











 (pdf)
(pdf)




 Permalink
PermalinkActa Comportamentalia
ISSN 0188-8145
Acta comport. vol.21 no.1 Guadalajara 2013
ARTÍCULOS
Efectos de variar la posición espacial de los estímulos contextualizado y contextualizador sobre el ajuste diferencial
Effects of varying the spatial position of the contextualized and contextualizing stimulus on the differential adjustment
Felipe Patrón Espinosa1; Carlos Torres Ceja; Carlos Flores Aguirre
Universidad de Guadalajara-CEIC (México)
RESUMEN
Dos estudios fueron realizados para evaluar el efecto de variar la localización de eventos de estímulo no contingentes (orden y lugar en el que ocurrieron) sobre el ajuste diferencial. En el primer estudio ocho ratas fueron expuestas a tres condiciones experimentales que se caracterizaron por la manipulación de las coordenadas espaciales en las que se entregó el agua (Espacio Fijo, Secuencial y Aleatorio) usando un programa de Tiempo Fijo 20s. En el segundo experimento, ocho ratas fueron expuestas a condiciones similares a las del primer experimento, con la diferencia que en este estudio se presentó una señal luminosa asociada a la localización de la entrega de agua. Los datos fueron analizados como un caso de la Función Contextual propuesta por Ribes y López (1985). Los resultados sugieren que el ajuste diferencial es afectado por: (a) la consistencia de la ocurrencia de estímulos en coordenadas espaciales fijas a través del tiempo; (b) la ocurrencia de la señal luminosa en conjunción con la presentación del agua, (c) la contigüidad espacial entre dichos estímulos. Se propusieron diversas unidades de medida con el fin de identificar el ajuste diferencial.
Palabras clave: ubicación espacial, estímulo contextualizado, contextualizador, ajuste diferencial, ratas.
ABSTRACT
Two studies were conducted to evaluate the effect of varying the spatial location of non-contingent stimuli events (in order and place of occurrence) on differential adjustment. In the first study, eight rats were exposed to three experimental conditions that were characterized by the manipulation of the spatial coordinates in which water was delivered (fixed, sequential or random spatial coordinates) using a Fixed Time 20s schedule. In the second experiment, eight rats were exposed to conditions similar to those in the first experiment, but now there was a signal stimulus correlated with the location of water delivery. Data were analyzed as a case of contextual function proposed by Ribes & López (1985). The results suggest that the differential adjustment is affected by: (a) the consistency of the occurrence of stimuli in fixed spatial coordinates through time; (b) the conjoint occurrence of signal and water delivery location; and (c) the spatial contiguity between these stimuli. Several measurement units to identify the differential adjustment related to contextual function were proposed.
Keywords: spatial location, contextualized, contextualizing stimulus, differential adjustment, rats.
Dentro de la propuesta interconductual iniciada por Kantor (1959/1978) se concibe a los eventos psicológicos como fenómenos naturales, al igual que cualquier otro evento que sea estudiado por una disciplina científica. Según este autor, los eventos psicológicos ocurren en coordenadas espacio-temporales, y por lo tanto, para una explicación de éstos no es necesario recurrir a entidades extraespaciales. Por el contrario, su explicación puede ser estructurada a partir del análisis funcional de los eventos que lo conforman y los cuales ocurren de igual manera en coordenadas espacio-temporales. Ribes y López (1985), a partir de la lógica de campo formulada por Kantor, propusieron una taxonomía en la que se concibe a los fenómenos psicológicos como una función o estructura de relaciones de contingencia (interdependencia) entre eventos de respuesta y de estímulo. Dicha taxonomía se conforma por cinco funciones de complejidad creciente e inclusiva (Contextual, Suplementaria, Selectora, Sustitutiva Referencial y Sustitutiva no Referencial). Cada una de estas funciones psicológicas evidencia como el comportamiento de los organismos se vuelve autónomo de sus determinantes biológicas a través de su interacción circunstancial con los eventos y objetos de su entorno.
El presente trabajo se centra exclusivamente en la función contextual, la cual se caracteriza por ser una interacción en la que la actividad de un organismo es mediada por las relaciones entre eventos que ocurren de forma independiente a ésta en coordenadas espacio-temporales consistentes en el entorno. Ribes (2004) señaló que esta función ejemplifica una relación de isomorfismo en la medida en que constituye un ajuste diferencial del organismo a los cambios en las contingencias entre estímulos. En otras palabras, el organismo reproduce conductualmente alguna propiedad de las contingencias experimentadas. A partir de lo anterior, el ajuste diferencial en esta función implicaría que la distribución en espacio y tiempo de la conducta se ve modulada por las propiedades de ocurrencia de los eventos de estímulo.
En un primer momento, Ribes y López (1985) describieron las funciones psicológicas a partir de casos paradigmáticos diferenciados por los elementos que constituyen cada una de éstas. Posteriormente, Ribes (1995, 2004) incluyó en la estructuración de estas funciones la diferenciación entre contingencias de ocurrencia y de función. La primeras hacen referencia a las condiciones de necesidad y suficiencia en la ocurrencia física de un evento. Por su parte, las contingencias de función describen las propiedades circunstanciales que se establecen entre propiedades de los objetos de estímulo y de las respuestas del organismo como efecto de las contingencias de ocurrencia consistentes. De acuerdo con Peña (comunicación personal), este tipo de contingencias prescriben las relaciones de dependencia (como en toda contingencia) entre las funciones de los elementos (de estímulo y/o de respuesta) relacionados.
