) e Butters, Lewis, Cermak e Goodglass (1973) obtiveram em tarefa semelhante resultados opostos ao de Baddeley e Warrington (1970). Butters e Cermak (1974) atribuíram este resultado conflitante principalmente ao fato de que os outros autores utilizaram pacientes de diversas etiologias (não apenas com a doença de Korsakoff), e também ao fato de que a metodologia por eles empregada permitia que os pacientes tivessem um tempo para repetir mentalmente a informação apresentada, facilitando, assim, a execução da tarefa.
Pacientes com lesões penetrantes confirmam o envolvimento de estruturas do diencéfalo na memória. O paciente N.A. ficou amnésico após sofrer um acidente com um florete em miniatura que penetrou em sua narina e atingiu estruturas do diencéfalo. Depois do acidente, ele não conseguia relembrar a maioria dos eventos correntes pouco tempo após eles terem acontecido (amnésia anterógrada), e nem de eventos que ocorreram logo antes da lesão (amnésia retrógrada). Sua amnésia não era acompanhada de qualquer outro déficit intelectual ou perceptual (Teuber, Milner & Vaughan, 1968). Inicialmente achava-se que a lesão se restringia ao núcleo medial dorsal esquerdo do tálamo (Squire & Moore, 1979), porém, dez anos após, Squire, Amaral, Zola-Morgan, Kritchevsky e Press (1989) verificaram, através de exames com ressonância magnética nuclear, que outras estruturas como os corpos mamilares, o trato mamilotalâmico esquerdo, entre outras, também foram atingidas.
Outro paciente (B.J.) tornou-se amnésico após um acidente com um taco de bilhar que, à semelhança de N.A., penetrou por sua narina atingindo o cérebro. Ressonância magnética nuclear mostrou que a lesão atingiu principalmente os corpos mamilares, permanecendo o tálamo intacto. A amnésia apresentada por este paciente, entretanto, restringia-se a materiais verbais (Dusoir, Kapur, Byrnes, Mackinstry & Hoare, 1990).
Embora seja comum atribuir o prejuízo de memória observado em pacientes com a doença de Korsakoff a lesões de estruturas como o núcleo medial dorsal do tálamo e/ou corpos mamilares (Mair, Warrington & Weiskrantz, 1979; Victor, Adams & Collins, 1971), outros autores sugerem que, mais importante que a destruição das massas nucleares mencionadas, seria a lesão do trato mamilotalâmico e da lâmina medular interna do tálamo (Cramon, Hebel & Schuri, 1985; Graff-Radford, Tranel, Van Hoesen & Brant, 1990; Markowitsch, 1988). O trato mamilotalâmico conecta os corpos mamilares aos núcleos do grupo anterior do tálamo. Pela lâmina medular interna trafegam fibras que conectam o núcleo medial dorsal do tálamo ao córtex prefrontal, córtex cíngulado anterior e amígdala. A dificuldade em se chegar a uma conclusão definitiva a respeito de qual estrutura diencefálica seria crítica no desenvolvimento da síndrome amnésica reside no fato de que a maioria das lesões observadas não se restringe a um determinado grupo de fibras ou núcleos (Markowitsh, 1988).
Em resumo, do ponto de vista neuroanatômico, dois grandes tipos de amnésia global estão amplamente documentados: a amnésia temporal e a amnésia diencefálica. Controvérsias a respeito das estruturas envolvidas no primeiro caso parecem satisfatoriamente resolvidas; na amnésia diencefálica ainda não.
Amnésia do lobo temporal e amnésia diencefálica: uma única entidade?
Dada a interconexão entre o hipocampo e os corpos mamilares através da fímbria-fórnix, entre os corpos mamilares e núcleos do grupo anterior do tálamo através do trato mamilotalâmico e entre a amígdala e o núcleo medial dorsal do tálamo, alguns pesquisadores consideram que qualquer interrupção bilateral deste circuito provoca a síndrome amnésica, que seria única em essência (Baddeley & Warrington, 1970; Warrington & Weiskrantz, 1974; Weiskrantz, 1987).
