











 (pdf)
(pdf)




 Permalink
PermalinkActa Comportamentalia
ISSN 0188-8145
Acta comport. vol.20 no.1 Guadalajara 2012
Artículo
Control espacio-temporal y generalización de la respuesta de palanqueo en ratas
Spatiotemporal control and response generalization of lever pressing by rats
Rogelio Escobar; Carlos A. Bruner1
Universidad Nacional Autónoma de México, Laboratorio de Condicionamiento Operante (México)
Resumen
Se estudió la distribución temporal de presiones reforzadas nominalmente en una palanca y la ocurrencia de presiones en palancas adyacentes usando dos programas de reforzamiento: intervalo fijo 60 s y tándem reforzamiento diferencial de otras conductas 30 s razón fija 1. Se expuso a tres ratas a cada programa en una cámara experimental que contenía siete palancas. Sólo las presiones en la palanca central (operativa) resultaron en comida. Con el programa de intervalo fijo, las presiones en la palanca operativa y en las palancas cercanas a la operativa aumentaron del inicio al final del intervalo entre reforzadores. Bajo el programa de reforzamiento tándem las presiones en la palanca operativa y en las palancas cercanas a la operativa aumentaron durante los últimos segundos del intervalo entre reforzadores. Con ambos programas de reforzamiento se encontró un gradiente de generalización de la respuesta con un máximo en la palanca operativa que no fue diferente del inicio al final del intervalo entre reforzadores. Las respuestas se entrelazaron en patrones repetitivos consistentes intra sujeto pero diferentes entre sujetos. Se concluyó que el responder generalizado sigue una distribución temporal similar a la que se observa en las respuestas que cumplen con el requisito de reforzamiento.
Palabras clave: Generalización de la respuesta, inducción de respuestas, ubicación espacial de la respuesta, control temporal, variabilidad de la respuesta, presiones a la palanca, ratas.
Abstract
The temporal distribution of nominally reinforced lever pressing by rats and the occurrence of unreinforced presses on seven adjacent levers were studied using two schedules of reinforcement: a fixed interval 60 s and a tandem differential reinforcement of other behavior 30 s fixed ratio 1. Three rats were exposed to each schedule of reinforcement in an experimental chamber containing seven adjoining levers. Only presses on the central (operative) lever produced food pellets. With the fixed-interval schedule, response rate on the operative lever and in those levers closest to the operative increased across the inter-reinforcement interval in a familiar "scallop". Under the tandem schedule, lever pressing on the operative and the inoperative levers increased during the last seconds of the inter-reinforcement interval. The relative spatial distribution of responses on the seven levers converged on the operative lever with the two schedules and this pattern did not differ from the beginning to the end of the interreinforcer interval. Both operative- and inoperative-lever pressing intertwined in repetitive patterns that were consistent within subjects but differed between subjects. These findings suggest that generalized responding follows a temporal distribution similar to that observed in responding that fulfill the reinforcement criterion.
Key words: Response generalization, response induction, response location, temporal control, response variability, lever pressing, rats.
Skinner (1938) en su reformulación de la ley del efecto afirmó que cuando una operante es seguida de un estímulo reforzante su fuerza aumenta. En esta afirmación está implícita una definición funcional tanto del reforzador como de la operante. Como un ejemplo, en condiciones de laboratorio se establece un requisito de reforzamiento como presionar una palanca con una cierta fuerza. Una vez que ocurre esta respuesta, se presenta un estímulo. Si por el efecto de la presentación del estímulo la frecuencia de las presiones a la palanca aumenta por encima de su nivel incondicionado de ocurrencia, el responder se define como operante y el estímulo como un reforzador. De esta forma, aunque las respuestas varíen topográficamente de una ocasión a otra, estas forman parte de una clase operante si están bajo el control de una contingencia de reforzamiento común (Catania, 1973, 2007; Segal, 1972).
Los efectos del reforzamiento, sin embargo, no se limitan a las respuestas que cumplen con el criterio para el reforzamiento, sino que se extienden a otras respuestas con las que comparten propiedades topográficas. Por ejemplo, Skinner (1938) demostró que cuando se impone un criterio mínimo de fuerza o de duración para reforzar las presiones a la palanca, el reforzamiento no solamente aumenta la frecuencia de las respuestas que cumplen con este requisito sino también ocurren respuestas que tienen una fuerza o duración menor o mayor que dicho requisito. De acuerdo con Skinner, estas respuestas no especificadas por el experimentador son topográficamente tan cercanas a la respuesta reforzada nominalmente que los efectos del reforzamiento se inducen o generalizan a ellas. Catania (2007) sugirió que estas respuestas generalizadas o inducidas pueden incluirse como parte de los efectos del reforzamiento en una misma clase funcional (véase también Baum, 1995). Debido a que el término de inducción se ha asociado con efectos no especificados del programa de reforzamiento (e.g., Staddon, 1977) en el presente estudio se prefirió usar el término de generalización de respuestas.