Genéricamente, la función contextual pudiera describir relaciones entre eventos de estímulo de los cuales uno de éstos se caracterizan por tener una relación directa con una respuesta, a decir de Ribes y López (1985) "... tiene la capacidad de producir una forma especifica de reactividad..." (pp. 67-68). Este tipo de estímulo es conceptualizado como mediador o contextualizador de la contingencia, en tanto que de éste depende la estructuración de la función. Por su parte, el otro evento de estímulo de la relación se prescribe como estímulo contextualizado debido a que su funcionalidad es dependiente de la relación consistente con el primero. Las relaciones de ocurrencia entre estos estímulos promueve se desarrollen contingencias de función en las que la respuesta asociada al estímulo mediador comience a desligarse en tiempo y espacio de éste y se relacione con el estímulo contextualizado de manera circunstancial.
Ribes (2007) describió una serie de casos funcionales para cada una de las funciones psicológicas basándose en la constancia o variabilidad de la ocurrencia en coordenadas espaciotemporales de los elementos que integran cada función. Estos casos funcionales proveen de una herramienta heurística en la labor de identificar cómo las relaciones definidas por su ubicación espacio-temporal de los eventos de estímulo y de respuesta influyen sobre la estructuración del fenómeno psicológico. El presente trabajo se centró en el estudio de las condiciones de variación en la ocurrencia de eventos de estímulo a partir de distintas coordenadas espaciales y su efecto en la estructuración del comportamiento psicológico. Suponemos que la ocurrencia en espacio de los eventos de estímulo se distingue por: (a) su presentación distal o proximal con respecto a otro evento de estímulo o de respuesta y, (b) por su ocurrencia en un orden espacial-secuencial con respecto a un evento de estímulo o de respuesta. De aceptarse lo anterior, las condiciones de ocurrencia de eventos de estímulo, según sus propiedades espaciales, se podrían dividir en: (a) la ocurrencia de eventos en Espacio Fijo, que refiere la ocurrencia de un evento de estímulo en un mismo lugar a través del tiempo; (b) Espacio Secuencial que hace referencia a que los eventos de estímulo ocurren en distintas ubicaciones espaciales pero con un orden secuencial a través del tiempo, y por último, (c) Espacio Variable que incluye los casos en los que los eventos de estímulo ocurren en distintas ubicaciones espaciales y en un orden aleatorio a través del tiempo.
A partir de la identificación de estas condiciones de ocurrencia de los eventos de estímulo y en consonancia con lo planteado por Ribes (2007), puede suponerse que la estructuración de la función contextual se verá afectada de manera diferencial por cada una de estas manipulaciones. En este contexto, el propósito del presente trabajo fue explorar el efecto de variar la ubicación espacial en la que ocurren los eventos de estímulo no contingentes a la conducta del organismo sobre la distribución espacio-temporal de la actividad del organismo. Consideramos que la distribución espacio-temporal de la actividad del organismo se verá modulada por la presentación de eventos de estímulo en diferentes coordenadas espaciales, permitiendo así identificar si el organismo se comportó diferencialmente ante dichos eventos. De esta manera, se desarrolla un primer experimento con el fin de evaluar el efecto de variar la ubicación espacial en la que se presenta un estímulo contextualizador (agua) no contingente a la conducta del organismo sobre el ajuste diferencial. La preparación de este experimento se basa en el caso de interrupción propuesto por Ribes y López (1985) para la función contextual.
EXPERIMENTO 1
MÉTODO
Sujetos
Se emplearon ocho ratas hembras de la cepa Wistar, de 3 meses de edad e ingenuas al inicio del experimento. Se dividieron en dos grupos de cuatro sujetos. Las ratas fueron expuestas a un régimen de privación de agua de 23 horas diarias. Después de cada sesión experimental tuvieron acceso libre a bebederos con agua durante 20 minutos en sus cajas habitación.
Aparatos
Se emplearon dos cámaras experimentales con 92 cm de largo por 45 cm de ancho y 33 cm de altura. Cada cámara contó con 3 dispositivos de estimulación, el primero ubicado a la mitad de la pared sur, el segundo a la mitad de la pared lateral y el tercero a la mitad de la pared norte. Cada dispositivo contó con una tecla translúcida de 2.5 cm de diámetro ubicada a 19 cm del piso, la cual podía iluminarse de color amarillo por medio de un foco de 28-V. Un dispensador que entregó de 0.01 cc de agua en cada activación se colocó debajo de cada tecla translucida a 5 cm del piso. El dispensador de agua se iluminó mediante un foco de luz amarrilla de 28-V cada vez que se entregó agua. Cada dispensador de agua contó con un sensor de respuestas de entrada de cabeza en el bebedero. La luz general de las cámaras fue proporcionada por un foco de 10W que fue pintado de color azul para reducir su iluminación. Ambas cámaras se encontraban dentro de un cubículo de aislamiento acústico, mismo que contaba con 4 ventiladores de 115W. La programación, registro y recolección de eventos se realizó mediante un equipo de cómputo que contó con una interfase y el software MED-PC IV para ambiente Windows. El análisis de datos se realizó a través de los software PASW Statistics 17 y SigmaPlot 11.0 (ver Figura 1).

Procedimiento
En este estudio las ratas de ambos grupos experimentales fueron expuestas a tres fases experimentales sin moldeamiento a bebedero y/o respuesta.
