










Serviços Personalizados
Journal

artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.7 n.2 São Paulo dez. 2005
ARTIGOS
The prey capture behaviour of Metepeira gressa (Araneae, Araneidae) on mealworms, flies and ants
Captura de presas de Metepeira gressa (Araneae, Araneidae) con larvas de tenébrio, moscas y hormigas
Carmen Vieira*
Universidad de la República, Montevideo, Uruguay
RESUMO
The prey-capture behavioral repertoire of females and juveniles of the araneid spider Metepeira gressa (Keyserling, 1892) was analyzed using mealworm larvae, flies, and ants as prey of the same size. Flies and ants are frequently encountered by the spider species, but the beetle larvae are not. During the experimental period each type of prey was offered only once to each spider. The first unit of immobilization is very important to subdue the prey. When capturing mealworms and ants, spiders performed Wrap as the first unit of the Immobilization phase, and while capturing flies the spiders used Bites. Wrap seems to be a discriminatory initial behavior on dangerous prey, such as ants. The discrimination between ants and mealworms was tested using the observed behavioral units in spite of both generated similar capture patterns. The only difference detected was the occurrence of the unit Other Locomotion, more frequently on mealworm. The spider has many difficulties in mealworm capture because of the large size of the prey and the strength and the peculiar defensive tactic (wriggling) that progressively destroys the spider web. In disagree with the expectations, the spiders spent more time and energy in the capture of mealworms.
Index terms: Prey-capture, Spider, Metepeira gressa.
ABSTRACT
The prey-capture behavioral repertoire of females and juveniles of the araneid spider Metepeira gressa (Keyserling, 1892) was analyzed using mealworm larvae, flies, and ants as prey of the same size. Flies and ants are frequently encountered by the spider species, but the beetle larvae are not. During the experimental period each type of prey was offered only once to each spider. The first unit of immobilization is very important to subdue the prey. When capturing mealworms and ants, spiders performed Wrap as the first unit of the Immobilization phase, and while capturing flies the spiders used Bites. Wrap seems to be a discriminatory initial behavior on dangerous prey, such as ants. The discrimination between ants and mealworms was tested using the observed behavioral units in spite of both generated similar capture patterns. The only difference detected was the occurrence of the unit Other Locomotion, more frequently on mealworm. The spider has many difficulties in mealworm capture because of the large size of the prey and the strength and the peculiar defensive tactic (wriggling) that progressively destroys the spider web. In disagree with the expectations, the spiders spent more time and energy in the capture of mealworms.
Index terms: Comportamiento de captura, Arañas, Metepeira gressa.
Many orb spiders are polyphagous and, in spite of this, several studies have shown that many spiders show different behavior towards different prey (Nentwig, 1987; Riechert & Luczak, 1982; Stowe, 1986). In his classic works Peters (1954, 1955) showed that prey capture consists of sequences that vary in the order and composition of displayed behaviors according to the prey type. He observed that the attack behaviors of Araneus diadematus (Clerck, 1757) on a dead fly were different from those on a living fly. The first quantitative analysis of behavioral sequences and discrimination of Argiope argentata (Fabricius, 1775) was made by Robinson & Olazarri (1971). Robinson & Robinson (1976) detected the spider´s discrimination of lepidoptera adults from other prey types in at least twelve araneid species. Viera (1981, 1983) described the tactic of Alpaida alticeps (Keyserling, 1865) over three kind of prey. Suter (1978) found that the vibrations induced on the web by each of the prey species were evaluated by Cyclosa turbinata and the spider could discriminate among prey items.Burgess and Witt (1976) analyzed the web design in Metepeira spinipes(Cambridge, 1903) and Uetz et al. (1980, 1982) described the prey capture. Metepeira labyrinthea (Hentz, 1847) was studied by Burgess & Uetz (1982) and the competition for prey was studied by Wise (1983). Uetz (1990) also studied qualitative characters of the prey capture in these species.
Metepeira gressa (Keyserling) is a solitary small spider (5 to 10 mm of cephalothorax length). She builds its webs in plants of Eryngiumsp. and its retreat is cryptic with Eryngium dry fruits. The web of M. gressa is an asymmetrical orbweb with a retreat and barrier web, slightly inclined with respect to the vertical plane (Viera, 1983, 1989). Viera (1994, 1995) also analyzed the prey capture and found discrimination in the predatory behavior of M. gressa elicited by prey that share the spiders habitat, such as ants and flies.
