










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista Psicologia e Saúde
versão On-line ISSN 2177-093X
Rev. Psicol. Saúde vol.8 no.1 Campo Grande jun. 2016
https://doi.org/10.20435/2177093X2016106
Um possível papel da neurofisiologia básica na explicação da mente pela psicologia
A possible role of basic neurophysiology in explanation of mind by psychology
Un posible papel de la neurofisiología básica en la explicación de la mente por la psicología
Flavio Kulevicz BartoszeckI; Yong Chiang ChangII; Amauri Betini BartoszeckIII
ISecretaria de Estado da Educação PR
IIUniversidade Federal do Paraná
IIIUniversidade Federal do Paraná
RESUMO
Este artigo tem como objetivo demonstrar a possibilidade de uma explicação material para a mente. Para tal, a Neurofisiologia pode dar exemplos em que a pesquisa básica pode perscrutar o fenômeno mental e defini-lo como material. As pesquisas básicas em Neurofisiologia podem ajudar as ciências humanas como a Psicologia a explicarem, de forma mais completa, aspetos da vida mental, sobretudo a vida mental humana. São analisados os circuitos do condicionamento associativo em Aplysia sp., aprendizagem e memória declarativa e não declarativa, condicionamento operante em vertebrados e contingência mecânica e intencional.
Palavras-chave: Neurofisiologia; Psicologia; Mente.
ABSTRACT
This paper aims to demonstrate a possible way for a material explanation of the Mind. In that matter, Neurophysiology can provide examples based in their basic research towards a materialistic approach of mental phenomena. The basics researches of Neurophysiology can help the human sciences such as Psychology in theirs explanation more fully aspects of mental life, mainly human mental lives. On Aplysia sp. are analyzed the circuits of associatives conditioned, learning and memory (declarative and not declarative), operant conditioned in vertebrates, mechanic and intentional contingency.
Key-words: Neurophysiology; Psychology; Mind.
RESUMEN
Este artículo tiene como objetivo demostrar la posibilidad de una explicación material para la mente. Para tal, la Neurofisiología puede dar ejemplos en que la pesquisa básica puede estudiar el fenómeno mental y definirlo como material. Las pesquisas básicas en neurofisiología pueden ayudar las ciencias humanas como la psicología a explicaren de forma más completa aspectos de la vida mental. Son analizados los circuitos del condicionamiento asociativo en Aplysia sp., aprendizaje y memoria declarativa y no declarativa, condicionamiento operante en vertebrados y contingencia mecánica y intencionado.
Palabras clave: Neurofisiología; Psicología; Mente.
Introdução
A tentativa de elucidar o comportamento humano (e animal) gerou várias abordagens de observação acurada e técnicas experimentais. Na linha das "ciencias duras", está a Fisiologia, que tem como objeto de estudo os constituintes estruturais e a atividade funcional dos seres vivos (Aires, 1985; Kandel et al., 1995; Fox, 2007). A mensuração da funcionalidade segue como sendo o atributo maior dessa área do conhecimento. Porém, ao chegar ao campo da explicação comportamental humana, a Fisiologia esbarra em barreiras, obstáculos esses que levaram muitos pesquisadores a afirmarem a dualidade da matéria, sendo o substrato mental diferente do substrato físico, resolvendo assim a problemática da explicação do mental (Turnbull & Solms, 2003).
A Psicologia, por outro lado, preocupa-se com as razões do comportamento humano e animal (Griffin, 1992; Matthews, 2007). Entenda-se aqui, razões pelo motivo de um animal reagir de certa forma ante a algum tipo de estímulo. Nesse ínterim, a Neurofisiologia poderia ser de real vantagem para a Psicologia em substanciar a busca pelas razões do comportamento, tanto animal quanto humano. Substanciar com medições, usando atributos físicos e químicos, para que não haja recaídas para o dualismo, ou seja, a afirmação o qual o mundo mental seria oriundo de partículas não "contabilizadas" pela ciência (Bartoszeck, 2006).