A identidade entre a amnésia do lobo temporal medial e a amnésia diencefálica é controversa, porém. Embora existam muitas semelhanças entre os dois quadros, algumas diferenças também foram observadas. Por exemplo: ao contrário dos pacientes com lesão de lobo temporal medial, pacientes com Korsakoff geralmente não percebem que têm problemas de memória e confabulam; também a amnésia retrógrada apresentada por estes últimos parece ser mais notável (Parkin, 1984). Diferença importante apontada por vários autores (Lhermitte & Signoret, 1972; Parkin, 1984; Rozin, 1976; Squire, 1982a) é que pacientes com a síndrome de Korsakoff freqüentemente apresentam sintomas que são tópicos de danos ao lobo frontal. Estas diferenças, segundo Parkin (1984), sugerem que a amnésia do lobo temporal e a amnésia diencefálica são entidades distintas. Advogando a favor da identidade entre amnésia do lobo temporal e diencefálica, porém, Squire (1982a) argumenta que os déficits cognitivos adicionais apresentados pelos pacientes com Korsakoff não são essenciais para a caracterização da síndrome amnésica, mas, influenciam a forma final do quadro apresentado. Em especial, este autor realça o comprometimento do lobo frontal nestes pacientes. Alguns outros autores, porém, acreditam na existência de mais uma diferença entre a amnésia temporal e a diencefálica, diferença esta que dificilmente poderia ser atribuída a outro déficit cognitivo que não ao problema de memória. Mais especificamente estes dois tipos de amnésia difeririam quanto à velocidade de esquecimento de materiais armazenados na memória de longo prazo. A velocidade de esquecimento de pacientes com lesão de diencéfalo, como N.A. e doentes de Korsakoff seria normal, enquanto que pacientes com lesão do lobo temporal medial esqueceriam anormalmente rápido (Lhermitte & Signoret, 1972; Mayes & Meudell, 1984; Parkin, 1982). Entretanto, em trabalho recente, Mckee e Squire (1992) não observaram diferenças estatisticamente significativas nas curvas de esquecimento de pacientes com lesão de lobo temporal medial e pacientes com lesão de diencéfalo.
Quantos tipos de memória existem?
Seja única ou múltipla em sua origem, a síndrome amnésica é de natureza seletiva, pois, embora os pacientes amnésicos, tanto diencefálicos como temporais, não consigam se lembrar da maioria dos eventos que experienciam no dia-a-dia, muitas funções de aprendizagem e memória estão preservadas, como por exemplo, a capacidade de adquirirem habilidades motoras, cognitivas e perceptuais e a capacidade de se beneficiarem do efeito de pré-exposição. Este fato levanta a questão fundamental a respeito da existência de diferentes sistemas de memória. Existiria pelo menos um segundo sistema de memória, independente de estruturas como o lobo temporal medial e o diencéfalo, através do qual seriam processadas aquelas funções preservadas. Vários pesquisadores, de fato, têm argumentado a favor da existência de múltiplos sistemas de memória (Cohen, 1984; Poulos & Wilkinson, 1984; Squire, 1986; Squire & Zola-Morgan, 1988;Tulving, 1983).
A noção de que a memória poderia ser dividida em componentes é antiga. Ainda no século passado William James1 (citado por Waugh & Norman, 1965) definiu, por introspecção, os conceitos de memória primária e memória secundária. A memória primária se estenderia por um curto prazo de tempo e dela fariam parte eventos pertencentes ao "presente psicológico" que após serem percebidos ainda não deixaram a consciência. A memória secundária pertenceria ao "passado psicológico" e já esteve ausente da consciência. Posteriormente estes conceitos evoluíram em memória de curto prazo (ou memória imediata) e memória de longo prazo, respectivamente. Atualmente existe alguma evidência experimental que permite diferenciar memória de curto prazo (retenção de itens por apenas alguns segundos) de memória de longo prazo (retenção de itens por intervalos maiores) (Baddeley & Warrington, 1970). A memória de curto prazo tem capacidade limitada (máximo de sete a nove itens) e o material nela armazenado apresenta uma taxa de esquecimento bem rápida (em torno de segundos). Apesar desta rápida taxa de esquecimento, a informação pode ser mantida por mais tempo na memória através de reverberação (rehearsal) (Atkinson & Shiffrin, 1971). Estudos neuropsicológicos têm confirmado e estendido a noção de memória de curto prazo, sob a rubrica de memória operacional (working memory) (Baddeley, 1986). Na síndrome amnésica, aparentemente, a memória de curto prazo encontra-se preservada, embora alguma discussão ainda exista com relação ao possível déficit deste tipo de memória apresentado por pacientes com Korsakoff.