La generalización de la respuesta es un proceso que permite a la conducta ajustarse a los cambios en las contingencias de reforzamiento y, por lo tanto, es un aspecto crucial en el moldeamiento de nuevas respuestas (Skinner, 1938; véase también Keller & Schoenfeld, 1950; Segal, 1972). Supongamos que el requisito de reforzamiento es presionar una palanca con una duración de entre 1 y 2 s. Si este requisito produjera respuestas únicamente con estas duraciones, un cambio súbito, aunque mínimo, en el requisito de reforzamiento, digamos a 3 s, resultaría en la extinción de las respuestas. Si, en contraste, ocurren respuestas de diferentes duraciones por un efecto de generalización de la respuesta, estas respuestas irrelevantes para el requisito inicial de reforzamiento producirían el reforzador cuando cambie el requisito. Para algunos autores este proceso asemeja, a nivel conductual, a la selección natural de las especies debido a los elementos comunes de variación y selección (e.g., Baum, 1995; 2005)
Algunos estudios han mostrado algunas propiedades de la generalización de la respuesta. En un estudio, Hefferline & Keenan (1963) expusieron a participantes humanos a un programa de razón fija (RF) 1 que resultaba en dinero como reforzador. Registraron un continuo de amplitud en los movimientos del pulgar y reforzaron sólo los movimientos que cumplían con un requisito tanto mínimo como máximo. Encontraron que la distribución de respuestas se centró en el requisito de reforzamiento de tal forma que las respuestas que cumplieron con el requisito de reforzamiento fueron más frecuentes que las respuestas que excedían o no alcanzaban el requisito. Las respuestas fuera del requisito de reforzamiento fueron menos frecuentes conforme su topografía se alejaba gradualmente de este requisito. Esto es, observaron un gradiente de generalización de la respuesta (véase también Herrick, 1964; Kuch, 1974). Cuando Hefferline y Keenan expusieron a los sujetos a sesiones de extinción, observaron que las respuestas que antes producían el reforzador disminuyeron y las respuestas que antes ocurrían fuera del criterio de reforzamiento aumentaron (véase también Antonitis, 1951; Skinner, 1938). De esta forma, el gradiente de generalización de la respuesta cambió de ser una función bitónica ascendente-descendente de la amplitud en el movimiento a mantenerse relativamente constante en las diferentes amplitudes.
Diferentes estudios han mostrado el efecto de otros parámetros del reforzamiento sobre la ocurrencia de la generalización de la respuesta. Por ejemplo, Escobar y Bruner (2007) mostraron que conforme se alarga la duración de una demora de reforzamiento aumenta la generalización de la respuesta, por lo que, el gradiente de generalización de la respuesta se vuelve cada vez más plano. Di Lollo, Ensminger y Notterman (1965) mostraron que la generalización de la respuesta aumentó conforme la magnitud del reforzamiento disminuyó. De estos estudios puede concluirse que las operaciones que reducen la fuerza de las respuestas dentro del requisito de reforzamiento aumentan la ocurrencia de respuestas fuera de este requisito.
Los efectos de los programas de reforzamiento sobre la distribución temporal de las respuestas que ocurren fuera del requisito de reforzamiento, sin embargo, no son claros. En un estudio, Arntzen, Sagvolden, y Slatta (1993) realizaron un experimento sobre generalización de la respuesta en el que utilizaron un programa de reforzamiento de intervalo fijo (IF) y registraron la ubicación espacial de la respuesta en 20 ranuras circulares usando ratas como sujetos. Los autores entregaron agua como reforzador únicamente cuando las respuestas ocurrían en una de las ranuras centrales (ranura operativa). Encontraron que las respuestas al inicio del intervalo entre reforzadores (IER) ocurrieron principalmente en una de las ranuras laterales lejos de la ranura operativa. Al final del IER las respuestas ocurrieron principalmente en la ranura operativa y en las ranuras cercanas a la operativa (véase también Sagvolden, Slatta, & Arntzen, 1988). Aunque este hallazgo parece ser consistente con la noción de que la porción inicial de un programa de IF puede tener efectos funcionalmente similares a la extinción (Lattal, St. Peter Pipkin, & Escobar, en prensa) que aumentan la generalización de la respuesta (e.g., Hefferline & Keenan, 1963), un aspecto del procedimiento impide hacer esta afirmación. El hecho de que al inicio del IER las respuestas ocurrieran en una ranura alejada de la ranura operativa es difícil de interpretar debido a que el recipiente para el agua se encontraba en uno de los paneles laterales de la cámara experimental cerca de donde ocurrieron estas respuestas. Otro hallazgo que complica la interpretación de estos resultados es que Notterman y Mintz (1965, p. 177-179) reportaron que en programas de IF las respuestas que excedían un criterio mínimo de fuerza de la respuesta ocurrieron con la misma frecuencia al inicio y al final del IER.