A Psicologia é uma ciência a qual estuda em princípio a atividade mental, particularmente a atividade humana (Gross, 1992). Contudo estudantes de graduação em Psicologia podem não corroborar com conceituação perpetrada pela comunidade científica demonstrando uma posição ambivalente sobre o status da Psicologia como ciência (Morales, Abranson, Nain, Aleixo Jr., & Bartoszeck, 2005; Bartoszeck, Abramson, & Place, 2005).
A Fisiologia tem, por objeto do estudo, a atividade corporal humana e animal (Ganong, 1993). A atividade corporal pode ser medida por instrumentos fisiológicos. Por exemplo, a pressão sanguínea de cão pode ser medida com o manômetro na aula prática de Fisiologia porque o sangue é conhecido como o substrato material da Fisiologia cardiovascular (Curi, Procopio, & Fernandes, 2005). Não há instrumento físico para medir a atividade psicológica humana, pois as técnicas empregadas com o ser humano carecem da resolução (sítio de medição muito amplo) e dado temporal (diferença entre o estímulo e resposta adequados). O único acesso à Psicologia de outro sujeito humano é através da conversação, ou seja, acesso indireto aos dados mentais (Wellman, 1990).
Portanto a identificação do substrato material que executa a atividade psicológica foi o objetivo e a esperança de filósofos, psicólogos e fisiologistas durante a longa história da civilização (Brazier, 1959; 1983; Churchland, 1986).
Contudo os psicólogos experimentais observaram, durante os anos iniciais do século XX, que algumas regras e leis fisiológicas poderiam também ser aplicadas à Psicologia. Por exemplo, o gato aprende abrir uma porta convertendo uma resposta eliciada por um estímulo e migrar para um novo estímulo que elicia uma nova resposta. A latência, ou período de tempo entre o estímulo e a resposta, diminui exponencialmente em cada ciclo dessa conversão, até o gato aprender abrir a porta. Essa diminuição exponencial das latências nos ciclos da aprendizagem-memória aplica-se em todos os comportamentos operantes sem exceção, em todos os animais, inclusive no ser humano (Thorndike, 1911). O cão normalmente saliva quando recebe um estímulo incondicionado, EI, à vista do alimento. Mas pode aprender a salivar ao estímulo condicionante, EC, como um som do sino, que normalmente não induz a resposta de salivação. Esse poder de aprender, memorizar e responder ao novo estímulo também está presente no comportamento clássico de todos os animais, incluindo o humano (Pavlov, 1927). A esperança de descobrir o substrato material da Psicologia foi prevista, "Devemos reconhecer que todas nossas ideias provisórias sobre a Psicologia serão consubstanciadas um dia baseadas nos substratos orgânicos" (Freud, 1914). Esse substrato foi sugerido como localizado na sinapse, "O sistema nervoso tem uma plasticidade de "fortalecer" a sinapse entre os neurônios quando um novo conhecimento é aprendido" (Hebb, 1949; LeDoux, 2002). Essas propostas, contudo, permaneceram como teorias abstratas porque as definições dos termos usados (e. g., fortalecer a sinapse) não puderam ser identificadas pela tecnologia da época dessas predições.