The main questions in this study were: how variable or plastic is predatory behavior, and more specifically, how does Metepeira gressarespond to prey it has never experienced? To address these questions, I first studied how M. gressa captures prey common in its environment and then examined its capture of mealworm larvae, Tenebrio Linnaeus, a putatively rare prey type.
Methods
Fifteen adult females and juvenile spiders (last stages) collected in April 1997 in Punta Espinillo (Montevideo) were used in these experiments. Adult males were not included because they do not construct orb webs (Viera & Costa, 1985, 1988). Voucher specimens were deposited in both the Museum of Comparative Zoology (Harvard University, USA; identified by H. W. Levi and W. Piel) and the Colección Entomológica de la Facultad de Ciencias, Montevideo, Uruguay (collection number 957).
Spider Housing
The spiders were maintained for three months under approximately constant laboratory conditions; mean temperature was 22.5ºC (± S.D. 2.67 º C) and the photoperiod was 12 h light/12 h dark. The spiders were housed in individual glass cylindrical jars of 14 cm height and 9 cm diameter covered with nylon mesh, with a dish with water and a frame to hold the spider´s web. Between experiments the spiders were fed with parts of dead insects, preventing the influence of experience with alive prey on subsequent experimentation.
Experimental Procedures
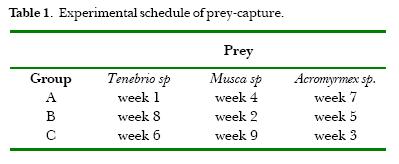
The spiders were transferred five days before each experiment to glass boxes of 30 cm height, 30 cm width and 10 cm depth, with a removable wall and a wooden frame to hold the web. Three groups (A, B and C), each composed of five spiders, received 3 different prey items of the same size as show in Table 1. The mentioned order of prey capture was made to prevent the possible influence of previous capture. Each prey type was offered once to each spider.
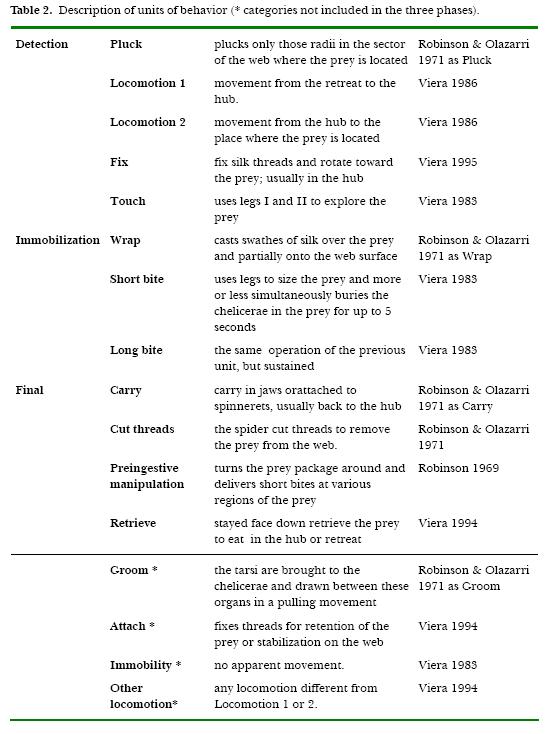
For observations, one wall of the glass box was removed, the back wall was covered with black paper and a lateral lamp of 445 lux was used to iluminate the web. Behavioral observations were made through a magnifying glass (2 X ,14.4 cm diameter). The capture sequence was divided into three behavioral phases (Viera, 1995): Detection, Immobilization and Final phase. Within these three phases, 16 units of behavior were observed. (Table 2).
Transition matrices were used to show the sequences and frequencies of behavioral units. Student´s T test with restrictions of the variance (Snedecor´s F) and discriminant analysis (Legendre & Legendre, 1984) were used to compare experimental groups. Discriminant analysis combines procedures from the analysis of variance and principal component analysis. One of its objectives is to discriminate as better as possible between g groups taking into account p variables. Other objective is to classify the observation. If the group are well separated, the discrimination is good, an viceversa. The correlation circle and the discriminant factorial axes is a graphic representation analog to those in PCA.