En otros estudios se han usado programas de reforzamiento que limitan la tasa de la respuesta que cumple con el criterio de reforzamiento. Por ejemplo, A. Bruner y Revusky (1961) determinaron la distribución temporal de las respuestas de presionar teclas de telégrafo con participantes humanos. Estos autores alinearon horizontalmente cuatro teclas de telégrafo y reforzaron únicamente las presiones en una de las teclas del centro (operativa) conforme a un programa de reforzamiento diferencial de tasas bajas (RDB) 8.2 s. Las presiones en las tres teclas restantes (inoperativas) no tenían consecuencias programadas. Encontraron que las respuestas en la tecla operativa se espaciaron conforme a la duración del programa de reforzamiento. Las presiones en las teclas inoperativas, en comparación, se organizaron en patrones repetitivos dentro del intervalo entre respuestas en la tecla operativa. Estos patrones fueron consistentes para cada sujeto pero fueron diferentes entre sujetos. Estos resultados sugieren que las presiones en las teclas inoperativas "llenan" el tiempo en el que las respuestas dentro del requisito de reforzamiento no ocurre y funcionan a la manera de marcadores de tiempo facilitando el espaciamiento de la respuesta operativa (véase Wilson & Keller, 1952).
No todos los estudios sobre generalización de respuestas han mostrado que las respuestas que no cumplen con el requisito de reforzamiento ocurren cuando la frecuencia de las respuestas que cumplen con el requisito es baja. Por ejemplo, en un estudio reciente, Escobar y Bruner (2007) estudiaron la generalización de la respuesta registrando la ubicación espacial de la respuesta en siete palancas alineadas horizontalmente. Sólo las presiones en la palanca central produjeron comida conforme a un programa de intervalo al azar (IA) 32 s. La distribución temporal de las presiones en la palanca operativa y en las palancas cercanas a la operativa fue similar a lo largo del intervalo entre reforzadores. Se observó una breve pausa post reforzamiento y después un aumento súbito en el número de respuestas que alcanzó un nivel asintótico y se mantuvo relativamente constante el resto del intervalo entre reforzadores. Es incierto, sin embargo, si este hallazgo es característico de los programas de IA o si es una propiedad de la generalización de respuestas independientemente del programa de reforzamiento que se emplee.
En el estudio de Escobar y Bruner (2007) y en el de Arntzen et al. (1993) las respuestas reforzadas nominalmente en la palanca operativa podían ocurrir libremente durante el IER. En comparación, A. Bruner y Revusky (1961) usaron programas que castigan negativamente las respuestas en una tecla o en una palanca operativa (RDB). A pesar de que aparentemente la restricción temporal de las respuestas reforzadas nominalmente podría ser responsable de las diferencias en la distribución temporal de las respuestas tanto dentro como fuera del requisito de reforzamiento, las diferencias en los procedimientos impiden hacer una conclusión sólida. Por lo tanto, en el presente se determinó si las respuestas que ocurren fuera del requisito espacial de reforzamiento se organizan en patrones temporales similares a los que se observan en las respuestas que cumplen con el requisito de reforzamiento usando dos programas de reforzamiento comparables: un programa de IF (que puede conceptualizarse como tándem TF RF 1) y un tándem RDO RF1. Este último es similar a un programa de RDB pero requiere solamente una respuesta para entregar el reforzador igual que un programa de IF. La diferencia con el programa de IF radica en que en el tándem RDO RF1 las respuestas que ocurren antes de la respuesta procuradora del reforzador reinician el contador del programa (i.e., se castigan negativamente), mientras que en el IF no tienen consecuencias programadas.
Método
Sujetos
Se utilizaron 6 ratas macho Wistar de tres meses de edad al inicio de la investigación y sin experiencia experimental. Las ratas se alojaron en cajas habitación individuales donde tuvieron acceso libre al agua. Durante todo el experimento se restringió el acceso a la comida para mantenerlas al 80% de su peso ad libitum. Las ratas se obtuvieron del bioterio de la Facultad de Psicología de la Universidad Nacional Autónoma de México y se mantuvieron conforme a los criterios éticos de la institución.
Aparatos
Se utilizó la cámara experimental descrita por Escobar y Bruner (2007). La cámara experimental medía 26 cm de alto por 29 cm de largo y 27 cm de fondo y se fabricó con dos paredes de acrílico y dos paneles de acero inoxidable. En el panel frontal a 5 cm del piso de la cámara se colocó una hilera de siete palancas alineadas horizontalmente con una separación de 1 mm entre sí. Cada palanca medía 1 cm de alto por 1.8 cm de ancho y entraba 2.5 cm en la cámara experimental. Se requería una fuerza de 0.15 N para cerrar el interruptor de cada palanca. En el centro del panel opuesto se colocó un comedero de lámina conectado a un dispensador de bolitas de comida BRS/LVE (Modelo DDH-020) que dejó caer bolitas de 25 mg, fabricadas remoldeando comida pulverizada para ratas. Arriba del comedero, se colocó un foco que proporcionó la iluminación general de la cámara. La cámara experimental se introdujo en una caja de madera sonoamortiguada equipada con un ventilador.
El control y registro de los eventos experimentales se realizó desde un cuarto adyacente mediante una computadora IBM acoplada a la cámara experimental con una interfaz Advantech PC-LabCard (modelo PCL-725) usando programas escritos en GW-BASIC.