Além da distinção tradicionalmente feita entre memória de curto e longo prazo, os estudos com pacientes amnésicos, relatados acima, levaram à distinção entre memória declarativa e memória de procedimento, dissociadas tanto funcionalmente como anatomicamente (Cohen, 1984; Squire, 1986).
A memória declarativa é a habilidade de armazenar e recordar ou reconhecer conscientemente fatos e acontecimentos; a lembrança pode ser declarada, isto é, trazida à mente verbalmente como uma proposição, ou não verbalmente como uma imagem (Saint-Cyr, Taylor & Lang, 1988; Squire, 1986). Seria este tipo de memória que estaria afetada em pacientes amnésicos.
A memória de procedimento é a capacidade de adquirir gradualmente uma habilidade percepto-motora ou cognitiva através da exposição repetida a uma atividade específica que segue regras constantes. Esta capacidade é implícita e independe da consciência, só podendo ser aferida através do desempenho do paciente (Saint-Cyr et al., 1988; Squire, 1986). Pacientes amnésicos teriam este tipo de memória preservada.
A aprendizagem de novas habilidades é um exemplo do que Schacter (1987a, 1987b) denomina memória implícita, isto é, conhecimento que para ser demonstrado não requer referência explícita a um episódio particular. A memória implícita contrapõe-se à memória explícita, que implica na recordação consciente de experiências vividas. O termo memória implícita, por ser mais genérico que memória de procedimento, permite acomodar melhor, nesta categoria, o efeito de pré-ativação que se apresenta intacto em pacientes amnésicos e pode ser observado após uma única exposição ao estímulo. Cohen (1984) considera a memória declarativa e a de procedimento mais ou menos como sinônimos de memória explícita e memória implícita, respectivamente.
Para Mishkin e Petri (1984) o produto do sistema de memória de procedimento é um hábito, e não uma memória propriamente dita, produto, esta última, do sistema declarativo. Com esta distinção entre hábito e memória pretendem estes autores remeter o sistema de aprendizagem de procedimento aos estudos clássicos do behaviorismo (aprendizagem de estímulo-resposta). Lembremo-nos de que no sistema de Hull o conceito de hábito ocupa posição central em sua teoria da aprendizagem (Hilgard, 1975).
Tulving (1983) propôs ainda uma outra distinção conceituai entre o que ele chama de memória episódica e memória semântica. O termo memória episódica refere-se ao armazenamento de informações pessoais que permite ao indivíduo se lembrar de eventos dos quais participou no passado. A memória semântica capacita o indivíduo a adquirir conhecimento factual do mundo, ou seja, conhecimento impessoal de fatos relevantes.
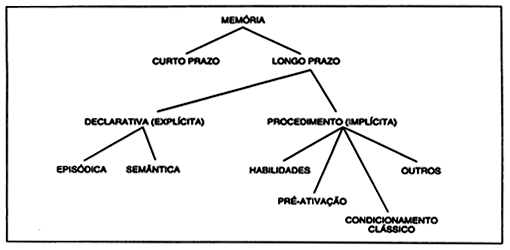
Estas diversas variantes de conceituação e principalmente de terminologia têm gerado certa confusão na literatura. Uma tentativa de estabelecer uma taxonomia para a memória, tentando acomodar várias destas divisões em um sistema coerente levou ao seguinte quadro, modificado de Squire (1986):
A memória declarativa ou explícita incluiria as memórias episódica e semântica. A memória de procedimento ou implícita incluiria habilidades (motoras, perceptuais e cognitivas), efeito de pré-ativação, condicionamento clássico, habituação, sensitização e tudo que foi aprendido mas que só pode ser aferido através do desempenho (Squire, 1986). Estes diversos tipos de memória pertenceriam à memória de longo prazo. Este quadro, entretanto, não é universalmente aceito. Mishkin e Petri (1984), por exemplo, consideram que o seu sistema de memória estaria relacionado com memória episódica ou declarativa e utilizam os termos memória semântica e de procedimento como pertencentes a um mesmo sistema: ao sistema de hábito. Parte pelo menos das dificuldades deste sistema de classificação deriva da ambigüidade dos conceitos de memória episódica e memória semântica.