Registro de potencial com materialidade da mente: nasce a Neurofisiologia
Os métodos desenvolvidos durante os anos iniciais do vigésimo século para registrar (Ling & Gerald, 1949) e para visualizar (Grinvald, 1985) o potencial (sinônimo com a energia e a voltagem) elétrico de uma célula nervosa, ou neurônio, identificado (Stewart, 1981), estabeleceram uma possibilidade de identificar o potencial elétrico como o substrato material da atividade psicológica. A correlação definitiva entre o potencial elétrico e a atividade psicológica foi demonstrada na experiência feita por Kandel e seus colegas na década dos anos setenta, com o reflexo de retração da branquia da Aplysia sp, a lebre do mar (Kandel, Schwartz, Jessell, 1991). Semelhante como o reflexo da salivação do cão, a Aplysia sp retrai sua brânquia quando seu manto é estimulado por um estímulo tátil incondicionado, EI, porque um potencial pós-sináptico (primeira fileira, Fig. 1c) é acionado no moto-neurônio L7 (Fig. 1a) pelo EI no neurônio sensorial, SE, em seu manto (Fig. 1b). Um estímulo tátil condicionante, EC, na cabeça dessa Aplysia sp. não elicia a retração de sua brânquia antes da associação porque a sinapse entre o neurônio L28 da cabeça e o moto-neurônio L7 (Fig. 1b) não é fortalecida para eliciar um potencial pós-sináptico. Após determinada vezes da associação entre o EC e o EI (segunda fileira, Fig. 1c), um potencial pós-sináptico pode ser eliciado pelo EC sem EI (terceira fileira, Fig. 1c) porque a pré-sinapse entre o SE e o L7 já foi fortalecida pelo EC repetidos no L28. A Aplysia sp. já aprendeu e memorizou retrair sua brânquia pelo EC na cabeça, ao invés pelo EI no manto. Esse novo potencial pós-sináptico no L7 é o substrato material desse novo conhecimento. Os EC repetidos alteram a enzima NMDA na membrana pós-sináptica do L7. A NMDA alterada permite a entrada do íon Ca++ para formar um potencial pós-sináptico no L7. Essa experiência definiu o termo predito por Hebb (1949) "fortalecer a sinapse". A descoberta do potencial elétrico como o substrato material da atividade psicológica resultou em um Prêmio Nobel para Eric. R. Kandel, em 2000. Assim a Neurofisiologia se consolida como a busca dos substratos materiais com possibilidade de medição, principalmente o registro elétrico.
Aprendizado e memória declarativa e não-declarativas
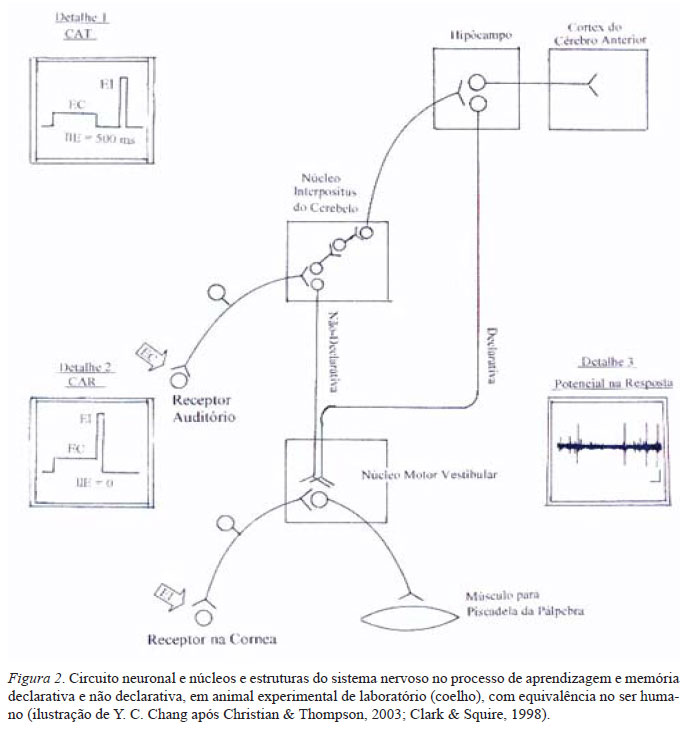
Após essa primeira experiência, muitas atividades psicológicas complexas foram explicadas analisando os potenciais elétricos que circulam nos sistemas nervosos animal e humano. Uma dessas experiências, feita com o reflexo de piscadela da pálpebra do coelho, demonstrou que a alteração dos potenciais elétricos é o substrato das duas atividades psicológicas da aprendizagem-memória, declarativa e não-declarativa. A aprendizagem-memória declarativa é quando aprendemos uma lição durante uma aula. Podemos declarar, ou descrever oralmente, para os outros o conteúdo aprendido e memorizado dessa lição. A aprendizagem-memória não-declarativa é quando aprendemos andar com a bicicleta. Apesar de retermos o item de memória nesse caso, não podemos declarar, ou descrever oralmente, para os outros como foi aprendido e o que foi memorizado.
