










Serviços Personalizados
Journal

artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.6 n.1 São Paulo jun. 2004
DOSSIER
Social play and spatial tolerance in tufted capuchin monkeys (Cebus apella)*
Brincadeira social e tolerância espacial em macacos-prego (Cebus apella)
Briseida Dôgo de Resende; Patrícia Izar; Eduardo B. Ottoni
Universidade de São Paulo
ABSTRACT
The aim of this work is to investigate the relations between social play, proximity and events of manipulative activities observed by conspecifics, in order to analise the dynamics involving tolerance and observation, important factors for an eventual social learning of tool use. The studied group lives in semifreedom at Tietê Ecological Park, SP, Brazil, and monkeys use stones to crack open encapsulated fruits. We did not find correlation between conspecific observation dyads and social play dyads because adults, who have limited participation at play, were the preferred conspecific observation targets in manipulative events, observed mainly by juveniles. There was a correlation between conspecific observation dyads and proximity dyads, indicating that social tolerance facilitates observation, but the correlation value was low, suggesting that monkeys, besides observing the individuals they stayed together with, also actively sought whom to observe.
Index terms: Play, Social learning, Primates, Cebus apella.
RESUMO
Este trabalho tem como objetivo investigar relações entre a brincadeira social, a proximidade e os eventos em que um macaco observa outro desempenhando atividades manipulativas, para analisar a dinâmica envolvendo tolerância e observação, fatores necessários para uma eventual aprendizagem social do uso de ferramentas. O grupo estudado vive em semiliberdade no Parque Ecológico do Tietê, SP, Brasil, e seus indivíduos utilizam pedras para acessar o conteúdo de frutos encapsulados. Não encontramos correlação entre as díades de observação e as díades de brincadeira, porque os adultos, que têm participação restrita na brincadeira, foram os principais alvos de observação co-específica em eventos manipulativos, sendo observados principalmente pelos jovens. Encontramos uma correlação entre as díades de observação e as díades de proximidade, indicando que tolerância social propicia a observação, mas o valor desta correlação foi baixo, sugerindo que os macacos, além de observarem aqueles com quem andavam juntos, também procuraram ativamente a quem observar.
Descritores: Brincadeira, Aprendizagem social, Primatas, Cebus apella.
Tufted capuchin monkeys (Cebus apella) are broadly distributed throughout South America. In natural habitats, group size varies from solitary to more than 30 animals (Lynch & Rímoli, 2000), with multi-male/multi-female structure (Terborgh, 1983). They stand out for their great manipulative capacity, what allows the non-stereotyped use of tools, comparable to hominoids. The possibilities for occurrence of social learning depend critically on the level of spatial tolerance among individuals, which determines the level of detail with which an "observer" can watch the manipulatory activity of a proficient conspecific (Coussi-Korbel & Fragaszy, 1995). The study of social relations can help understand the dynamics involving tolerance and behavior observation, precondition for eventual processes of social learning.
Among these social relations, social play attracts attention because it is a relevant feature of immature primates, which are frequent learners (Walters, 1987). The concept of "play" is difficult to define (Beckoff & Byers, 1998), but it is suggested that it can function as a training for adaptatively relevant behaviors and can also play a role in the construction of social structures (Fagen, 1981; Walters, 1987). It is a non-agonistic behavior that incorporates many physical components of the behavioral pattern of adults, like those used during aggression (without their immediate consequences), being exaggerated, repetitive and varied.
The aim of this work is to investigate the relations between social play, proximity and the events in which a monkey observes a conspecific performing manipulative activities, especially tool-aided nutcracking. Our hypothesis was that social play, facilitating the construction of social bonds, could also help to increase spatial tolerance (as suggested in Resende & Ottoni, 2002) towards conspecific observers, an important factor for the occurrence of social learning.
Method
Subjects and Study Area
The studied group lives in an area of 180,000m2 at Tietê Ecological Park, São Paulo, Brazil, where, besides being provisioned, they forage on naturally available food items (fruits, leaves, small prey). Among a variety of fruit trees, the palm tree Syagrus romanzoffiana is important because of its nuts, which are cracked by the monkeys with the aid of stones (Ottoni & Mannu, 2001).
From March/2000 to March/2002, group size varied from 17 to 23 individuals, due to dispersion, births and deaths. By the time data collection ended, there were 11 adults or sub-adults (six males, five females), four juveniles (three males, one female) and an infant female.