Estruturas cerebrais envolvidas com memória implícita
As estruturas cerebrais envolvidas na memória implícita certamente abarcam regiões muito mais extensas do sistema nervoso e ainda não foram bem identificadas. Além disso, o conceito de memória implícita engloba vários tipos de memória que, embora satisfaçam todos os critérios de independência da consciência, são dessemelhantes entre si em outros aspectos.
O envolvimento do corpo estriado na memória de procedimento foi sugerido por Mishkin, Malamut e Bachevalier (1984). Segundo eles, o neoestriado, principalmente o núcleo caudado, teria um papel importante no estabelecimento do que ele chama de hábito. Por suas conexões anatômicas, o neoestriado seria uma estrutura apropriada para promover ligação entre estímulos e ações, pois recebe projeções de várias áreas do córtex, incluindo áreas sensoriais, e envia fibras a estruturas subcorticais que fazem parte do sistema de controle dos movimentos (Mishkin & Appenzeller, 1987).
Algumas evidências diretas deste envolvimento já existem. Na tentativa de melhor caracterizar demências de etiologias diversas, Martone, Butters e Payne (1984) compararam o desempenho de pacientes portadores do mal de Huntington, de Korsakoff e indivíduos normais no teste de leitura de palavras invertidas, no qual tríades de palavras eram apresentadas de forma que parecessem projetadas em um espelho. O procedimento foi determinado de forma que os indivíduos fossem submetidos a uma tarefa de aprendizagem de habilidade perceptual (teste de memória de procedimento) e a uma tarefa de reconhecimento de palavras já vistas (teste de memória declarativa). Na primeira tarefa os experimentadores mediam o tempo que o indivíduo demorava para ler cada tríade. Na segunda tarefa, os sujeitos deviam apontar, no meio de uma lista, as palavras apresentadas anteriormente. Os resultados mostraram uma dupla dissociação no desempenho de pacientes com Huntington e Korsakoff, ou seja, os pacientes com Korsakoff tiveram um desempenho pobre na tarefa de reconhecimento mas aprenderam tão eficientemente quanto indivíduos normais a tarefa de ler as palavras invertidas. O desempenho dos indivíduos com Huntington foi inferior ao desempenho dos controles normais nesta última tarefa. Na tarefa de reconhecimento, entretanto, seu desempenho foi normal. O mal de Huntington é uma doença transmitida geneticamente que resulta em uma atrofia progressiva dos gânglios da base. Os sintomas mais comuns são a presença de movimentos coreiformes e de um processo de demenciação. Martone et al. (1984) sugeriram então que pacientes cuja demência é resultado de atrofia nos gânglios da base, têm problemas mais relacionados à aquisição de conhecimento procedimental. Entretanto, a patologia apresentada por estes pacientes é complexa e os prejuízos cognitivos não se restringem à memória de procedimento. Em verdade, estes pacientes não apresentam desempenho normal em alguns testes de memória declarativa que envolvem recordação livre, mas apresentam, se não desempenho normal, pelo menos próximo ao normal, em tarefas de reconhecimento (Butters, 1985). é possível entretanto que alguns prejuízos observados em pacientes com Huntington sejam devidos a alterações corticais também presentes nestes indivíduos (Saint-Cyr, Taylor & Lang, 1988). Saint-Cyr et al. (1988) encontraram em pacientes com a doença de Parkinson - portadores, portanto, de disfunção dos gânglios basais - prejuízo seletivo numa tarefa cognitiva tida como exemplo de aprendizagem de procedimento (torre de Hanoi simplificada), enquanto que o desempenho em testes de memória declarativa permanecia intacto.
Segundo Squire (1986), o efeito de pré-ativação encontra-se reduzido em doentes de Alzheimer, o que pode ser uma pista para envolver o córtex cerebral com este fenômeno. Alguns outros estudos apontam para o córtex posterior como uma das estruturas responsáveis pela pré-ativação, independentemente de regiões límbico-diencefálicas essenciais para a memória declarativa (Cave & Squire, 1992).