Results
Fly Capture
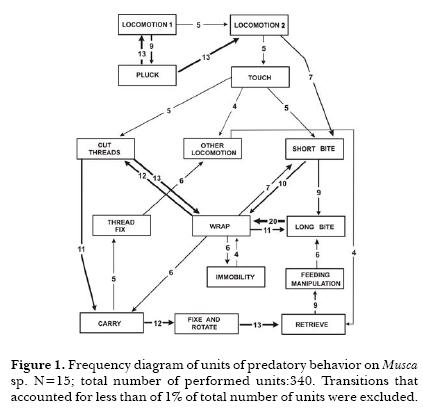
Detection phase started with Pluck (13 from 15 observations). Most of spiders started Immobilization with Short Bites. The association between Wrap and Short Bites was minor than the association between Wrap and Long Bite. Cut threads was strongly related with Wrap. The unit Immobility was frequent (23 times). Groom had a high occurrence, associated to the Immobilization phase. (Fig.1)
Ant Capture
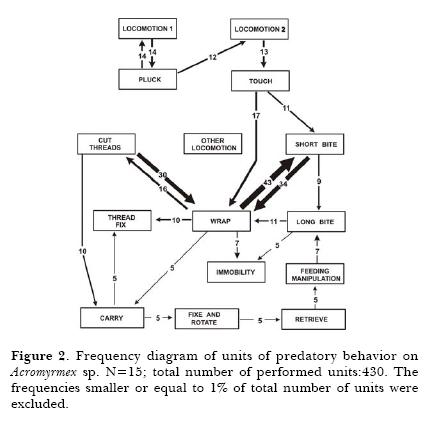
Detection phase started with Pluck (14 from 15 observations). All spiders started the Immobilization Phase with Wrap followed by Short Bites; these two units had a weak association with Long Bite. The relation between Immobilization and Final phases was through the succession Wrap- Cut threads. Groom had a high occurrence (9 from 15 prey-capture sequences). The unit Immobility was the most frequent (27 times). (Fig.2).
Mealworm Capture
All the individuals made Pluck (in retreat) as a first unit of behavior, followed by Locomotion 1 (from retreat to the hub). Thirteen spiders made Pluck in the hub and Locomotion 2 to arrive at the prey location. Ten spiders made Locomotion after Pluck. After arrived to the prey, 13 spiders performed Touch and 2 directly made Wrap. These units composed the Detection Phase.
The Phase of Immobilization started with Wrap in 11 cases, with Short Bite in three cases and with Long Bite in one case. A strong association between Wrap and Short Bite (Fig.3) was observed in all cases; the association between Short Bite and Long Bite was weak. The unit Other locomotion was related with Wrap, Short Bite and Long Bite with a high frequency (n=85). The unit Cut threads was closely related to Wrap, being the first unit a nexus between Immobility and Final phases. The Final phase showed high stereotypy. Eleven specimens made Fix and Rotate and 4 made Attach after Carry. Seven individuals made Retrieve and Long Bite after Fix and Rotate. Three individuals made Groom after Fix and Rotate, 2 individuals made Immobility and Touch twice.
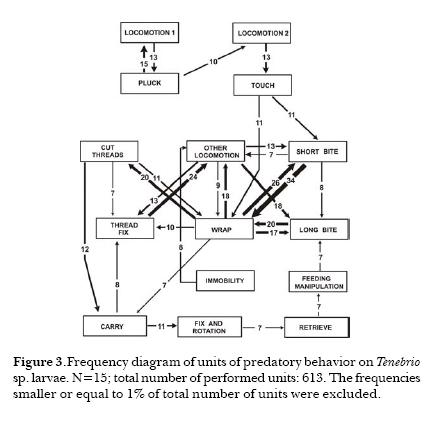
Groom occurred 8 times and is mostly related with Immobility and Retrieve; few interactions with Touch, Wrap and Long Bite occurred.
The unit Immobility had a high occurrence being followed by Other Locomotion, Wrap, Pluck, Retrieve, Touch, Carry, Cut Thread and Manipulation.
Comparison Among Captures
There were significant differences in total capture duration, between fly and mealworm and between fly and ant (Table 3). Also the table shows significant differences in the duration of immobilization of fly related to both mealworm and ant. Because of the similarity observed in the capture duration between mealworm and ant, I compared the averages of the occurrences of 7 units of behavior (Table 4). Only one significant difference in the unit Other locomotion was found. Discriminant analysis among the three types of prey-capture was done (Fig. 4). In this analysis of frequency, 8 variables were used: occurrence of Other Locomotion, occurrence of units of the Detection Phase, occurrence of units of Immobility Phase, occurrence of total number of units, total duration of the capture, duration of Detection, duration of Immobility, and duration of the Final Phase. The analysis showed a correlation between the variables and for each variable of the three experimental prey groups (fly, mealworm and ant). Significant F values were showed for Other Locomotion (0.27%), in the total number of behavioral units and with a lower significance in the frequency of units in the Immobility Phase, suggesting the importance of this variable for the discrimination of the prey. The diagram of sequences showed that the unit Other Locomotion was strongly related with Thread Cut, Wrap and mainly with Fixing of Threads.