Las tres fases en que consistió el estudio se distinguieron por la presentación de agua durante 5 segundos en coordenadas espaciales fijas, secuenciales o variables, empleando un programa de Tiempo Fijo de 20 segundos (TF 20) para todas las condiciones. Para el Grupo 1, la Fase 1 consistió en la presentación del agua exclusivamente en el bebedero 1, mientras que en la Fase 2 el agua se presentó de manera secuencial en cada uno de los bebederos. En la Fase 3, el agua se presentó en un orden aleatorio en los distintos bebederos. Por su parte, el Grupo 2 se expuso a las siguientes condiciones: La primer fase fue idéntica a la Fase 1 del Grupo 1, mientras que en las fases 2 y 3 la presentación del agua fue aleatoria y secuencial, respectivamente. Las fases constaron de 15 sesiones experimentales cuya duración fue de 40 minutos cada una.

RESULTADOS
Poder identificar cuándo el comportamiento psicológico de un organismo se encuentra en cierto nivel de complejidad requiere de criterios. Como se mencionó anteriormente, para el caso de la función contextual se supone que el organismo es capaz de ajustarse diferencialmente a las contingencias presentes en su entorno, es decir, el organismo reproduce conductualmente alguna propiedad temporal y/o espacial con la que se presentan los eventos de estímulo. A partir esta lógica, es posible desarrollar unidades de medición específicas que permitan identificar el ajuste diferencial en una situación determinada, teniendo en cuenta que las preparaciones experimentales variaron las propiedades espaciales de los eventos de estímulo y por lo tanto, el organismo para lograr entrar en contacto con dicha estimulación debió modular su actividad según las propiedades espaciales de los eventos presentados. A continuación se describe las unidades de medida empleadas para estos propósitos:
1. Respuestas de muestreo, hace referencia a la frecuencia de respuestas de introducción de cabeza que se presentan fuera del periodo de entrega de agua.
2. Respuestas durante entrega de agua, hace referencia al número de respuestas de introducción de cabeza que se presentan durante el periodo de entrega de agua y en el bebedero en el que se entrega la misma.
3. Porcentaje de entregas de agua con al menos una respuesta. Ésta refiere el porcentaje de ocasiones durante una sesión en los que por lo menos el organismo introdujo una vez la cabeza al bebedero durante la entrega de agua.
A continuación se presentan los resultados del estudio en función de dichas medidas.
La Figura 2 muestra la frecuencia de respuestas durante la entrega de agua y la frecuencia de respuestas de muestreo (eje Y1), así como el porcentaje de entregas de agua con al menos una respuesta (eje Y2) para los sujetos del Grupo 1 en cada una de las fases experimentales. Los resultados obtenidos mostraron porcentajes por debajo del 60% de entregas de agua con por lo menos una respuesta a lo largo de las tres fases experimentales en cada una de las ratas. Durante la Fase 1, se observaron los porcentajes más altos de esta medida. Se observó una disminución a valores por debajo del 40% en las Fases 2 y 3. Por su parte, la frecuencia de respuestas durante los periodos de entrega de agua se mantuvo relativamente constante a lo largo de todo el experimento mostrando valores cercanos a las 50 respuestas por sesión en todas las ratas. Por último, la medida de frecuencia de respuesta de muestro fue la que mostró una mayor variación intra-condición y entre ratas. Durante la Fase 1, todas las ratas mostraron frecuencias de respuesta de muestreo cercanas al valor de 400 en las primeras sesiones, observándose un decremento irregular conforme transcurrió la fase, con excepción de la rata R1, quien mantuvo dichos valores a lo largo de la condición. Durante la Fase 2, las ratas R1 y R2 mostraron una frecuencia de respuesta de muestreo variable comprendida entre los valores de 100 a 400, mientras que las ratas R3 y R4 mostraron una ejecución por debajo de las 200 respuestas a lo largo de la fase.
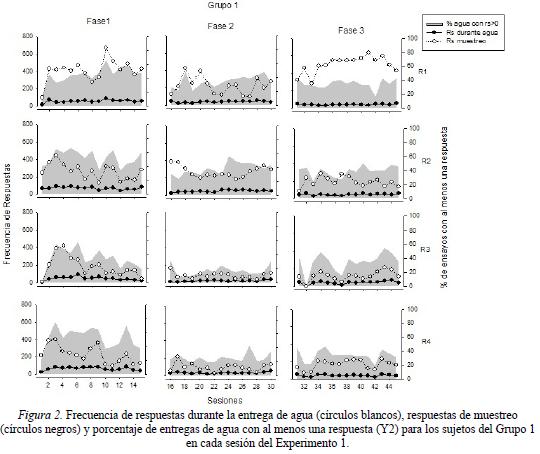
En la Fase 3, todas las ratas mostraron una frecuencia de respuestas de muestreo cercano al valor de 200, con excepción de la rata R1 que mostro una frecuencia de respuestas entre 400 y 600 a lo largo de esta fase.