Procedimiento
Todas las ratas recibieron una sesión de entrenamiento a comer en la que se les entregaron bolitas de comida dentro de la cámara experimental hasta que las ratas se aproximaron consistentemente al comedero y consumieron 50 de estas bolitas.
Para facilitar la descripción del procedimiento y de los resultados, las palancas se numeraron de 1 a 7 respecto a su posición de izquierda a derecha vistas del interior de la cámara experimental. Debido a que estudios previos mostraron que la exposición a programas de RDB antes de la exposición a programas de IF en ratas como sujetos resulta en tasas de respuesta notablemente bajas (Tatham, Wanchisen, & Yasenchack, 1993), se utilizó un diseño entre grupos. Sin otro tipo de entrenamiento, para tres ratas (F1, F2 y F3) se utilizó un programa de reforzamiento de IF 60 s vigente en la palanca 4 (operativa). En las otras tres ratas (D1, D2 y D3) se utilizó un programa de reforzamiento tándem RDO 30 s RF 1. La duración del programa de RDO se seleccionó con base en pruebas previas a este experimento para igualar la frecuencia de reforzamiento obtenida en los dos programas de reforzamiento. Las presiones en las palancas 1, 2, 3, 5, 6 y 7 (inoperativas) se registraron pero no tuvieron consecuencias programadas. Los programas de reforzamiento estuvieron vigentes durante 30 sesiones que terminaban después de de 1 hora o de entregar 50 reforzadores (50 bolitas de comida). Las sesiones experimentales se llevaron a cabo siete días por semana y los sujetos se introdujeron a la cámara experimental siempre en el mismo orden.
Resultados
La tasa de respuesta individual en cada palanca y la frecuencia de reforzamiento obtenida bajo el programa de reforzamiento de IF y bajo el programa tándem RDO 30 s RF 1 se muestran, respectivamente, en las Tablas 1 y 2. La tasa de respuesta fue mayor con el programa de IF que con el tándem RDO RF 1. La tasa de reforzamiento fue cercana a un reforzador por minuto para todas las ratas. Para la mayoría de las ratas bajo ambos programas de reforzamiento, la tasa de respuesta fue mayor en la palanca 4 que en el resto de las palancas. La excepción fue la Rata D3 en la que se encontró que la tasa de respuesta fue mayor en la palanca 7.
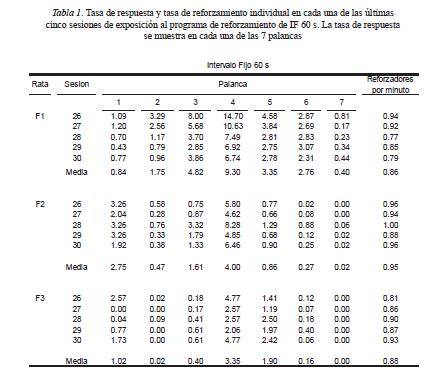
La Figura 1 muestra la distribución temporal de las presiones en las 7 palancas dentro del intervalo entre reforzadores obtenido. La abscisa en cada panel muestra el tiempo que transcurrió durante el IER expresado como un porcentaje de la duración total de cada IER. La ordenada muestra la tasa de respuestas en cada una de las siete palancas durante cada subintervalo del IER como un porcentaje de la tasa máxima de respuesta. Para las tres ratas bajo el programa de IF se encontró que las presiones en la palanca fueron relativamente infrecuentes durante la primera mitad del IER y se observó una aceleración en el último tercio del IER. Para las Ratas F1 y F2 se encontró que en las palancas inoperativas cercanas a la operativa, el patrón de responder siguió la misma distribución temporal pero con una tasa más baja que en la palanca operativa. En la Rata F3 se encontró un aumento de aproximadamente la mitad del IER al final de este intervalo en las palancas 1 y 3. Para las Ratas D1 y D2 bajo el programa de reforzamiento tándem RDO RF 1, las presiones en la palanca operativa aumentaron y después disminuyeron durante los primeros 9 subintervalos del IER. Durante el último subintervalo, se observó un aumento notable en las presiones en la palanca operativa. dos subintervalos del IER. Se eligieron dos subintervalos a partir de la inspección visual de la Figura 1 para analizar las distribuciones espaciales cuando las presiones en todas palancas fueron aparentemente similares. Las respuestas se muestran como porcentajes para facilitar las comparaciones en la distribución espacial de las respuestas independientemente de los cambios en la tasa de respuesta.
Este aumento se observó también en las palancas cercanas a la operativa estas dos ratas. Para la Rata D3 se encontró que las presiones en la palanca operativa aumentaron ligeramente al final del IER, mientras que en la palanca 7 las presiones fueron relativamente altas durante los primeros 8 subintervalos del IER y disminuyeron durante los últimos dos subintervalos. Un patrón similar aunque con una tasa notablemente baja se observó en la palanca 5.