Similar ao reflexo da retração da brânquia da Aplysia sp. e no reflexo da salivação do cão, o coelho pisca sua pálpebra somente quando sua córnea é estimulada, porque o estímulo tátil incondicionado, EI, no neurônio sensorial da córnea elicia uma resposta composta dos potenciais elétricos no moto-neurônio do núcleo vestibular (detalhe 3, Fig. 2) para contrair os músculos da pálpebra (parte inferior, Fig. 2). O estímulo condicionante, EC, sonoro no ouvido do coelho (centro, Fig. 2), não elicia esse reflexo da piscadela da pálpebra, porque esse EC precisa passar através de muitas sinapses "fracas" nos núcleos pontino e ce-rebelar, para alcançar os motoneurônios vestibulares (resumidas em Christian & Thompson, 2003). Essas sinapses fracas foram fortalecidas somente após a associação dos EC com o EI em muitas vezes, até o coelho aprender piscar sua pálpebra escutando o som, EC, ao invés de receber um EI tátil na córnea.
Esses EC e EI podem ser associados em dois paradigmas, o condicionamento associativo de traço, CAT, no qual o EC e o EI são separados por um intervalo inter estímulos, IIE, de 500 ms (detalhe 1, Fig. 2), e o condicionamento associativo retardado, CAR, no qual o EI coincide com o EI sem intervalo, ou IIE = 0 (detalhe 2, Fig. 2). É conhecido no ser humano (Clark & Squire, 1998) que o CAT induz a aprendizagem-memória declarativa (uma lição no exemplo), e o CAR induz a aprendizagem-memória não-declarativa (bicicleta no exemplo). Foi demonstrado que esses 500 ms de IIE possibilitou o fortalecimento das sinapses entre o núcleo interpositus do cerebelo e o hipocampo para consolidar a nova memória no córtex do cérebro anterior (direita, acima, Fig. 2) para ser declarada, quando no IIE = 0, somente as sinapses entre o núcleo interpositus do cerebelo e o núcleo vestibular são fortalecidos. A memória também é consolidada nesses núcleos, mas sem a declaração verbal pelo córtex do cérebro anterior. Foi comprovado que esses dois tipos de atividades psicológicas, declarativa e não-declarativa, estão presentes não somente no coelho e no humano, mas também em rato, e camundongo (Cotterill, 2001), e na Aplysia sp. (Brembs, Lorenzetti, Reyes, Baxter, & Byrne, 2002) porque as alterações semelhantes dos potenciais elétricos durante CAT e CAR são demonstradas nesses animais, apesar da diferença anatômica de seus sistemas nervosos.
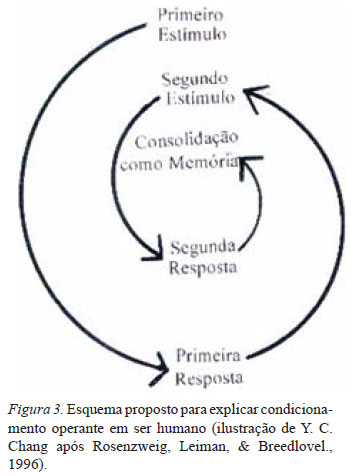
O estímulo é aplicado pelo experimentador no condicionamento clássico nos exemplos acima. Esse estímulo evoca uma resposta no sujeito. Em outro exemplo, o condicionamento operante, o estímulo é gerado pelo próprio sujeito. O primeiro estímulo gerado pelo sujeito evoca a primeira resposta no mesmo sujeito (Fig. 3). Essa primeira resposta torna-se o segundo estímulo para evocar a segunda resposta. Os estímulos e as respostas vão circulando no circuito neuronal da aprendizagem-memória pelo condicionamento operante até que a última resposta satisfatória é consolidada na memória desse sujeito. Portanto, para entender a diferença entre o condicionamento clássico e o condicionamento operante, é necessário entender a diferença entre o estímulo aplicado por outro sujeito e o estímulo aplicado pelo mesmo sujeito.