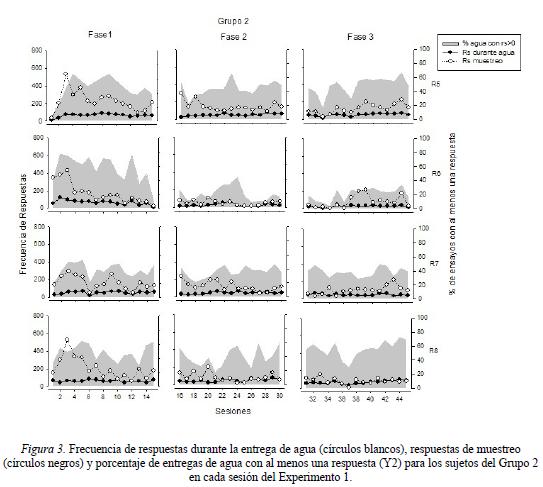
La Figura 3 muestra la frecuencia de respuestas durante la entrega de agua y la frecuencia de respuestas de muestreo (eje Y1), así como el porcentaje de entregas de agua con al menos una respuesta (eje Y2) para los sujetos del Grupo 2 en cada una de las fases experimentales. Los resultados en este grupo mostraron una variabilidad notoria en los porcentajes de entregas de agua con al menos una respuesta entre sujetos y entre condiciones, observándose en todos los casos valores por debajo del 80% a lo largo del estudio. Durante la Fase 1, las ratas R5 y R6 obtuvieron porcentajes comprendidos entre el 40 y el 80% de entregas con respuesta, mientras que mantuvieron un porcentaje comprendido en el rango entre el 40 y el 60% de esta medida. Durante la Fase 2, todas las ratas mostraron porcentajes de entregas con una respuesta comprendidos entre el 30 y el 60% de las entregas, con excepción de la rata R6, cuyos porcentajes oscilaron entre el 20 y 40%. Por su parte, en la Fase 3, se observó un desempeño variable en el porcentaje de entregas de agua con respuesta en las primeras diez sesiones, mostrando una mayor consistencia en la ejecución a partir de la segunda mitad de la condición experimental. Con excepción de la rata R6, que mostró porcentajes por debajo de el 30% a lo largo de la fase, el resto de las ratas (R5, R7 y R8) obtuvieron porcentajes entre el 40 y el 70% de entregas con al menos una respuesta.
Al igual que en el Grupo 1, la frecuencia de respuestas durante el periodo de entrega de agua se mantuvo relativamente constante a lo largo de todo el experimento mostrando valores alrededor de las 50 respuestas por sesión. Finalmente, con respecto a la frecuencia de respuestas de muestreo se observó una mayor variación en la ejecución durante la Fase 1 en todas las ratas, observándose una mayor frecuencia en las primeras sesiones (oscilando entre las 100 y 500 respuestas), para posteriormente disminuir su frecuencia a valores entre 50 y 100 respuestas en las últimas sesiones. Durante la Fase 2, se mantuvo la ejecución en todas las ratas en un rango de frecuencia entre las 50 y las 200 respuestas. Con excepción de la rata R5, la cual mostró una frecuencia relativamente constante de 200 respuestas de muestreo por sesión, el resto de las ratas mostró un decremento gradual a partir de la segunda mitad de la fase, llegando a valores cercanos a 50 respuestas de muestreo por sesión. Por último, en la Fase 3, se mostró un efecto contrario a la fase previa en la medida que en tres de las cuatro ratas (R5, R6 y R7) se observó un incremento en la frecuencia de respuestas de muestreo a partir de la sesión 37. La frecuencia observada en estas ratas fue de entre 50 y 200 respuestas de muestreo por sesión. La rata R8, se mantuvo con una frecuencia alrededor de las 50 respuestas de muestreo a lo largo de toda la fase experimental.
DISCUSIÓN
El Experimento 1 se realizó con el propósito de explorar el efecto de variar la ubicación espacial en la que se entrega el agua de manera no contingente sobre el ajuste diferencial en ratas. Los resultados obtenidos a partir de las medidas empleadas sugieren que éstas son sensibles para la detección de la modulación de la actividad de la rata bajo estas condiciones. En primer lugar, aunque todas las ratas fueron sometidas a condiciones de entrega de agua similar, el tipo de contacto que tuvo cada una de las ratas con la presentación del agua fue distinto. Esto se observa al analizar el porcentaje de entregas de agua con por lo menos una respuesta en conjunción con la frecuencia de respuesta durante la entrega. Los datos observados pudieran dar cuenta tanto de las diferencias entre sujetos como de la variabilidad intra sujeto observados en este estudio.
Por otro lado, los datos obtenidos respecto de la frecuencia de las respuestas de muestreo, parecieran sugerir que la presentación del agua en coordenadas espaciales fijas promueven un decremento gradual en esta medida conforme transcurren las sesiones, lo que pudiera indicar un efecto en la disposición espacial de la actividad de la rata en términos de su orientación hacia el dispositivo de entrega de agua. Este efecto pudo haberse generalizado a las condiciones de entrega espacial secuencial y aleatoria en la medida en que la frecuencia de respuestas de muestreo se mantuvo relativamente constante en ambos grupos y en ambas condiciones, con excepción de las ratas R1 y R2, quienes mostraron ejecuciones variables a lo largo de ambas condiciones. Suponemos que la diferencialidad se evidencia en el momento en el que el organismo reduce sus respuestas de introducción de cabeza en los periodos en los que no hay entrega de agua, y aumenta estas respuestas en los períodos en los que los bebederos se encuentran activos. Sin embargo, los datos de esta medida no permiten hacer esta aseveración ya que se mantuvieron en valores relativamente constantes a lo largo de todo el experimento.
La variación en el porcentaje de entregas de agua con por lo menos una respuesta de bebedero bajo las condiciones de cambio en espacio secuencial y aleatorio pudiera sugerir que las ratas no hicieron contacto con la contingencia programada. Sin embargo, este efecto también se obtuvo en la condición de espacio fijo para la entrega, aunque en menor medida ya que se observaron porcentajes mayores de esta medida bajo esta condición, lo que puede suponer que el cambio en las condiciones, de espacio fijo a espacio secuencial, afecta la manera en la que el organismo entra en contacto con los eventos programados ya que esta última contingencia implica el desplazamiento de la rata en el espacio experimental además de una re-orientación respecto del resto de dispositivos de entrega de agua. Es posible que la disminución en el porcentaje de entregas de agua con por lo menos una respuesta esté asociado a un efecto de primacía (Ribes & Torres, 2000) respecto de la orientación de la rata en espacio vinculada a el bebedero en el que se entregó en la Fase 1, y así afectar la diferencialidad mostrada durante dicha fase. Estos datos, aunque sin ser conclusivos, son congruentes con la propuesta de Ribes (1995), que supone que la promoción de contingencias de función depende de contingencias de ocurrencia consistentes en tiempo y espacio.