O papel essencial do cerebelo no condicionamento clássico de respostas motoras (piscar o olho) foi estabelecido claramente por Thompson e seus colaboradores, trabalhando com coelhos (Thompson, 1986). Em seres humanos o cerebelo parece desempenhar papel semelhante, pois uma lesão na circuitaria cerebelar de uma paciente provocou um grande déficit na aquisição de condicionamento clássico da resposta de piscar (Solomon, Stowe & Pendlbeury, 1989).
Perspectivas
Estudos neuropsicológicos têm conseguido fazer a identificação de estruturas e conexões cerebrais importantes para a memória, ou mais precisamente, para as memórias, pois a existência de múltiplos sistemas de memória vem sendo atestada por tais estudos.
Em que pese o grande progresso ocorrido na área, muito resta por descobrir e muitos pontos polêmicos precisam ser esclarecidos, como a questão da identidade ou não da amnésia do lobo temporal medial e da amnésia diencefálica.
Grande empenho está sendo colocado atualmente na identificação de estruturas relacionadas ao processamento de informações dos tipos abarcados pelo conceito de memória implícita.
O papel de cada estrutura, isto é, o que cada uma delas faz para tornar possível as funções psicológicas abrangidas sob a rubrica geral de memória, continua quase inteiramente um mistério. Outra questão pouco tocada ainda refere-se à s interações entre as diversas estruturas envolvidas.
REFERÊNCIAS BIBLIOGRÁFICAS
ALBERT, M.S.; BUTTERS, N.; LEVIN, J. Temporal gradients in the retrograde amnesia of patients with alcoholic Korsakoff's disease. Archives of Neurology, v.36, n.4, p.211-6, 1979. [ Links ]
ATKINSON, R.C.; SHIFFRIN, R.M. The control of short-term memory. Scientific American, v.225, n.2, p.82-90, 1971. [ Links ]
BADDELEY, A.D. Working memory. Oxford, Clarendon Press, 1986. [ Links ]
BADDELEY, A.D.; WARRINGTON, E.K. Amnesia and the distinction between long-and short-term memory. Journal of Verbal Learning and Verbal Behavior, v.9, n.2, p.176-89, 1970. [ Links ]
BENSON, D.F.; MARSDEN, C.D.; MEADOWS, J.C. The amnestic syndrome of posterior cerebral artery occlusion. Acta Neurológica Scandinavica, v.50, p. 133-45, 1974. [ Links ]
BROOKS, D.N.; BADDELEY, A.D. What can amnesic patients learn? Neuropsychologia, v. 14, p.111-22, 1976. [ Links ]
BUTTERS, N. Alcoholic Korsakoff s syndrome: some unresolved issues concerning etiology, neuropathology, and cognitive deficits. Journal of Clinical and Experimental Neuropsychology, v.7, n.2, p. 181-210, 1985. [ Links ]
BUTTERS, N.; CERMAK, L.S. The role of cognitive factors in the memory disorders of alcoholic pacients with the Korsakoff syndrome. Annals of the New York Academy of Sciences, v.233, p.61-75, 1974. [ Links ]
BUTTERS, N.; LEWIS, R.; CERMAK, L.S.; GOODGLASS, H. Material-specific memory deficits in alcoholic Korsakoff patients. Neuropsychologia, v.11, n.3, p.291-9, 1973. [ Links ]
CAVE, C.B.; SQUIRE, L.R. Intact and long-lasting repetition priming in amnesia. Journal of Experimental Psychology: Learning, Memory and Cognition, v.18, n.3, p.509-20, 1992. [ Links ]
CERMAK, L.S.; BUTTERS, N.; GOODGLASS, H. The extent of memory loss in Korsakoff patients. Neuropsychologia, v.9, p.307-15, 1971. [ Links ]
COHEN, N.J. Preserved learning capacity in amnesia: evidence for multiple memory systems. In: Squire, L.R.; Butters, N., eds. The neuropsychology of memory. New York, Guilford Press, 1984. p. 83-103. [ Links ]