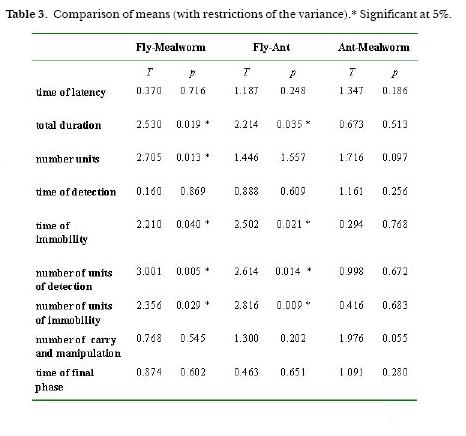
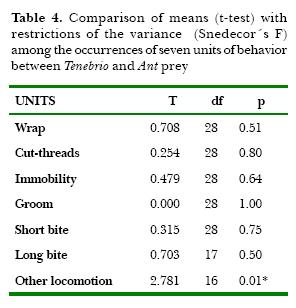
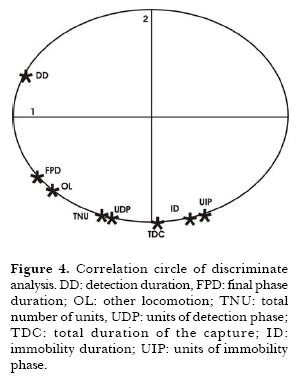
Discussion
The similarity of predatory behavior in adult females and large juveniles of Metepeira gressa described by Viera (1983) and Viera and Costa (1985, 1988) allowed me to use both groups together as a homogeneous group. Robinson & Robinson (1976) demonstrated the influence of learning in the second encounter of the spiders with the same type of prey. In the present paper the absence of differences among captures on mealworm according the different order of given prey, allow me to discard influences of immediately previous captures on other prey.
Robinson and Mirick (1971), Japyassú and Ades (1990), Ades, Vieira and Japyassú (1990) showed that the size of the prey affects the tactic used during prey capture. Japyassú and Viera (2002) found that the spider varies its behavior according to prey size and kind. Taking into account prey size, I avoided the influence of the size using prey of similar size to the spider. Avoidance behaviors of flies obviously are not dangerous for the spider; they remain immobile during a brief time, as observed by Hays (1985). After the spider plucks, the fly moves and shakes strongly. Pluck can be useful for the spider, because allows it to locate immobile objects. Also, the level of fly stickiness is minor too (Eisner & Dean, 1976). The spider´s high response speed and the small number of units involved in the Detection phase allow a fast capture of flies, suggesting an adaptive tactic of the spider.
Ants and mealworms generated similar capture patterns. The only difference detected was the occurrence of the unit Other Locomotion, more frequently on mealworm. This unit appeared related to the Immobilization Phase and mainly to the web-repairing process. The wriggling behavior of mealworm in the web usually broke several sticky threads and some radii, eliciting an increase in the frequency of Other Locomotion and a higher number of units (but not statistically significant) to immobilize the prey. Based in my observations this spider did not show expulsed tactics like other spiders as a Nephilengys cruentata (Japyassú & Viera, 2002). Prey capture and discriminatory tactics are limited and seems to follow basically a pattern, in agreement with Robinson & Robinson (1976).
Mielle (1978) showed in Tegenaria parietina (Fourcroy) (Agelenidae) that juveniles raised with one type of living prey, can discriminate other prey. That discrimination is instinctive and Ibarra-Núñez (1984) experimentally proved that young spiders of Tegenaria sp. without previous experience had the ability to discriminate among different preys. Using flies and ants, Viera (1995) agreed with this statement for Metepeira gressa(Araneidae).
In Tenebrio captures another factor, such as defensive tactics, is added to the unknown prey factor. This defensive procedure is not dangerous for the spider, but the mealworm breaks the web and consequently the spider performs a more complex behavior. This fact shows the strong association between the availability of fixed behavioral patterns and the adaptive elements in the response to each new situation.