A contingência intencional e contingência mecânica
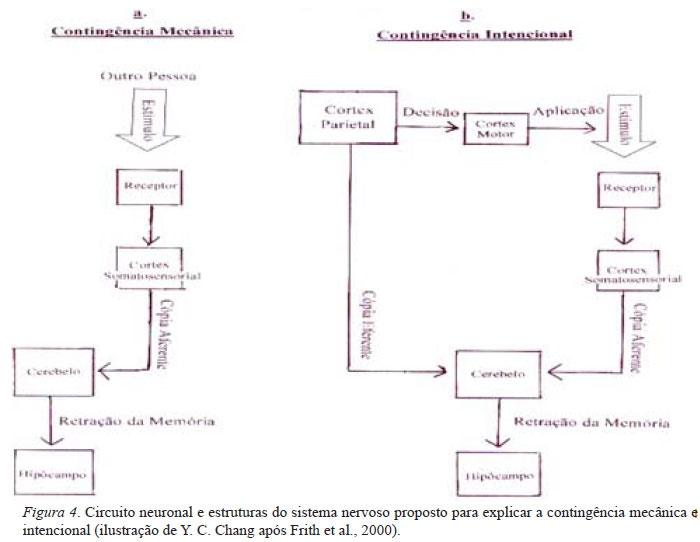
Um assunto tem intrigado os filósofos e cientistas desde Aristóteles: a impossibilidade de fazer cócegas em si mesmo (Bartoszeck, Kruzielski, & Chang, 2007). Ou seja, desde os tempos anteriores à Ciência, foi observado um comportamento curioso no ser humano: podemos fazer cócegas nos outros, porém, somos impossibilitados de realizar tal feito em nós mesmos. A neurofisiologia básica pode tentar elucidar tal "mistério", o qual intrigava os filósofos e pesquisadores utilizando as ferramentas de análise dos circuitos neuronais.
Testando a sensação psicológica de "cócega" induzida por um estímulo mecânico aplicado por outro sujeito e por mesmo sujeito, dois circuitos neuronais, a contingência mecânica e a contingência intencional, foram identificados pela neuro-imagem de ressonância magnética funcional, fMRI (Frith, Blakemore, & Wolpert, 2000). Esses dois circuitos (Fig. 4a e Fig. 4b) são semelhantes. Um sentido tátil é eliciado no córtex somato-sensorial por um estímulo mecânico (flecha, direta, cima, Fig. 4), como um contato do dedo, no receptor periférico em ambas as contingências. Esse sentido tátil é enviado como uma "cópia aferente" para ser analisado no cerebelo. Após análise, o cerebelo retrai uma memória prévia armazenada no hipocampo como uma sensação psicológica na contingência mecânica (Fig. 4a).
Quando um estímulo é originado no córtex parietal do próprio sujeito (Fig. 4b), o córtex parietal desse sujeito envia um comando eferente para o córtex motor para aplicar esse estímulo mecânico a fim de estimular um sentido tátil no seu próprio corpo e envia simultaneamente uma "cópia eferente" desse estímulo mecânico para seu cerebelo. O cerebelo compara o sentido tátil estimulado pelo estímulo virtual na "cópia eferente" com o sentido tátil desencadeado pelo estímulo real na "cópia aferente" e retém outra memória também armazenada no hipocampo, como se fosse uma sensação psicológica diferente da contingência intencional. Isto é um exemplo de duas sensações psicológicas diferentes, cócega e não-cócega, induzidas por um mesmo sentido tátil fisiológico dependendo do processamento através de dois circuitos neuronais diferentes no sujeito normal. Quando o anel da circuitaria da cópia eferente é interrompido pela esquizofrenia, o paciente pode sentir a cócega feita por si mesmo, porque esse paciente perdeu a contingência intencional (Blakemore, 2002). Essas duas contingências demonstram também o mecanismo da "animação" usado pelo organismo para distinguir o estímulo aplicado por outro organismo vivente e o estímulo aplicado por um objeto inanimado, por exemplo, um estimulador no laboratório (Blakemore, Boyer, Pachot-Chouard, Meltzoff, Segebarth, & Decety, 2003).
Considerações Finais
Este artigo tentou demonstrar a possibilidade de utilização dos estudos em Neurofisiologia Básica a favor de alicerçar-se as discussões tanto filosóficas quanto psicológicas. A Psicologia é tida como uma Ciência da Mente, porém, mesmo os estudantes de tal disciplina estão ainda indecisos com respeito ao papel explicativo da Psicologia quanto ao fenômeno mental.
O estudo da neurofisiologia básica é promissor no tocante a sistemas de organismos fechados e geralmente simples, utilizando modelos animais entre os invertebrados ou pequenos mamíferos. Assim, as técnicas de pesquisa em neurônios unitários ou em circuitos neuronais poderiam evidenciar uma materialidade da mente (Dethier & Stellar, 1970; Abramsom, 1994; Humphrey, 1994).