COHEN, N.J.; SQUIRE, L.R. Preserved learning and retention of pattern-analyzing skill in amnesia: dissociation of knowing how and knowing that. Science, v.210, n.4466, p.207-10, 1980. [ Links ]
CORKIN, S. Acquisition of motor skill after bilateral medial temporal-lobe excision. Neuropychologia, v.6, p.255-65, 1968. [ Links ]
CORKIN, S. Lasting consequences of bilateral medial temporal lobectomy: clinical course and experimental findings in H.M. Seminars in Neurology, v.4, p.249-59, 1984. [ Links ]
CRAMON, D.Y.; HEBEL, N.; SCHURI, U.S. A contribution to the anatomical basis of thalamic amnesia. Brain, v.108, p.993-1008, 1985. [ Links ]
DAMASIO, A.R.; ESLINGER, P.J.; DAMASIO, H.; VAN HOESEN, G.W.; CORNELL, S. Multimodal amnesic syndrome following bilateral temporal and basal forebrain damage. Archives of Neurology, v.42, n.3, p.252-9, 1985. [ Links ]
DUSOIR, H.; KAPUR, N.; BYRNES, D.P.; MACKINSTRY, S.; HOARE, R.D. The role of diencephalic pathology in human memory disorder. Evidence from a penetrating paranasal brain injury. Brain, v.113, p.1695-1706, 1990. [ Links ]
GRAFF-RADFORD, N.R.; TRANEL, D.; VAN HOESEN, G.W.; BRANT, J.B. Diencephalic amnesia. Brain, v.113, p. 1-25, 1990. [ Links ]
HILGARD, E.R. Teorias da aprendizagem. São Paulo, EPU, 1975. p. 150-229. [ Links ]
HOREL, J.A. The neuroanatomy of amnesia: a critique of the hippocampal memory hypothesis. Brain, v.101, p.403-45, 1978. [ Links ]
JONES-GOTMAN, M. Right hippocampal excision impairs learning and recall of a list of abstract designs. Neuropsychologia, v.24, p.659-70, 1986. [ Links ]
JONES-GOTMAN, M.; MILNER, B. Right temporal-lobe contribution to image-mediated verbal learning. Neuropsychologia, v.16, p.61-71, 1978. [ Links ]
LHERMITTE, F.; SIGNORET, J.L. Analyse neuropsychologique et differenciation des syndromes amnèsiques. Revue Neurologique, v.126, p.161-78, 1972. [ Links ]
LISHMAN, W.A. Cerebral disorder in alcoholism: syndromes of impairment. Brain, v.104,p.1-20, 1981. [ Links ]
MCKEE, R.D.; SQUIRE, L.R. Equivalent forgetting rates in long-term memory for diencephalic and media temporal lobe amnesia. Journal of Neuroscience, v.12, p.3765-72, 1992. [ Links ]
MAIR, W.G.P.; WARRINGTON, E.K.; WEISKRANTZ, L. Memory disorder in Korsakoff's psychosis. A neuropathological and neuropsychological investigation of two cases. Brain, v.102, p.749-83, 1979. [ Links ]
MARKOWITSCH, H.J. Diencephalic amnesia: a reorientation towards tracts? Brain Research Reviews, v.13, p.351-70, 1988. [ Links ]
MARTONE, M.; BUTTERS, N.; PAYNE, P. Dissociation between skill learning and verbal recognition in amnesia and dementia. Archives of Neurology, v.41, p.965-70, 1984. [ Links ]
MAYES, A.R.; MEUDELL, P.R. Problems and prospects for research on amnesia. In: Squire, L.R.; Butters, N., orgs. The neuropsychology of memory. New York, Guilford Press, 1984. p. 134-44. [ Links ]
MILNER, B.; CORKIN, S.; TEUBER, H.L. Further analysis of the hippocampal amnesic syndrome: 14-year follow-up study of H.M. Neuropsychologia, v.6, p.215-34, 1968. [ Links ]
MISHKIN, M. Memory in monkeys severely impaired by combined but not by separated removal of amygdala and hippocampus. Nature, v.273, p.297-8, 1978. [ Links ]
MISHKIN, M. A memory system in the monkey. Philosophical Transactions of the Royal Society of London, v. 298, p.85-95, 1982. [ Links ]
MISHKIN, M.; APPENZELLER, T. The anatomy of memory. Scientific American, v.256,p.62-71, 1987. [ Links ]