En el siguiente estudio, se pretende evaluar si el ajuste diferencial asociado a la entrega no contingente de agua se ve afectado con la introducción de una señal luminosa asociada a diferentes condiciones de presentación espacial del agua. Esta condición experimental se relaciona con el caso de asociación propuesto por Ribes y López (1985).
EXPERIMENTO 2
MÉTODO
Sujetos
Se emplearon 8 ratas hembras de la cepa Wistar de 3 meses de edad e ingenuas al inicio del experimento. Se dividieron en 2 grupos de 4 sujetos. Las ratas fueron expuestas a un régimen de privación de agua de 23 horas diarias. Después de cada sesión experimental tuvieron acceso libre a bebederos con agua durante 20 minutos en sus cajas habitación.
Aparatos
Los mismos que en el Experimento 1.
Procedimiento
Las tres fases de las que constó el diseño experimental se distinguieron por la presentación de una luz y de agua en posiciones espaciales fijas, secuenciales o variables, empleando un programa TF 20 para todas las condiciones. Específicamente, se presentó una luz por 5 segundos antes de cada entrega de agua. Al concluir la presentación de la luz se entregó agua por 5 segundos. Para el Grupo 1 la presentación de la luz fue en una posición espacial fija a través de las fases, esto quiere decir que la señal se presentó exclusivamente en el dispositivo 1. Durante la Fase 1, tanto la señal como el agua se presentaron en una posición espacial fija, solamente en el bebedero 1 a través de los ensayos, mientras que en la Fase 2 la presentación del agua fue en orden secuencial en cada uno de los bebederos, es decir, primero se entregó en el dispositivo 1, después en el dispositivo 2, posteriormente en el dispositivo 3, nuevamente en el dispositivo 1, y así sucesivamente. Finalmente, en la Fase 3 el agua se presentó en un orden aleatorio en los distintos bebederos. Para el Grupo 2 se presentaron tanto la señal como el agua en el mismo dispositivo a través de las fases, es decir, siempre fueron espacialmente contiguos. La Fase 1 para este grupo consistió en la presentación de los estímulos en una posición espacial fija, siendo esta fase idéntica a la Fase 1 del Grupo 1. La Fase 2 se caracterizó por la presentación de ambos estímulos en coordenadas espaciales secuenciales, primero se presentaron en el dispositivo 1, después en el 2 y así sucesivamente. La Fase 3 fue similar a la Fase 2 con la excepción de que el orden en el que se presentaron los estímulos en el mismo dispositivo fue aleatorio. Las fases constaron de 15 sesiones experimentales que tuvieron una duración de 40 minutos cada una.
RESULTADOS
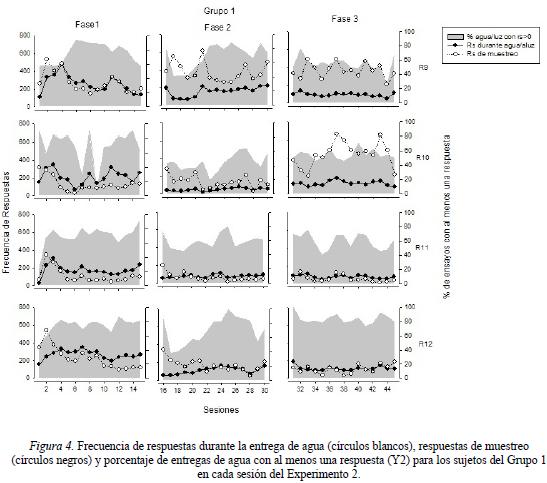
La Figura 4 muestra la frecuencia de respuestas durante la entrega de agua y la frecuencia de respuestas de muestreo (eje Y1), así como el porcentaje de entregas de agua con al menos una respuesta (eje Y2) para los sujetos del Grupo 1 en cada una de las fases experimentales. Los resultados obtenidos mostraron porcentajes de entregas de agua con por lo menos una respuesta variable en un rango entre 60% y 90% para todas las ratas durante la Fase 1, exceptuando la rata R10, que mostró valores cercanos al 20% durante las sesiones 7 y 9. En las primeras sesiones de la Fase 2 todas las ratas mostraron porcentajes alrededor de del 40%. A partir de las sesiones de la segunda mitad de la Fase 2, todas las ratas presentaron porcentajes cercanos al 80%, con excepción de la rata R10, que presentó valores cercanos a 60%. Durante la Fase 3, todas las ratas mostraron porcentajes alrededor del 60%, exceptuando a la rata R12, cuyos porcentajes mostrados se acercaron al 80%. Por su parte, la frecuencia de respuestas durante los periodos de entrega de agua durante las primeras sesiones mostró valores de 300 a 600 respuestas que fue disminuyendo conforme transcurrió la Fase 1, con excepción de la rata R12, que mantuvo su ejecución entre 200 y 300 respuestas por sesión a lo largo de toda la primer fase. En la Fase 2 se observó una frecuencia de respuestas durante la entrega de agua en un rango entre 50 y 100 respuestas para las ratas R10 y R11, mientras que para las ratas R9 y R12 se observó una ejecución creciente llegando a valores cercanos a las 200 respuestas a partir de la segunda mitad de la fase. Durante la Fase 3, esta medida se mantuvo relativamente constante, mostrando valores cercanos a las 100 respuestas por sesión en todas las ratas. Por último, la medida de frecuencia de respuesta de muestro fue la que mostró una mayor variación intra-condición y entre ratas. Durante la Fase 1, todas las ratas tuvieron frecuencias de respuesta de muestreo que varió en un rango de las 50 a las 500 respuestas por sesión en la primera parte de la fase y posteriormente llegaron a valores entre 200 y 300 en la segunda mitad de la fase. Durante la Fase 2, las rata R9 obtuvo una frecuencia de respuesta de muestreo variable comprendida entre los valores de 300 a 600, mientras que las demás ratas mostraron una ejecución que fue decrementando hasta valores por debajo de las 200 respuestas. En la fase 3, las ratas R 9 y R10 obtuvieron una frecuencia de respuestas de muestreo que varió en un rango entre las 200 y 600 respuestas, mientras que las ratas R11 y R12 mantuvieron una ejecución relativamente contante alrededor de las 100 respuestas a lo largo de toda la fase.