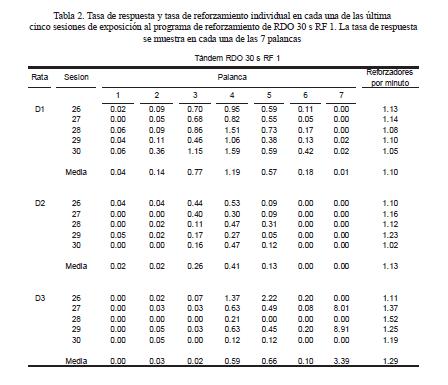
Para analizar los cambios en la distribución espacial de las respuestas al inicio y al final del IER, la Figura 2 muestra el número de respuestas en cada palanca como un porcentaje de la tasa global de respuestas en las siete palancas. Los dos paneles de la izquierda muestran los datos con el programa de reforzamiento de IF y los dos paneles de la izquierda muestran los datos con el programa de reforzamiento tándem RDO 30 s RF s. Bajo cada programa de reforzamiento se analizaron las respuestas durante los primeros y los últimos
Para todas las ratas con el programa de reforzamiento de reforzamiento de IF y para las Ratas D1 y D2 con el programa tándem RDO 30 s RF 1 se encontró que las presiones más frecuentes ocurrieron en la palanca operativa. En general, para las Ratas F1, F2, D1 y D2, las presiones en las palancas inoperativas disminuyeron conforme las palancas se encontraban más alejadas de la palanca operativa. No se encontraron cambios sistemáticos entre el inicio y el final del IER. Para la Rata F3 con el programa de IF se encontró que ocurrió un porcentaje de presiones similar en las palancas 1 y 3 pero tampoco se encontraron diferencias sistemáticas entre el inicio y el final del IER. Para la Rata D3 con el programa de reforzamiento tándem RDO RF 1 se encontró que al inicio del IER las presiones fueron notablemente más frecuentes en la palanca 7 que en el resto de las palancas. En las palancas 4 y 5 se encontró un porcentaje de respuestas cercano a cero. Las presiones en la palanca 4 aumentaron y en la palanca 7 disminuyeron durante el final del intervalo entre reforzadores.
Para determinar si las distribuciones espaciales y los patrones temporales de las presiones en las siete palancas que se observaron en las Ratas F3 y D3 se debieron a alguna organización secuencial de las presiones a la palanca, se analizó la probabilidad de ocurrencia de pares de eventos (presiones en las palancas y entrega de reforzadores) y se determinó si estas probabilidades fueron consistentes para cada sujeto. Con la frecuencia de cada presión en todas las palancas (eventos 1 a 7) o la entrega del reforzador (evento ER), se calculó la probabilidad de que una presión en cualquiera de las siete palancas o un reforzador fuera seguido por una presión en alguna otra palanca (sólo se contaron los cambios de palanca) o por un reforzador (véase Escobar & Bruner, 2007). Por ejemplo, si se observó el siguiente patrón de eventos, 4-4-5-4-6-ER, se contarían los pares 4-5, 5-4, 4-6, y 6-ER y se les asigna una probabilidad a partir del total de pares de eventos en una sesión. Con fines de comparación intra y entra sujetos, este dato se calculó en las dos últimas sesiones bajo cada uno de los dos programas de reforzamiento para todas las ratas y se muestra en la Figura 3. Para enfatizar la regularidad de los patrones que formaron las secuencias de dos eventos, cada celda con un valor mayor de cero aparece sombreada.
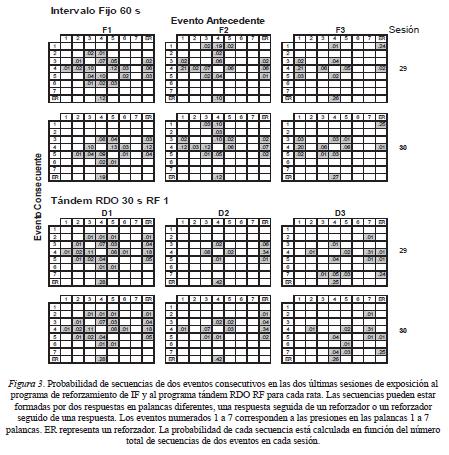
Se encontró que los patrones que forman las probabilidades de pares de eventos fueron consistentes intra sujeto pero fueron diferentes entre sujetos. Para las Ratas F1, F2, D1 y D2 se encontró que los pares de eventos más frecuentes incluyeron una presión en la palanca operativa y una presión en una palanca cercana a la operativa o un reforzador (e.g., 4-ER, ER-4, 5-4, 4-5, 4-3 y 3-4). En la Rata F3 se encontró que los pares de eventos más frecuentes incluyeron una presión en la palanca 4 y una presión en la palanca 1 y una presión en la palanca 4 seguida de un reforzador. En la Rata D3 los pares de eventos más frecuentes incluyeron presiones en las palancas 4 y 7 y una presión en la palanca 4 seguida de un reforzador.
Discusión
Se encontró que para los tres sujetos expuestos al programa de reforzamiento de IF y para dos sujetos (D2 y D3) expuestos al programa tándem RDO RF, la distribución espacial de respuestas en las siete palancas replicó el gradiente de generalización de respuestas (e.g., Escobar & Bruner, 2007; Hefferline & Keenan, 1963; Herrick, 1964; Kuch, 1974). Esto es, las respuestas alcanzaron un máximo que coincidió con el requisito de reforzamiento y disminuyeron conforme las respuestas se alejaron de dicho requisito.