Estudos aparentemente sem relação como os realizados em minhocas e Aplysia sp, podem fornecer pistas e sanar alguns mistérios relatados por filósofos e psicólogos. São desconhecimentos, originados de nosso parco entendimento com respeito ao conteúdo mental. Este está relegado à esfera do desconhecido e inviolável, tanto por motivos religiosos e ideológicos, quanto técnicos (incapacidade de observar e registrar atividade em neurônios singulares, embora poli-registro, em humanos).
Portanto, com o entendimento de técnicas já consolidadas da Neurofisiologia (no uso do modelo animal) e o desenvolvimento de maiores entendimentos dos circuitos neuronais humanos, há possibilidade da Materialidade da Mente, demonstrando, em termos claros e seguros para estudantes de graduação, pós-graduação e pesquisadores, o caráter científico das disciplinas interessadas no fenômeno mental, como a Psicologia.
Referências
Abramson, C. I. (1994). A primer in invertebrate learning: The behavioral perspective. Washington, DC: American Psychological Association. [ Links ]
Aires, M. (Ed.). (1985). Fisiologia básica. Rio de Janeiro: Guanabara Koogan. [ Links ]
Bartoszeck, A. B., Abramson, C. I., & Place, A. J. (2005). Perception of students in South of Brazil of status of Psychology as Science. Psychology Reports, 97(3),750-756. [ Links ]
Bartoszeck, F. K. (2006, jan./jun.). Tipos de dualismos na Filosofia da mente. Revista Internacional de Filosofia Clínica, 3,82-96. [ Links ]
Bartoszeck, F. K., Kruzielski, L., Chang, Y. C. (2007, dez.). Intencionalidade da sensação uma conceituação funcionalista de identidade. Jornal de Ciências Cognitivas, Sociedade Portuguesa de Ciências Cognitivas. Disponível em http://jcienciascognitivas.home.sapo.pt/07-12_bartoszeck.html. [ Links ]
Blakemore, S. J. (2002). Schizophrenia and brain imaging. In H. D'Haenen, J. A. den Boer, & P. Wollner (Eds.). Biological Psychiatry (pp. 649-661). New York: J. Wiley & Sons. [ Links ]
Blakemore, S. J., Boyer, P., Pachot-Chouard, M., Meltzoff, A., Segebarth, C., & Decety, J. (2003). The detection of contingency and animacy from simple animation in the human brain. Cerebral Cortex, 13,837-844. [ Links ]
Brazier, M. A. B. (1959). The historical development of neurophysiology. In H. W. Magoun (Ed.), Handbook of Physiology (1st ed., pp. 1-74). Washington, DC: Am. Physiol. Soc. [ Links ]
Brazier, M. A. B. (1983). A history of neurophysiology in the 17th and 18th centuries. New York: Raven Press. [ Links ]
Brembs, B., Lorenzetti, F. D., Reyes, F. D., Baxter, D. A., & Byrne, J. H. (2002). Operant reward learning in Aplysia. Science, 296,1706-1709. [ Links ]
Christian, K. M., & Thompson, R. F. (2003). Neural substrates of eyeblink conditioning: Acquisition and retention. Learning and Memory, 10,427-455. [ Links ]
Churchland, P. S. (1986). Neurophylosophy: Toward a unified science of the Mind-Brain. Cambridge, Mass: MIT Press.
Clark, R. E., & Squire, L. R. (1998). Classical conditioning and brain systems: The role of awareness. Science, 280,77-81. [ Links ]
Coterill, R. M. J. (2001). Cooperation of the basal ganglia, cerebellum, sensory cerebrum and hippocampus: Possible implications for cognition, consciousness, inte lligence and creativity. Prog. Neurobiol., 64,1-33. [ Links ]
Curi, R., Procopio, J., & Fernandes, L. C. (Orgs.). (2005). Praticando Fisiologia. São Paulo: Manole. [ Links ]
Dethier, V. G., Stellar, E. (1970). Comportamento animal. São Paulo: Edgard Blucher. [ Links ]
Fox, S. I. (2007). Fisiologia humana. Barueri, SP: Manole.