La Figura 5 muestra la frecuencia de respuestas durante la entrega de agua y la frecuencia de respuestas de muestreo (eje Y1), así como el porcentaje de entregas de agua con al menos una respuesta (eje Y2) para los sujetos del Grupo 2 en cada una de las fases experimentales. Los resultados obtenidos por las ratas R13 y R14, mostraron un porcentaje de entregas de agua con al menos una respuesta constante a lo largo de todo el estudio, observándose valores entre un rango del 60% y el 90%. Por su parte, la rata R16 mostró porcentajes de entregas de agua con al menos una respuesta en un rango entre el 20% y el 60% a lo largo del estudio Por último, la rata R15 mostró porcentajes bajos de esta medida (menores al 20%) a lo largo de las tres fases experimentales. La frecuencia de respuestas durante el periodo de entrega de agua mostró una ejecución relativamente constante intra sujeto a lo largo del estudio. Los resultados mostraron que la frecuencia de respuestas durante entrega de agua para las ratas R13 y R14 se mantuvo en un rango entre las 200 y 400 respuestas, mientras que esta frecuencia para la rata R16 se mantuvo en un rango entre las 100 y 200 respuestas durante todas las sesiones. La rata R15 mantuvo una frecuencia menor a las 30 respuestas a lo largo de todo el experimento. Finalmente, con respecto a la frecuencia de respuestas de muestreo se observó una mayor variación en la ejecución durante la Fase 1 para las ratas R13 y R14, observándose una mayor frecuencia en las primeras sesiones, oscilando entre las 200 y 400 respuestas, mientras que la rata R16 mantuvo una frecuencia de respuestas de muestreo cercano al valor de 100 a lo largo de la fase. La rata R15 obtuvo una frecuencia de respuestas menor a 30 a lo largo de todas las fases. Durante las Fases 2 y 3, las ratas R13, R14 y R16 mantuvieron una frecuencia de respuestas de muestreo en un rango de 50 a 200 respuestas.
DISCUSIÓN
El Experimento 2 se realizó con el propósito de explorar el efecto de variar la ubicación espacial en la que se presenta un estímulo contextualizador (agua no contingente) y un estímulo contextualizado (luz) sobre el ajuste diferencial en ratas. Al igual que para el Experimento 1, los resultados obtenidos a partir de las medidas empleadas sugieren que éstas son sensibles para la detección de la modulación de la actividad de la rata bajo condiciones de entrega no contingente de agua. Los resultados del presente experimento mostraron cierta variabilidad, entre sujeto y entre condiciones al igual que en el Experimento 1, sin embargo, los resultados mostrados por el Grupo 2 fueron relativamente consistentes.
En primer lugar, se observa que los datos obtenidos respecto de la frecuencia de las respuestas de muestreo, replican el efecto mostrado en la misma medida durante el Experimento 1, en particular, el decremento gradual de estas respuestas conforme transcurren las sesiones en las que la presentación del agua sea en coordenadas espaciales fijas. Por otro lado, las ratas R9 y R10 durante la Fase 3, mostraron un incremento en la frecuencia de respuestas de muestreo a diferencia del resto de los sujetos del estudio. Se podría sugerir, de manera no concluyente, que este efecto fue producido por la distancia entre la luz y la entrega de agua, que fue la manipulación que caracterizó a este grupo.
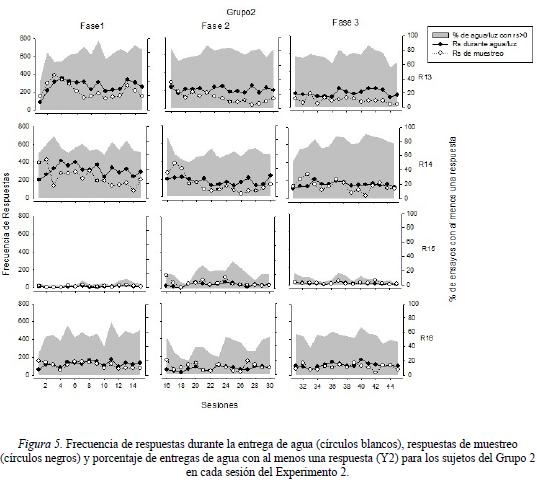
Con respecto al porcentaje de entregas de agua con al menos una respuesta, los resultados mostraron niveles relativamente constantes para todas las ratas durante las diferentes fases experimentales. Este resultado fue diferente al obtenido por el Grupo 1, en el que los porcentajes mostraron variabilidad, haciendo posible sugerir que dicha constancia en los porcentajes fue promovida por la presentación de la luz de forma espacialmente contigua a la entrega de agua.