The use of terrestrial space was intense, possibly due to the absence of predators and to the existence of large open areas. Most of the tool use episodes occurred on the ground.
Procedure
Data were collected from March/2000 to July/2002. Social Play was registered through Animal Focal Sampling (Altmann, 1974): each subject was followed for 10 minutes, from two to eight times a week (subjects under three years-old) or twice a month (subjects over over three years-old). Social Play was operationally defined as a non-agonistic interaction between subjects involving running, chasing, biting, poking, rolling or jumping towards the partner ("play fighting" and "rough-and-tumble"). Play was performed on the ground, in the bushes, in the trees or on buildings like green-houses or roofs. We registered the time spent playing and the identity of Social Play partners. These data were transformed into an n x n matrix (n = individuals) that was used to calculate the Jaccard's Similarity Index (J):
J(x, y) = a /a + b + c
where a = play time between x and y; b = time x social played with subjects other than y; c = time y social played with subjects other than x.
In order to study Proximity relationships, we registered (through Focal Scan sampling, after each Focal Animal sampling) all the individuals who were within 10m or less from the focal subject. At an n X p matrix (individuals X instants), "1" was assigned to these subjects and those who were not present were assigned "0". This matrix was converted to an n X n matrix with Jaccard's Similarity Index (J) (where a = frequency in which x and y were scored near; b = frequency of scores in which x was registered, but y was not; c = frequency of scores in which y was registered, but x was not).
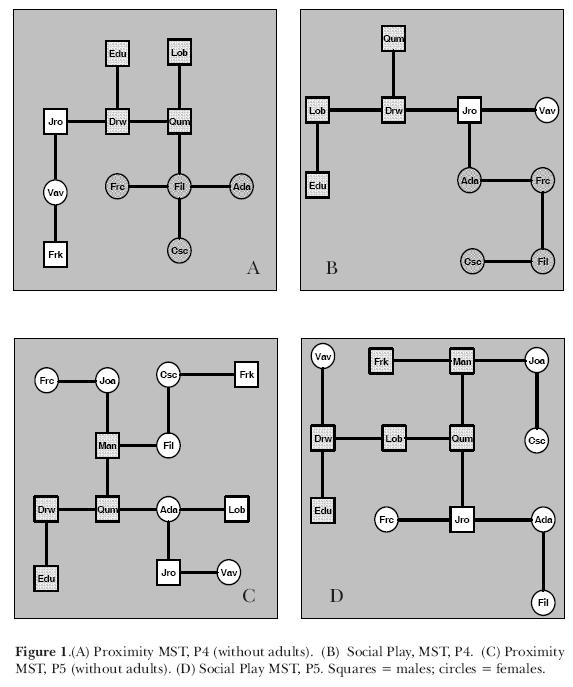
The similarity matrices of Proximity and Social Play were transformed into dissimilarity matrices using the complementary values of Jaccard's indexes (1-J) (Izar & Sato, 1997). Minimum Spanning Trees (MST) were built from the dissimilarity matrices of Social Play and Proximity, what allows a graphical visualization of the strongest relations between group members. As adults and subadults did not play much, they were excluded from both trees and we compared the social structures found in the Proximity and Social Play MSTs. Male juveniles tend to form subgroups for Proximity and Social Play (Izar, 1994). To check the occurrence of this phenomenon here, the Organization Index (OI) was calculated for male and female juvenile subgroups. This index, which varies from zero to one, is a measure of the level of association between individuals in a given subgroup. The larger the organization of a subgroup at the studied tree, the closer to one that value will be.
OI = S ni Log ni/n Log n,
where n is the number of subjects found at the male (or female) juvenile subgroup of Social Play MSTs and ni is the number of juvenile males (or females) from subgroups found at the Proximity MSTs. Proximity and Social Play data refer to the same period, that is, the same play relations are included at the Proximity and Social Play MSTs. However, besides play behavior, the Proximity MST also includes other activities. Thus, if play is an important activity in the Proximity relations of a subject, the liaisons observed at the Social Play MST will be reflected at the Proximity MST. If the subject spent too long performing other social activities (such as grooming, resting or eating together) and few time playing, its liaisons can be different at the Proximity and Play MSTs. The OIs calculated in this work show how the Proximity relations reflect Social Play relations.