Para los mismos cinco sujetos (F1, F2, F3, D1, D2) se encontró que la distribución temporal de las presiones en las palancas inoperativas cercanas a la operativa reprodujo la distribución temporal de las respuestas en la palanca operativa. Un hallazgo notable es que con el programa de IF, en las palancas inoperativas cercanas a la operativa, se encontraron "festones" con un nivel asintótico menor al encontrado en la palanca operativa. Este hallazgo es congruente con el resultado de Arntzen et al. (1993; véase también, Sagvolden et al., 1988) de que al final del IER en un programa de IF las respuestas en una ranura operativa y en las ranuras cercanas a la operativa fueron más frecuentes que las respuestas en el resto de las ranuras. Diferente del estudio de Arntzet et al., sin embargo, en el presente estudio el patrón espacial de respuestas no fue diferente en el inicio o en el final del IER. Este hallazgo sugiere que la posición espacial del recipiente para el agua en el estudio de Artzen et al. fue responsable de que al inicio del IER ocurrieran respuestas en las ranuras cercanas al recipiente. En contraste, este resultado del presente estudio es congruente con los hallazgos de Notterman y Mintz (1965, p. 177-179) de que la distribución en la fuerza de la respuesta no varió del inicio al final del IER usando programas de reforzamiento de IF con un criterio mínimo de fuerza. A pesar de que hubiera sido esperable que debido al efecto local del inicio del IER como extinción (Lattal et al., en prensa) la distribución espacial de las respuestas al inicio del IER fuera relativamente plana bajo ambos programas de reforzamiento (cf. Hefferline & Keenan, 1963), los hallazgos del presente estudio sugieren que desde el inicio del IER se establece un patrón característico de respuestas que aumenta en frecuencia a partir, aproximadamente, de la mitad del IER.
Los resultados sugieren que tanto las respuestas que cumplen con el requisito de reforzamiento como las respuestas que no alcanzan a cumplir con el requisito se distribuyen de la misma forma dentro de un IER. Este hallazgo es consistente con la explicación de Catania (2007), de que tanto las respuestas que cumplen con el requisito de reforzamiento como las respuestas topográficamente cercanas al requisito pueden considerarse dentro de una misma clase funcional que está controlada por los efectos del reforzamiento.
Esta última afirmación podría parecer contradictoria con los análisis de las conductas inducidas por el programa (CIP) (e.g., Staddon, 1977). Las CIP ocurren cuando se entrega reforzamiento intermitente dependiente o independiente de la respuesta (e.g., IF o TF por comida). Estas conductas ocurren en diferentes porciones del IER y algunos autores las han caracterizado como producto de diferentes "estados motivacionales" o "emocionales" durante este intervalo (e.g., Staddon, 1977; Timberlake & Lucas, 1991). Staddon clasificó a las CIP en términos de su ubicación dentro del IER. Por ejemplo, si dentro de la cámara experimental se añade una pipeta con agua, los sujetos (ratas o palomas) beben agua durante el inicio del IER donde la comida no ocurre. A la conducta de beber la clasifico como interina debido a que ocurre al inicio del IER. Otras conductas inducidas pueden ocurrir al final del IER. Por ejemplo, acercarse al comedero o picar la pared del comedero, de acuerdo con Staddon, son CIP que por su ubicación en el IER se clasifican como terminales. A partir de los hallazgos del presente estudio puede sugerirse que las conductas caracterizadas como terminales podrían ser parte de un gradiente de generalización de la respuesta.
Si bien Staddon (1977) describió que las conductas terminales ocurren con programas de reforzamiento independiente de la respuesta, aún con estos programas es necesario que los sujetos se aproximen al comedero y consuman el alimento. Debido a que la entrega de alimento aumenta la frecuencia de estas conductas, hacer contacto con el comedero puede caracterizarse como una operante reforzada con el alimento. Los resultados del presente experimento sugieren que la conducta de picar para consumir la comida podría generalizarse a otras ubicaciones espaciales fuera del comedero y esta conducta seguiría un patrón temporal similar al de la conducta que cumple con el criterio de reforzamiento. Esta interpretación, aunque aun necesita de más evidencia empírica, hace innecesaria la caracterización de las conductas terminales como inducidas por el programa y diferentes de la clase operante. Aunque los datos del presente experimento no permiten hacer afirmaciones acerca de las conductas interinas, otros autores han sugerido que podrían estar mantenidas por un reforzador diferente al que mantiene a la operante blanco (véase Pierce & Cheney, 2004; Roca & Bruner, 2011).
Escobar y Bruner (2007), reportaron que la distribución temporal de las presiones palancas fue similar en las 7 palancas durante el IER usando programas de IA. Este hallazgo es congruente con los resultados del presente estudio pero es contradictorio con los resultados de A. Bruner y Revusky (1961). A. Bruner y Revusky encontraron con programas de reforzamiento de RDB, que las presiones en tres teclas inoperativas ocurrieron durante los periodos en los cuales la respuesta que cumplió con el requisito de reforzamiento fue infrecuente.