The spiders used the similar attack tactics in response to both Tenebrio and Acromyrmex prey: wrap and then biting. The evolution of the functional aspects of wrapping was discussed by Robinson, Mirick and Turner (1969) and Robinson and Robinson (1976). Wrap-bite is used on several dangerous prey (Robinson & Robinson, 1981). The behavior observed in Metepeira gressa suggests that the spider considers Tenebrio as a dangerous prey, probably due to its strong defensive movements such as wriggling.
The high correlation between the number of units performed during the Immobilization Phase and its duration, indicates that the complexity of the capture behavior on mealworm is caused by the high occurrence of units and not by an increase of duration of units of behavior. The discriminant analysis showed strong association between the total number of units and the number of Detection phase units. This emphasizes the importance of this phase in the discrimination. The Final phase had less variation, and apparently it did not weight in discrimination, because the prey after the immobilization seems to be a package to carry, independently of the degree of difficulty to be immobilize.
I had hypothesized that Acromyrmex would be the most difficult prey, but the results did not support that hypothesis. I suggest that the difficulty present in the capture of Tenebrio, was due to the novelty factor added to the instinctive character of the discrimination. In spite of this the spider present units to immobilize and could eat it. In this study Metepeira gressa has shown a wide repertoire of potential units and sequences of predatory behavior. This spider would be capable of choosing, among a limited set of tactics, the right one according to each type of prey.
References
Ades, C., Viera, C., & Japyassú, H. (1990). Estrategias de caça na aranha Nephilengys cruentata (Araneae, Tetragnathidae) diante de presas diferentes. Em Anais 42a Reunião da Sociedade Brasileira para o Progresso da Ciência (pp. 473-474). [ Links ]
Burgess, J. W., & Uetz, G. W. (1982). Social spacing strategies in spiders. In P. N. Witt & J. Rovner (Eds.), Spider communication: Mechanisms and ecological significance (pp. 317-352). Princeton, MN: Princeton University Press. [ Links ]
Burgess, J. W., & Witt, P. N. (1976). Spiders webs: Design and engineering. Interdiscisplinary. Scientific Revue, 1, 322-335. [ Links ]
Eisner, T., & Dean, J. (1976). Ploy and counterploy in predatory-prey interactions: Orb-weaving spider versus bombardier beetles. Proceedings of the National Academy of Sciences, 73(4), 1365-1367. [ Links ]
Hays, H. E. (1985). Predator- prey interaction: Garden spiders and house flies. Proceedings of Pennsylvania Academy Society, 59, 29-32. [ Links ]
Ibarra-Núñez, G. (1984). La etogénesis de la depredación en las arañas del género Tegenaria (Agelenidae): I. La discriminación de las presas en las ninfas sin experiencia. Folia Entomológica Mexicana, 61, 113-114. [ Links ]
Japyassú, H. F., & Ades, C. (1990). Influência do tamanho da presa na seqüência predatória de Nephilengys cruentata (Araneidae). Em Resumos. Congresso Brasileiro da Zoologia (p. 27). Londrina, PR. [ Links ]
Japyassú, H. F., & Viera, C. (2002). Predatory plasticity in Nephilengys cruentata (Araneae:Tetragnathidae): Relevance for philogeny reconstruction. Behaviour, 139, 529-544. [ Links ]
Legendre, L., & Legendre, P. (1984). Ecologie numerique. Paris: Masson. [ Links ]
Mielle, D. (1978). Contribution a l´etude du comportment prédateur et des mécanismes de toléránce dans le genre Tegenaria (Araneae, Agelenidae). Thésé de 3éme Cycle. Université de Nancy I. [ Links ]
Nentwig, W. (1987). The prey of spiders. In W. Nentwig (Ed.), Ecophysiology of spiders (pp. 249-263). Berlin: Springer-Verlag. [ Links ]
Peters, H. M. (1954). Estudios adicionales sobre la estructura de la red concéntrica de las arañas. Comunicaciones del Instituto Tropical de Investigaciones Científicas de la Universidad de El Salvador, 1, 1-18. [ Links ]