MISHKIN, M.; MALAMUT, B.; BACHEVALIER, J. Memories and habits: two neural systems. In: Lynch, G.; McGaugh, J.L.; Weinberger, N.M., eds. Neurobiology of learning and memory. New York , Guilford Press, 1984. p. 65-77. [ Links ]
MISHKIN, M.; PETRI, H.L. Memories and habits: some implications for the analysis of learning and retention. In: Squire, L.R.; Butters, N., eds. The neuropsychology of memory. New York, Guilford Press, 1984. p.287-96. [ Links ]
PARKIN, A.J. Amnesic syndrome: a lesion-specific disorder? Cortex, v.20, n.4, p.479-508, 1984. [ Links ]
PARKIN, A.J. Residual learning capability in organic amnesia. Cortex, v.18, n.3, p.417-40, 1982. [ Links ]
PENFIELD, W.; MATHIELSON, G. Memory: autopsy findings and comments on the role of hippocampus in experiential recall. Archives of Neurology, v.31, n.3, p.145-54, 1974. [ Links ]
PENFIELD, W.; MILNER, B. Memory deficit produced by bilateral lesions in the hippocampal zone. Archives of Neurology and Psychiatry, v.79, p.475-97, 1958. [ Links ]
POULOS, C.X.; WILKINSON, D.A. A process theory of remembering: its application to Korsakoff amnesia and a critique of context and episodic-semantic theories. In: Squire, L. R.; Butters, N., orgs. The neuropsychology of memory. New York, Guilford Press, 1984. p.67-82. [ Links ]
ROZIN, P. The psychobiological approach to human memory. In: Rosenzweig, M.R.; Bennet, E.L., orgs. Neural mechanisms of learning and memory. Cambridge, M. I. T. Press, 1976. p.3-48. [ Links ]
SAINT-CYR, J.A.; TAYLOR, A.E.; LANG, A.E. Procedural learning and neostriatal dysfunction in man. Brain, v.111, p.941-59, 1988. [ Links ]
SARTER, M; MARKOWITSCH, H.J. The amygdala's role in human mnemonic processing. Cortex, v.21,p.1-24, 1985. [ Links ]
SCHACTER, D.L. Implicit expressions of memory in organic amnesia: learning of new facts and associations. Human Neurobiology, v.6, p. 107-18, 1987a. [ Links ]
SCHACTER, D.L. Implicit memory: history and current status. Journal of Experimental Psychology: Learning, Memory and Cognition, v.13, n.3, p.501-18, 1987b. [ Links ]
SCOVILLE, W.B. The limbic lobe and memory in man. Journal Neurosurgery, v.11, p.64, 1954. [ Links ]
SCOVILLE, W.B.; MILNER, B. Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery and Psychiatry, v.20, p. 11-21, 1957. [ Links ]
SHALLICE, T. From neuropsychology to mental structure. Cambridge, Cambridge University Press, 1988. [ Links ]
SHIMAMURA, A.P. Priming effects in amnesia: evidence for a dissociable memory function. Quarterly Journal of Experimental Psychology. Sect.A: Human Experimental Psychology, v.38, n.4, p.619-44, 1986. [ Links ]
SHIMAMURA, A.P.; SQUIRE, L.R. Korsakoff's syndrome: a study of the relation between anterograde amnesia and remote memory impairment. Behavioral Neuroscience, v.100, n.2, p.165-70, 1986. [ Links ]
SMITH, M.L.; MILNER, B. The role of the right hippocampus in the recall of spatial location. Neuropsychologia, v.19, n.6, p.781-93, 1981. [ Links ]
SOLOMON, P.R.; STOWE, G.T.; PENDLBEURY, W.W. Disrupted eyelid conditioning in a patient with damage to cerebellar afferents. Behavioral Neuroscience, v.103, n.4, p.898-902, 1989. [ Links ]
SQUIRE, L.R. Comparisons between forms of amnesia: some deficits are unique to korsakoff's syndrome. Journal of Experimental Psychology: Learning, Memory and Cognition, v.8, n.6, p.560-71, 1982a. [ Links ]
SQUIRE, L.R. Mechanisms of memory. Science, v.232, p.1612-9, 1986. [ Links ]