Conspecific Observation was registered when the subject turned its face towards other monkey, persisting at this posture and at a distance inferior to 1m. The activity of the monkeys who were being observed by the conspecific was also registered and consisted mainly of foraging behavior, such as food manipulation and nutcracking. These data were collected through Focal Animal Sampling (as previously described for Social Play) and Ad libitum Sampling. There is a bias towards Nutcracking Observation, since Ad libitum Sampling refers only to nutcracking episodes. We built an n X n asymmetric matrix from the Conspecfic Observation frequency, in which columns represented the observation "targets" and lines represented the "observers", and a symmetric n X n matrix , in which cells represented the frequency of association between each dyad during observation events. Within this data set, we applied the K Test (Hemelrijk, 1990) to correlate the matrices of Conspecific Observation with the matrices of Proximity and of Social Play, where we considered only those individuals who played at least at 5% of the total Social Play time.
From Proximity and Social Play MSTs, we calculated, for each subject, two Vertice Degrees (VD = number of subjects to whom they are linked): Social Play VD and Proximity VD. This measure reflects the subjects' "Popularity", as those who have stronger relations with a higher number of monkeys are linked to more subjects, resulting in a higher VD. In order to check if the Popularity was related to Conspecific Observation, that is, to check if the most observed animals were also the most popular ones, the frequency of Conspecific Observation was correlated to Social Play and Proximity's VD using Spearman Test .
A Kruskall-Wallis Test was conducted to check if there were differences in the time dedicated to Social Play by Adults/Subadults, Juveniles and Infants. If significance was detected, Mann-Whiney tests were applied to find which age classes were responsible for the significance. Kruskall-Wallis and Mann-Whitney tests were also applied to check if Conspecific Observations differed according to age.
Results and Discussion
Social Play represented around 4% of the studied monkeys' time (total Social Play time/ total focal sampling time). However, considering only adults and subadults, we noticed that they dedicated 0.08% to Social Play, while immatures (juveniles + infants) did that in 4.8% of the time. There is a significant difference between time dedicated to Social Play by adults/ subadults, juveniles and infants [Kruskal-Wallis: ?2 (2, n = 22) = 9403, p = 0.009], and adults played less than infants and juveniles (Mann-Whitney : adults and juveniles, z = 3.06, p = 0.003; infants and adults, z = -2215, p = 0.027. Medians: infants, m = 2795.9; juveniles, m = 3293.4; adults, m = 230,03). Considering Conspecific Observation in manipulative events, monkeys from different age classes had different chances of being "targets" [Kruskal-Wallis: ?2 (2, n = 20) = 7239, p = 0.027]. Adults were significantly more frequent "targets" than juveniles and infants (Mann-Whitney : infants and adults, z = -2484, p = 0.013; infants and juveniles, z = -2466, p = 0.014; juveniles and adults = non-significant. Medians: infants, m = 1; juveniles, m = 12; adults, m = 15). Considering only the nutcracking episodes, there was no difference between episodes in which juveniles and adults were observed, in spite of the fact that juveniles had a higher cracking rate (total number of nutcracking episodes from all member of the age category/months of data collection: adults = 0.82; juveniles = 1.60).
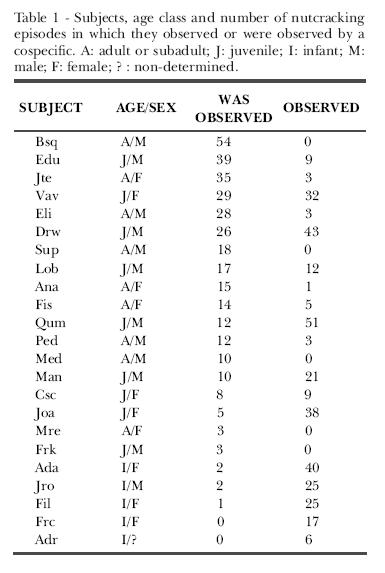
The distribution of the frequency of Conspecific Observation for adults or subadults, juveniles and infants differed from what would be expected by chance [Kruskal-Wallis: ?2 (2, n = 20) =11677, p = 0.003]. Juveniles and infants observed their conspecifics significantly more than adults (Mann-Whitney: adults and infants, z =-1985, p =0.047; adults and juveniles, z = -3217, p = 0.001. Medians: infants, m = 25; juveniles, m = 21; adults, m = 1). No adult observed more than 5% of the episodes, but four were observed in more than 5% of the episodes: the dominant male (15.75%), an adult female (10.20%) and two other adult males (8.16 % e 5.25 %). The other adult females were observed only by their offspring. Table 1 shows the subjects, their age/sex and the number of episodes they observed or were observed by a conspecific.