DISCUSIÓN GENERAL
El propósito general de este trabajo fue explorar los efectos de variar la ubicación espacial en la entrega no contingente de agua bajo distintas condiciones de señalización en el ajuste diferencial en ratas, asociado a la Función Contextual propuesta por Ribes y López (1985). Dicho ajuste se caracteriza por el responder diferencial del organismo a la ocurrencia de eventos de estímulo. Desde esta perspectiva, la estructuración de esta función supone que el organismo es modulado conductualmente a partir de las propiedades espacio-temporales de los eventos de estímulo que se presentan en el entorno (Torres, Ortiz, Rangel & González, 2012).
En relación con lo anterior, el análisis de los resultados obtenidos nos permiten sugerir que los sujetos en ambos experimentos mostraron cierta sensibilidad a los elementos que estructuraron la situación experimental (ubicación y temporalidad en la entrega de agua y la señal luminosa). Las medidas de porcentaje de entrega de agua con por lo menos una respuesta y la de frecuencia de respuesta durante el periodo de entrega pudiéramos reconocerlas como medidas de contacto con la contingencia, mientras que funcionalmente la medida de frecuencia de respuesta de muestreo nos esté dando cuenta de segmentos de respuesta que le permiten al sujeto ajustarse a las propiedades temporales y espaciales de los eventos de estímulo, en este caso, considerando a la entrega de agua como evento Contextualizador (Ribes & López, 1985). Por lo tanto, estas medidas en conjunto podrían brindar evidencia de ajuste diferencial respecto de la estructura experimental propia de los estudios presentes.
De acuerdo con lo anterior, el resultado más consistente se aprecia en la variabilidad de la ejecución observada en el presente trabajo. El tipo de contingencia que se estructuró en ambos estudios supone la entrega de agua en intervalos regulares de manera independiente de la actividad de las ratas (TF 20s). En este sentido, la única respuesta requerida (Ribes-Iñesta & Montes, 2009) estuvo relacionada con el consumo de ésta. Esto implica que la actividad de la rata se vea modulada por la ubicación espacial del bebedero en que estaba disponible el agua. Los datos obtenidos en ambos estudios sugieren que las ratas no entraron en contacto con la entrega de agua en todos los ensayos, por lo que se observaron diferencias entre ratas y entre condiciones. Suponemos que estas diferencias se explican por el momento en el cada rata entró en contacto con la contingencia programada. Teniendo en cuenta que los resultados muestran diferencias entre ratas a pesar de que, por lo menos en la Fase 1, las condiciones experimentales fueron similares para ambos grupos de cada experimento, es posible sugerir que la ejecución de cada rata varió dependiendo del momento en el que éstas comenzaron a responder de manera consistente ante las ocurrencias de la señal y/o del agua (Ribes- Iñesta & Montes, 2009). De aceptarse lo anterior, se podría suponer que la posterior ejecución de cada una de las ratas se vio afectada por el momento en el que se entró en contacto con la contingencia programada.
A pesar de la variabilidad observada entre condiciones, un resultado que se muestra en la mayoría de las ratas de ambos estudios, es un porcentaje de entregas de agua con por lo menos una respuesta mayor en la Fase 1, en comparación con el resto de las fases. En términos de la propuesta realizada por Ribes-Iñesta y Montes (2009) en relación a las propiedades funcionales de las distintas formas de respuesta, una posible explicación para este resultado se basa en que al entregarse el agua en coordenadas espaciales fijas, la Respuesta Contingente incluye a la Respuesta Requerida, mientras que en las fases en las que se entregó el agua en coordenadas espaciales secuenciales o aleatorias, dichas respuestas se distinguen haciendo más complejo el ajuste. En el primer caso, la rata podría permanecer en un área cercana al único bebedero que entregó agua (Respuesta Compatible), consumiendo cada vez que éste se activara (Respuesta Contingente). Por otro lado, en el caso en el que se entregara agua en distintas ubicaciones espaciales, la rata se vería forzada a desplazarse de un bebedero a otro (Respuesta Requerida) para lograr consumir el agua.
Para ambos grupos del Estudio 2, exceptuando la rata R15, el resultado anterior es acompañado por una frecuencia alta de respuestas durante el periodo de entrega de agua. Continuando con la terminología empleada en el párrafo anterior, es posible sugerir que esta alta frecuencia es explicada por la presentación de una señal luminosa, la que afectaría el desplazamiento hacia el bebedero próximo por activarse (Respuesta Requerida) y favoreciendo el contacto con la entrega de agua. En este sentido, la señal se conceptualiza como un evento Contextualizado (Ribes & López, 1985) en la medida en que adquiere una relación funcional con el Estímulo Contextualizador.
Como mencionó anteriormente en el apartado de la discusión para el Estudio 2, la alta frecuencia en las respuestas durante el periodo de entrega de agua se muestra más consistente en las ratas R13 y R14 del Grupo 2. Este grupo se expuso a condiciones en las que la presentación de la señal y el agua fueron espaciotemporalmente contiguas. Partiendo de que la señal favorece el contacto con la entrega del agua al promover el desplazamiento hacia el bebedero próximo en activarse, es posible sugerir, que en las condiciones a las que se expusieron las ratas del Grupo 1, la señal facilitó exclusivamente el contacto con propiedades temporales de la entrega de agua. Esto en tanto que la señal no varió su ubicación espacial, y por lo tanto no fue contigua espacialmente a todas las entregas de agua. En este sentido, la señal indicaba el momento de entrega pero no su ubicación. Por otro lado, las condiciones a las que se expuso a las ratas del Grupo 2, se caracterizaron porque la señal varió de ubicación espacial en conjunto con la entrega de agua. Suponemos que a diferencia de lo ocurrido en el Grupo 1, esta contigüidad entre estímulo y lugar de entrega favoreció el contacto con propiedades, tanto temporales como espaciales de la ocurrencia de agua, lo cual favoreció la anticipación por parte de las ratas en el momento y lugar de ocurrencia.