SQUIRE, L.R. The neuropsychology of human memory. Annual Review of Neuroscience, v.5, p. 241-73, 1982b. [ Links ]
SQUIRE, L.R.; AMARAL, D.G.; ZOLA-MORGAN, S.; KRITCHEVSKY, M.; PRESS, G. Description of brain injury in the amnesic pacient N.A. based on magnetic resonance imaging. Experimental Neurology, v.105, p.23-35, 1989. [ Links ]
SQUIRE, L.R.; BUTTERS, N., orgs. The neuropsychology of memory. New York, Guilford Press, 1984. [ Links ]
SQUIRE, L.R.; Moore. Dorsal thalamic lesion in a noted case of human memory disfunction. Annals of Neurology, v.6, p.503-6, 1979. [ Links ]
SQUIRE, L.R.; ZOLA-MORGAN, S. The medial temporal lobe memory system. Science, v.253, p. 1380-6, 1991. [ Links ]
SQUIRE, L.R.; ZOLA-MORGAN, S. Memory: brain systems and behavior. Trends in Neurosciences, v.11, n.4, p.170-5, 1988. [ Links ]
SQUIRE, L.R.; ZOLA-MORGAN, S.; CHEN, K.S. Human amnesia and animal models of amnesia: performance of amnesic patients on tests designed for the monkey. Behavioral Neuroscience, v. 102, p.210-21, 1988. [ Links ]
TEUBER, H.L.; MILNER, B.; VAUGHAN, H.G. Persistent anterograde amnesia after stab wound of the basal brain. Neuropychologia, v.6, p.267-82, 1968. [ Links ]
THOMPSON, R.F. The neurobiology of learning and memory. Science, v.233, p.941-7, 1986. [ Links ]
TULVING, E. Elements of episodic memory. Oxford, Clarendon Press, 1983. [ Links ]
VICTOR, M.; ADAMS, R.D.; COLLINS, G.H. The Wernicke-Korsakoff Syndrome. Philadelphia, F.A. Davis, 1971. [ Links ]
VOLPE, B.T.; HIRST, W. The characterization of an amnesic syndrome following hypoxic ischemic injury. Archives of Neurology, v.40, p.436-40, 1983. [ Links ]
WARRINGTON, E.K.; WEISKRANTZ, L. Amnesic syndrome: consolidation or retrieval? Nature, v.228, p.629-30, 1970. [ Links ]
WARRINGTON, E.K.; WEISKRANTZ, L. The effect of prior learning on subsequent retention in amnesic patients. Neuropsychologia, v.12, n.4, p.419-28, 1974. [ Links ]
WAUGH, N.C.; NORMAN, D.A. Primary memory. Psychological Review, v.72, n.2, p.89-104, 1965. [ Links ]
WEISKRANTZ, L. Neuroanatomy of memory and amnesia: a case for multiple memory systems. Human Neurobiology, v.6, n.2, p.93-105, 1987. [ Links ]
WEISKRANTZ,L.; WARRINGTON, E.K. Conditioning in amnesic patients. Neuropsychologia, v.17, p. 187-94, 1979. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R. Medial temporal lesions in monkeys impair memory on a variety of tasks sensitive to human amnesia. Behavioral Neuroscience, v.99,n.1,p.22-34, 1985. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R. Preserved learning in monkeys with medial temporal lesions: sparing of motor and cognitive skills. Journal of Neuroscience, v.4, p. 1072-85, 1984. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R.; AMARAL, D.G. Human amnesia and the medial temporal region: enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. Journal of Neuroscience, v.6, n.10, p.2950-67, 1986. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R.; AMARAL, D.G. Lesions of the amygdala that spare adjacent cortical regions do not impair memory or exarcebate the impairment following lesions of the hippocampal formation. Journal of Neuroscience, v.9, n.6, p. 1922-36, 1989. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R.; MISHKIN, M. The neuroanatomy of amnesia: amigdala-hippocampus versus temporal stem. Science, v.218, p. 1337-9, 1982. [ Links ]
1 William James, The principles of psychology, New York, Holt, 1890, v.1.