Peters, H. M. (1955). Contribuciones sobre la etología y ecología comparada de las arañas tejedoras tropicales. Comunicaciones del Instituto Tropical de Investigaciones Científicas de la Universidad de El Salvador, 1(2), 37-46. [ Links ]
Riechter, S., & Luczak, J. (1982). Spider foraging: Behavioral responses to prey. In P. N. Witt & J. Rovner (Ed.), Spider communication (pp. 353-386). New Jersey: Princeton University Press. [ Links ]
Robinson, B., & Robinson, M. H. (1981). Ecología y comportamiento de algunas arañas fabricadoras de redes en Panamá: Argiope argentata, A. savignyi, Nephila clavipes y Eriophora fuliginea (Araneae: Araneidae). Academia Panameña de Medicina y Cirugía, 6(1), 90-117. [ Links ]
Robinson, M. H., & Olazarri, J. (1971). Units of behavior and complex sequences in the predatory behavior of Argiope argentata (Fabricius) (Araneae: Araneidae). Smithsonian Contributions of Zoology, 65, 1-36. [ Links ]
Robinson, M. H., Mirick, H., & Turner, O. (1969). The predator behavior of some araneid spiders and the origin of immobilization wrap. Psyche 76, 486-501. [ Links ]
Robinson, M. H., & Robinson, B. (1976). Discrimination between prey types: An innate component of the predatory behavior of araneid spiders. Zeitschrift Tierpsychology, 41, 266-276 [ Links ]
Stowe, M. K. (1986). Prey specialization in the Araneidae. In W. A. Shear (Ed.), Spiders: Webs, behavior and evolution (pp. 101-131). Stanford,MN: Stanford University Press. [ Links ]
Suter, R. B. (1978). Cyclosa turbinata (Araneae: Araneidae): Prey discrimination via web-borne vibrations. Behavior, Ecology and Sociobiology, 3, 283-296. [ Links ]
Uetz, W. G. (1992). Foraging strategies of spiders. Trends in Ecology and Evolution, 7(5), 155-159. [ Links ]
Uetz , G. W., Johnson, A. D., & Schemscke, W. (1978). Web placement, web structure, and prey capture in orb-weaving spiders. Bulletin of Britanic. Arachnology Society, 4, 141-148. [ Links ]
Viera, C. (1983). Comportamiento de captura de Alpaida alticeps (Keyserling, 1879) (Araneae,Araneidae) sobre Metepeira sp. (Hymenoptera, Formicidae). Comunicaciones de las. En III Jornadas de Ciencias Naturales (pp. 12- 14). Uruguay. [ Links ]
Viera, C. (1986). Comportamiento de captura de Metepeira sp A (Araneae, Araneidae) sobre Acromyrmex sp. (Hymenoptera, Formicidae) en condiciones experimentales. Aracnología, 6, 1-8. [ Links ]
Viera, C. (1989). Características de la tela orbicular de Metepeira sp. A (Araneae, Araneidae). Boletín de la Sociedad Zoológica, 5(2a época), 5-6. [ Links ]
Viera, C. (1994). Analisis del comportamiento depredador de Metepeira gressa (Keyserling) (Araneae, Araneidae) en condiciones experimentales. Aracnologia, 8, 1-9. [ Links ]
Viera, C. (1995). Discriminación por Metepeira gressa(Keyserling) (Araneae, Araneidae) en condiciones experimentales sobre dos presas frecuentes en el medio. Journal of Arachnology, 23, 17-24. [ Links ]
Viera, C., &. Costa, F. G. (1985). Captura de presas por machos adultos de Metepeira sp. A (Araneae, Araneidae). Actas Jornadas de Zoologia, 5-7. [ Links ]
Viera, C., & Costa, F. G. (1988). Análisis del comportamiento de captura de presas por machos adultos de Metepeira sp. A (Araneae, Araneidae), utilizando telas de juveniles y hembras adultas co-específicos. Journal of Arachnology, 16, 141-152. [ Links ]
Wise, D. H. (1983). Competitive mechanism in a food-limited species: Relative importance of interference and explotaitive interactions among labyrinth spiders (Araneae: Araneidae). Oecologia, 48, 252-256. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Carmen Viera
Universidad de la República, Facultad de ciencias
Iguá, 4225, 11400, Montevideo, Uruguay.
E-mail: cviera@fcien.edu.uy
Received June 14, 2005
Revision received September 30, 2005
Accepted October 13, 2005
* I am very grateful to F.G. Costa and G. Francescoli for critical review and helpful comments, to M. Sarasola and M. Arim for the help with the discriminant analysis, and to R.B. Suter for his comments.