Los resultados obtenidos en el presente trabajo pueden ser comparados con los resultados de estudios realizados bajo otros marcos teóricos. Para esto, es necesario señalar que la taxonomía de funciones estímulo-respuesta propuesta por Ribes y López (1985), dentro de la que se incluye a la Función Contextual, tuvo como contribución principal el intentar cubrir la clasificación y descripción exhaustivas de una gran diversidad de fenómenos psicológicos con una lógica jerárquica de inclusividad progresiva (Ribes, 2004). De esta manera, muchos de los datos obtenidos en estudios realizados desde diversas tradiciones teóricas en psicología pudieran ser recuperados y reinterpretados atendiendo a las categorías y criterios para la identificación de los distintos niveles de organización funcional de la conducta que se proponen en esta taxonomía.
En estudios en los que se ha empleado el paradigma del condicionamiento clásico o Pavloviano existen diversas investigaciones que han demostrado que la contigüidad espacial entre el estímulo condicional y el estímulo incondicional provocan una mayor magnitud de respuestas condicionales, mientras que la ausencia de contigüidad entre dichos estímulos promueven un efecto contrario sobre la respuesta condicional (Testa, 1975; Rescorla & Cunningham, 1979; Bruner & Landaverde, 1985; Cabrera & Vila, 1986).
A pesar de que este resultado no puede tomarse como equivalente a los obtenidos en el presente estudio, es posible compararlos si se considera que las preparaciones experimentales empleadas son situaciones genéricas ante las que el organismo se ajusta al entorno y por lo tanto los resultados podrían ser reinterpretados como evidencia del desarrollo de ajuste diferencial. Como se mencionó anteriormente, los resultados obtenidos por los ratas R13 y R14 parecen corroborar el supuesto de que la contigüidad espacial entre el estímulo condicional y el estímulo incondicional promueven una mayor magnitud en la respuesta condicional, suponiendo que ésta promueve mayor contacto con la entrega del estímulo incondicional.
Finalmente, se considera que estudios posteriores permitirán dar cobertura empírica a preguntas que se generaron a partir de este estudio y no fue posible responderlas por completo, así como permitirán explorar el efecto de variar la ubicación espacial de los eventos de estímulo que participan en formas de organización funcional del comportamiento más complejas incluidas en la formulación de Teoría de la Conducta propuesta por Ribes y López (1985).
REFERENCIAS
Bruner, C., & Landaverde, J. (1985). Los efectos de variar la ubicación espacial de un estímulo en la situación de automoldeamiento/automantenimiento. Revista Mexicana de Análisis de la Conducta, 11, 11-20. [ Links ]
Cabrera, R., & Vila, J. (1986). La localización espacial del estímulo condicionado determina la naturaleza de la respuesta condicionada. Revista Mexicana de Análisis de la Conducta, 12, 19-32. [ Links ]
Kantor, J. R. (1959/1978). Psicología Interconductual: un ejemplo de construcción científica sistemática. México: Trillas. [ Links ]
Rescorla, R. A., & Cunningham, C. L. (1979). Spatial contiguity facilitates pavlovian second-order conditioning. Journal of Experimental Psychology: Animal Behavior Processes, 5, 151-161, Available via: http://dx.doi.org/10.1037/0097-7403.5.2.152 [ Links ]
Ribes, E. (1995). Causalidad y contingencia. Revista Mexicana de Análisis de la Conducta, 21, 123-142. [ Links ]
Ribes, E. (2004). Acerca de las funciones psicológicas: Un post-scriptum. Acta Comportamentalia, 12, 117- 127. [ Links ]
Ribes, E (2007). Estados y límites del campo, medios de contacto y análisis molar del comportamiento: reflexiones teóricas. Acta Comportamentalia, 15, 229-259. [ Links ]
Ribes, E., & López, F. (1985). Teoría de la conducta: un análisis de campo y paramétrico. México: Trillas. [ Links ]
Ribes-Iñesta, E. & Montes, E. (2009). Interacción de la privación de agua y los estímulos correlacionados con la entrega de agua en programas de reforzamiento continuo y de intervalo. Revista Mexicana de Análisis de la Conducta, 35, 61-85. [ Links ]
Ribes-Iñesta, E. & Torres, C. (2000). The spatial distribution of behavior under varying frequencies of temporally scheduled water delivery. Journal of the Experimental Analysis of Behavior, 73, 2, 195-209. [ Links ]
Testa, T. L. (1975). Effects of similarity of location an temporal intensity pattern of conditioned and unconditioned stimuli on the acquisition of conditioned suppression in rats. Journal of Experimental Psychology, 104, 114-121. [ Links ]
Torres, C., Ortiz, G., Rangel, N., & González, V. (2012). Análisis del concepto situación en la estructuración de las funciones psicológicas. En Padilla, M. & Pérez-Almonacid, R. (Ed.) La función sustitutiva referencial: análisis histórico-crítico / avances y perspectivas (pp.149-167). New Orleans, LA: University Press of the South. [ Links ]
Received: May 25, 2012
Accepted: February 11, 2013
1 La presente investigación fue posible gracias a la beca #45231 otorgada por el Consejo Nacional de Ciencia y Tecnología al primer autor para cursar la Maestría en Ciencia del Comportamiento. Enviar correspondencia a Felipe Patrón, Centro de Estudios e Investigaciones en Comportamiento, Francisco de Quevedo #180 (Arcos Vallarta), Guadalajara, Jal. 44100, correo electrónico: patron6d5@hotmail.com