El patrón temporal de respuestas observado bajo el programa de reforzamiento tándem RDO RF1 en la Rata D3 difirió notablemente del resto de los sujetos y es consistente con los hallazgos de A. Bruner y Revusky (1961). Por un lado, la distribución espacial de las respuestas en las siete palancas alcanzó un máximo en la palanca inoperativa 7 y sólo se observaron algunas respuestas en las palancas 4 y 5. Por otro lado, la distribución temporal de las presiones en las palancas mostró que las presiones en la palanca 7 predominaron durante el IER y sólo aumentaron en la palanca operativa (4) durante el último subintervalo del IER. Este resultado sugiere que se estableció supersticiosamente una cadena de respuestas en dos palancas (4 y 7) (e.g., Catania, 1966).
Wilson y Keller (1952) ofrecieron una explicación similar en el caso de conducta colateral a la reforzada en el intervalo entre reforzadores. Estos autores observaron patrones repetitivos de conducta como aproximarse a un bebedero o lamerse durante el IER con programas de RDB. Los patrones fueron consistentes intra sujeto pero diferentes para cada sujeto (véase también Laties, Weiss, & Weiss, 1966). Los autores sugirieron que esta conducta colateral a la reforzada formó una cadena con la conducta reforzada. Si la conducta colateral ocurría durante un periodo de tiempo mayor al requisito de reforzamiento, entonces esta conducta seguida de la respuesta especificada era reforzada. En este sentido, la conducta colateral facilitó el espaciamiento de las respuestas reforzadas y podría sugerirse que funcionó como un "marcador de tiempo".
La interpretación de Wilson y Keller (1952) es congruente con los resultados obtenidos en la Rata D3 en la que se observó que los patrones secuenciales de dos eventos mostraron que las secuencias más frecuentes involucraron presiones en la palanca 7 seguidas por presiones en la palanca 4 y después por el reforzador (7-4, 4-ER, ER-7). Es importante señalar que la probabilidad de que una respuesta en la palanca 4 fuera seguida de una presión en la palanca 7 fue notablemente baja, lo cual sugiere que las secuencias de respuestas no consistieron únicamente de repeticiones 4-7 y 7-4 durante el IER, sino de repeticiones en la palanca 7 seguidas de una respuesta en la palanca 4. Al igual que en los estudios de A. Bruner y Revusky (1961) y de Wilson y Keller, las respuestas en la palanca 7, al establecerse supersticiosamente, parecieron facilitar el espaciamiento de las respuestas en la palanca 4. En conclusión cuando accidentalmente se refuerza un patrón que incluye tanto a las respuestas en operandos inoperativos como a las respuestas en el operando operativo, la distribución temporal de las respuestas inoperativas dentro del IER puede diferir de la distribución de la respuesta operativa.
Algunos autores (e.g., Baum, 1995; 2005) han considerado a la generalización de la respuesta como un símil a escala individual de la variación y selección natural de las especies. Las similitudes son evidentes dado que las variaciones en la topografía de la operante permiten el ajuste de las respuestas a los cambios en las contingencias de reforzamiento de manera similar a como las variaciones en una especie permiten la supervivencia de algunos miembros ante la presión selectiva. Una operante se establece por medio del reforzamiento diferencial (selección conductual) de manera análoga a como se forma una nueva especie a partir de la selección natural (e.g., Glenn, Ellis, & Greenspoon, 1992). Si bien la analogía es, al menos, intrigante, los diferentes niveles de observación oscurecen la interpretación de las similitudes entre el reforzamiento diferencial y la selección natural. En el estado actual de conocimiento, en el que aún es dudosa la contribución de dicha metáfora al conocimiento en análisis de la conducta, sería especulativo argumentar por un proceso común a ambos niveles de selección (cf., Skinner, 1981). Queda únicamente decir que es necesario realizar más investigación sobre el fenómeno de la generalización de respuestas para determinar la utilidad de la metáfora de la selección natural en el análisis experimental de la conducta.
Referencias
Antonitis, J. J. (1951). Response variability in the white rat during conditioning, extinction, and reconditioning. Journal of Experimental Psychology, 42, 273–281.
Arntzen, E., Sagvolden, T., & Slatta, K. (1993). Methylphenidate hydrochloride (Ritalin) reduces operant responding in rats by affecting the spatial and temporal distributions of responses. The Psychological Record, 43, 223-233. [ Links ]
Baum W. M. (1995). Introducción al análisis molar de la conducta. Revista Mexicana de Análisis de la Conducta, 21, 7-25. [ Links ]
Baum W. M. (2005). Understanding behaviorism: Behavior, culture, and evolution (2a ed.) Malden, MA: Blackwell [ Links ]
Bruner, A., & Revusky, S. H. (1961). Collateral behavior in humans. Journal of the Experimental Analysis of Behavior, 4, 349–350.