Most Conspecific Observation events involved observers at least one year younger than the "targets" (83%). Only a small part involved observers more than one year older than the "targets" (6.4%). When we divided these numbers by the total number of possible dyads for each category (same age/ observers at least one year older/ observers least one year younger), dyads with observers at least one year younger appeared in an adjusted frequency 2.35 times higher than same age dyads, an extremely significant difference (Goodnes-of-Fit: X2 = 63966; d.f.. = 2; p < 0.0001). From the 22 events in which a (at least one year) younger subject was observed by an older individual, juvenile Darwin was the "target" in 12. Juveniles with higher nutcracking rates (Edu and Vavá) were not so frequently observed. Darwin had the highest Proximity and Social Play VDs among juveniles. However, when we considered the "targets" from all the Conspecific Observation events and the number of liaisons in the MSTs, no correlation between Popularity and frequency of Conspecific Observation was found. Despite having the highest Social Play VD and the second highest Proximity VD, juvenile Darwin was only the sixth most observed monkey (8% of the events).
Within the set of dyads of "observers" and "targets", considering the asymmetric matrix, there was no correlation between Conspecific Observation and Proximity (Tau Kr = 0.132, one tailed right-sided p = 0.1334, n = 20, permutations = 2000) and between Conspecific Observation and Social Play (Tau Kr = 0.012, one tailed right-sided p = 0.4468, n = 13, permutations = 2000). Considering the symmetric matrix, there was a significant, but low, correlation between Proximity and Conspecfic Observation (Tau Kr = 0,212, one tailed right-sided p = 0.006, n = 20, permutations = 2000). The same result was found for the correlation between Conspecific Observation and Social Play (Tau Kr = 0,143, one tailed right-sided p = 0.0485, n = 13, permutations = 2000).
Social Play dyads formed by same-aged monkeys and different-aged monkeys (difference of 12 months or more) appeared in a 1:1 proportion. Considering the frequency of Social Play, we normalized the number of registered dyads by possible number of dyads, and no preference for partner age was found (X2 = 0.208, d.f. = 1, p = 0.6483). However, considering the average Social Play time of the dyads within same-aged and different-aged monkeys, we noticed that they spent more time playing with same-aged monkeys (X2 = 2742636, d.f.=1, p < 0.0001).
In order to analyze the MSTs of Proximity and Social Play, we divided the data in six periods (P1 to P6), according to births and deaths that occurred during data collection. Figure 1 show the Proximity and Social Play MSTs from periods 4 e 5, as an example of the structures observed at the MSTs. Comparing the Organization Indeces, we found exactly the same subgroup of juvenile males at Proximity and Social Play MSTs in three periods (OI = 1 for periods 3, 5 and 6), that is, Proximity MSTs reflected Social Play MSTs. OIs in other periods were: P1 = 0.69, P2 = 0.25 and P4 = 0.51. Juvenile males who were not linked to the age subgroup at the Proximity MST were peripheral, almost dispersing (Frank) or had other strongest Proximity relations, as a newborn sibling (as juvenile Lobato and his sister Ada). During the second period, OI was low because female juvenile Joana was separating the group of juvenile males, once her strongest relations were with three juvenile males. Among the females, there are no juvenile subgroup formation at the Proximity and Social Play MST, except for the fourth period, where there was a dyad. Considering all immature females (infants and juveniles), there were subgroups at the Social Play MSTs from periods 5 and 6. These subgroups were also seen in the Proximity MSTs built without adults, showing the link between juvenile females and infants, which maybe related to a special type of allomaternal care, also called "play mothering" (Smith, 1978). At the Proximity MSTs including adults, infant females are linked to their mothers.
We hypothesized that Social Play could promote spatial tolerance, thus increasing opportunities to social learning. However, we found that Social Play is a typical interaction of immature monkeys, promoting spatial proximity only among individuals in this age class, specially juvenile males. Moreover, although monkeys preferred playing with same-age partners, they preferred observing older "targets" during manipulative events.