Freud, S. (1914). On Narcissism. London: Hogarth Press. [ Links ]
Frith, C. D., Blakemore, S. J. & Wolpert, D. M. (2000). Abnormalities in the awareness and control of action. Phil Trans. Roy. Soc. London, Biol. Sci., 355,1771-1788. [ Links ]
Ganong. W. F. (1993). Review of Medical Physiology. East Norwalk, Connecticut: Appleton & Lang.
Griffin, D. R. (1992). Animals minds. Chicago, Ilinois: Univ. of Chicago Press.
Grinvald, A . (1985). Real-time optical mapping of neuronal activity. Ann. Rev. Neurosci., 8,263-305. [ Links ]
Gross, R. D. (1992). Psychology-the science of mind and behavior. Dunton Green, UK: Hodder & Stoughton.
Hebb, D. O. (1949). The organization of behavior. New York: J. Wiley & Sons. [ Links ]
Humphrey, H. (1994). Uma história da mente: A evolução e gênese da consciência. Rio de Janeiro: Campus. [ Links ]
Kandel, E. R., Schwartz, J. H., & Jessell, T. M. (1991). Principles of neural science. (3rd ed.). New York: Elsevier. [ Links ]
Kandel, E. R., Schwartz, J. H., & Jessell, T. H. (1995). Fundamentos da Neurociência e do Comportamento. Rio de Janeiro: Prentice-Hall do Brasil. [ Links ]
LeDoux, J. (2002). Synaptic self: how our brains become who we are. New York: Penguin Putman. [ Links ]
Ling, G. N., & Gerald, R. W. (1949). The normal membrane potential of frog sartorius fibers. J. Cell. Comp. Physiol., 34,383-396. [ Links ]
Matthews, E. (2007). Mente: Conceitos-chave em filosofia. Tradução Michelle Tse. Porto Alegre, RS: Artmed.
Morales, B. L., Abranson, C. I., Nain, S., Aleixo Jr., N, & Bartoszeck, A. B. (2005). Status of Psychology as a Science in Norttheast Brazil: undergraduate student's perceptions. Psychological Reports, 96(1),109-114. [ Links ]
Pavlov, I. P. (1927). Conditional reflexes: An investigation of the physiological activity of the cerebral cortex. Translated by G. N. Anrep. London: Oxford Press. [ Links ]
Rosenzweig, M. R., Leiman, A. L., Breedlove, S. M. (1996). Biological Psychology. Sunderland: Sinauer Associates. [ Links ]
Stewart, W. W. (1981). Lucifer dyes - Highly fluorescent dyes for biological tracing. Nature, 292,17-21. [ Links ]
Thorndike, E. L. (1911). Animal intelligence. New York: MacMillan. [ Links ]
Turnbull, O., & Solms, M. (2003). Mind and brain-how they relate. In The brain and the inner world: An introduction to the neuroscience of subjective experience. New York: Other Press Books. [ Links ]
Wellman, H. M. (1990). The child's theory of mind. Cambridge, MA: MIT Press.
 Endereço de Contato:
Endereço de Contato:
Flavio Kulevicz Bartoszeck
Faculdade Padre João Bagozzi, CEEBJA Paulo Freire
Curitiba, Paraná, Brasil. Cep 80230-060
Recebido: 02/10/2014
Última revisão: 28/03/2016
Aceite final: 12/04/2016
Sobre os autores:
Flavio Kulevicz Bartoszeck - Especialista em Neuropsicologia, Faculdade Padre João Bagozzi, CEEBJA Paulo Freire, Curitiba, Brasil. E-mail: flaviookb@gmail.com
Yong Chiang Chang - Doutor em Ciências Biológicas pela Universidade Federal do Paraná (UFPR). Laboratório de Neurofisiologia, Instituto de Saúde Dr. Bezerra de Menezes, Curitiba, Brasil.
Amauri Betini Bartoszeck - Fellow in Basic Medical Education, University of Washington, WA. Prof. Adjunto de Fisiologia, Laboratório de Neurociência & Educação Científica, Universidade Federal do Paraná, Brasil.


{kind=link}
{kind=link}