Catania, A. C. (1966). Concurrent operants. In W. K. Honig (Ed.), Operant behavior: Areas of research and application (pp. 213-270). New York: Appleton-Century-Crofts. [ Links ]
Catania, A. C. (1973). The nature of learning. In J. A. Nevin & G. S. Reynolds (Eds.), The study of behavior: Learning, motivation, emotion and instinct (pp. 31-68). Glenview, IL: Scott, Foresman. [ Links ]
Catania A. C. (2007). Learning (4a ed.). Cornwall-on-Hudson, NY: Sloan Publishing. [ Links ]
Di Lollo, V., Ensminger, W. D., & Notterman, J. M. (1965). Response forcé as a function of amount of reinforcement. Journal of Experimental Psychology, 70, 27-31. [ Links ]
Escobar, R. & Bruner, C. A. (2007). Response induction during the acquisition and maintenance of lever pressing with delayed reinforcement. Journal of the Experimental Analysis of Behavior, 88, 29-49. [ Links ]
Glenn, S. S., Ellis, J., & Greenspoon, J. (1992). On the revolutionary nature of the operant as a unit of behavioral selection. American Psychologist, 47, 1329-1336. [ Links ]
Hefferline, R. F., & Keenan, B. (1963). Amplitude-induction gradient of a small-scale (covert) operant. Journal of the Experimental Analysis of Behavior, 6, 307–315.
Herrick, R. M. (1964). The successive differentiation of a lever displacement response. Journal of the Experimental Analysis of Behavior, 7, 211–215.
Keller, F. S., & Schoenfeld, W. N. (1950). Principles of psychology. New York: Appleton-Century-Crofts. [ Links ]
Kuch, D. O. (1974). Differentiation of press durations with upper and lower limits on reinforced values. Journal of the Experimental Analysis of Behavior, 22, 275–283.
Laties, V. G., Weiss, B., & Weiss, A. B. (1969). Further observations on overt "mediating" behavior and the discrimination of time. Journal of the Experimental Analysis of Behavior, 12, 43-57. [ Links ]
Lattal, K.A., St. Peter Pipkin, C., & Escobar, R. (in press). Operant extinction: Elimination and generation of behavior. In G. J. Madden (Ed.-en-Jefe), W. V. Dube, T. D. Hackenberg, G. P. Hanley, & K. A. Lattal (Eds. Asoc.), APA handbooks in psychology. APA handbook of behavior analysis, Vol. 2: Translating core principles into practice. Washington, DC: American Psychological Association. [ Links ]
Notterman, J. M., & Mintz, D. E. (1965). The dynamics of response. New York: Wiley & Sons. [ Links ]
Pierce, W. D., & Cheney, C. D. (2004). Behavior Analysis and Learning (3a ed). Mahwah, NJ: Lawrence Erlbaum Associates. [ Links ]
Roca, A., & Bruner, C. A. (2011). Un análisis del origen del consumo excesivo de agua del beber inducido por el programa. Revista Mexicana de Análisis de la Conducta, 37, 177-204. [ Links ]
Sagvolden, T., Slatta, K., & Arntzen, E. (1988). Low doses of methylphenidate (Ritalin) may alter the delay-of-reinforcement gradient. Psychopharmacology, 95, 303-312. [ Links ]
Segal, E. F. (1972). Induction and the provenance of operants. In R. M. Gilbert, & J. R. Millenson (Eds.), Reinforcement: Behavioral analyses (pp. 1–34). New York: Academic Press.
Skinner, B. F. (1938). The behavior of organisms: An experimental analysis. New York: Appleton-Century-Crofts. [ Links ]
Skinner, B. F. (1981). Selection by consequences. Science, 213, 501-504. [ Links ]
Staddon, J. E. R. (1977). Schedule-induced behavior. In W. K. Honig & J. E. R. Staddon (Eds.), Handbook of operant behavior (pp. l25-152). Englewood Cliffs, NJ: Prentice-Hall. [ Links ]
Tatham, T. A., Wanchisen, B. A., & Yasenchack, M. P. (1993). Effects of differential-reinforcement-of-low-rate schedule history on fixed-interval responding. The Psychological Record, 43, 289-297. [ Links ]
Timberlake, W., & Lucas, G. A., (1991). Periodic water, interwater interval, and adjunctive behavior in a 24-hour multi-response environment. Animal Learning and Behavior, 19, 369-380. [ Links ]
Wilson, M. P. and Keller, F. S. (1953). On the selective reinforcement of spaced responses. Journal of Comparative and Physiological Psychology, 46, 190-193. [ Links ]
Received: August 22, 2011
Accepted: September 09, 2011
1 El primer autor agradece a G. Mayela Hernández Ruiz y a Nadia M. Santillán Bernal por su ayuda en la elaboración de las tablas y las figuras. Dirigir correspondencia a: Rogelio Escobar (rescobar@servidor.unam.mx). Laboratorio de Condicionamiento Operante, Facultad de Psicología, UNAM. Av. Universidad 3004, Col. Copilco – Universidad, C.P. 04510. Del. Coyoacán, México, D.F.