When we considered the direction of the interaction, we also did not find the hypothesized correlation between Proximity and Conspecific Observation. This result is probably related to the influence of age on Observation, because while immatures were the main observers, adults were primarily observed. When we considered Conspecific Observation as a symmetric interaction, thus eliminating the effect of age on observation, we found that correlation. Hence, monkeys who stayed together tended to be also together at the Conspecific Observation events. The tolerance exhibited while they stayed together was extended to the events of Conspecific Observation. A potential learner near a nutcracking event and tolerated by the "cracker" had the opportunity of eating nuts from the nutcracking site and of manipulating the available stones and nuts, what could work as a reinforcer for nutcracking sites' exploration. Thus, spatial tolerance could make learning by "stimulus-" and "local enhancement" easier.
Nevertheless, this correlation between Conspecific Observation and Proximity (in the symmetric matrix) was low, indicating that Conspecific Observation events may be more than just a natural consequence of being together. It is possible that the monkeys, specially immatures, actively seek to observe some individuals' manipulatory behavior; it raises the question of what features make an individual more likely than others to become a target of observation while manipulating objects. The fact that Conspecific Observation does not seem to be an entirely passive consequence of previous proximity to the observed nutcracker points out to the possibility that features related to the proficiency of the "target" in nutcracking might influence the observers' interest and target choices. Further research will be needed to clarify these questions.
References
Altmann, J. (1974). Observational study of behaviour: sampling methods. Behavior, 49, 223-265. [ Links ]
Beckoff, M. & Byers, J. (Orgs.) (1998). Animal Play: Evolutionary, Comparative, and Ecological Perspectives. Cambridge: Cambridge University Press. [ Links ]
Coussi-Korbel, S. & Fragaszy, D. M. (1995). On the relation between social dynamics and social learning. Animal Behaviour, 50, 1441-1553. [ Links ]
Fagen, R. (1981). Animal Play Behavior. New York: Oxford University Press [ Links ]
Izar, P. (1994). Análise da Estrutura Social de um grupo de macacos-prego (Cebus apella) em condições de semi-cativeiro. Dissertação de Mestrado, Instituto de Psicologia, Universidade de São Paulo, SP. [ Links ]
Izar, P. & Sato, T. (1997). Influência de abundância alimentar sobre a estrutura de espaçamento interindividual e relações de dominância em um grupo de macacos-prego (Cebus apella). Em S. Ferrari & H.Schneider (Eds), A Primatologia no Brasil 5. Belém, PA. [ Links ]
Lynch & Rímoli (2000) Demography and social structure of group of Cebus apella nigritus (Goldfuss, 1809, Primates/Cebidae) at Estação Biológica de Caratinga, Minas Gerais. Neotropical Primates, 8, 44 - 49. [ Links ]
Ottoni, E. B. & Mannu, M. (2001). Semifree-ranging tufted capuchins (Cebus apella) spontaneously use tools to crack open nuts. International Journal of Primatology, 22, 347-358. [ Links ]
Resende, B.D. & Ottoni, E. B. (2002). Brincadeira e Aprendizagem do uso de ferramentas em macacos-prego (Cebus apella). Estudos de Psicologia (Natal), 7, 173-180. [ Links ]
Smith, E.O. (1978). Social Play in Primates. Academic Press, New York. [ Links ]
Terborgh (1983). Five new world primates. Princeton University Press, Princeton. [ Links ]
Walters, J. (1987). Transition to adulthood. Em B.B. Smuts; R. M. Seyfarth; R. M. Wrangham; T.T. Struhsaker (Eds.), Primate Societies (pp.358-369). Chicago: University of Chicago Press. [ Links ]
 Correspondence
Correspondence
Briseida Dôgo de Resende
Av. Diógenes Ribeiro de Lima, 2000, Bloco 11, apto 11, Alto de Pinheiros
05458 001, São Paulo, SP, Brasil
E-mail: briseida@usp.br
Received September 03, 2003
Revision received November 17, 2003
Accepted March, 2004
* Paper presented at the III Congresso Norte-Nordeste de Psicologia, João Pessoa, Brazil, 2003. We thank the staff of Tietê Ecological Park, where we conducted our study and Carlos Eduardo Guidorizzi de Carvalho, who helped in data collection.
Financial support: FAPESP, 99/11573-2.

