











 (pdf)
(pdf)




 Permalink
PermalinkRevista de Etologia
ISSN 1517-2805 ISSN 2175-3636
Rev. etol. v.4 n.2 São Paulo dez. 2002
ARTIGOS
O comportamento interespecífico de defesa do quero-quero, Vanellus chilensis (Molina, 1782) (Charadriiformes, Charadriidae)*
Interspecific aggressive behavior in the southern lapwings, Vanellus chilensis (Molina, 1782) (Charadriiformes, Charadriidae)
Leny Cristina Milléo Costa
Pontifícia Universidade Católica do Paraná
RESUMO
Com o objetivo de analisar seu comportamento numa situação de conflito interespecífico, em resposta a intrusos e predadores, quero-queros Vanellus chilensis foram observados na área urbana de Curitiba, na área rural da Fazenda Experimental da Cargill, em Toledo, e no cativeiro do Passeio Público em Curitiba. Registrou-se o número de intrusos, o número de quero-queros que atacam e perseguem, a freqüência dos padrões motores diante das invasões direcionadas e não direcionadas e das perseguições e tipos de ataques. Estratégias comportamentais como manobras de distração, escape, ataque, ameaças, foram observadas dependendo do período anual. No período não reprodutivo, os quero-queros responderam com posturas de fuga, andando, correndo e voando. No período reprodutivo, as respostas mais freqüentes foram manobras de distração durante a incubação dos ovos e a proteção dos filhotes. As respostas defensivas foram relacionadas à distância e ao modo de aproximação dos intrusos.
Descritores: Comportamento defensivo, Conflitos interespecíficos, Quero-queros, Vanellus chilensis.
ABSTRACT
Interspecific conflict responses of Southern Lapwings Vanellus chilensis to intruders and predators were observed in the urban area of Curitiba, in the Cargill experimental farm in Toledo, and in the Passeio Público Park of Curitiba, Brazil in captivity. Number of intruders, number of lapwings attacking and pursuing, frequency of response to directed and undirected invasions, of persecutions and types of attacks were recorded. The use by lapwings of distraction displays, escape, attack, and threat behavior varied according to season. In the non-reproductive season, lapwings escaped from intruders by walking, running and flying. In the reproductive season, the most frequent responses were distraction displays, performed during the incubation of eggs and the protection of nestling. Defensive responses depended on the distance and the type of approach of intruders.
Keywords: Defensive behavior, Interspecific conflict, Southern lapwings, Vanellus chilensis.
O quero-quero, Vanellus chilensis, ocorre da América Central até a Terra do Fogo e em todo o Brasil (Sick, 2001). É uma ave originalmente encontrada em áreas de campo com terrenos lodosos ou às margens dos rios e dos lagos (Walters, 1980). Trata-se de uma ave gregária cujo grupo varia em tamanho de acordo com o período anual (Costa, 1985). No período não reprodutivo, os grupos variam de quatro a oitenta aves (Costa, 1999); no período reprodutivo, os quero-queros apresentam a monogamia e a poligamia como sistemas de acasalamento e como forma de agrupamento (Walters, 1980, 1990; Walters & Walters, 1980).
A presença do trio reprodutivo foi registrada entre outras espécies de Vanellus. Berg (1993) notou que Vanellus vanellus se reproduzem em agregações de semi-colônias e, mesmo sendo monogâmicos, podem, algumas vezes, tornar-se poligâmicos em áreas de alta densidade populacional. Vanellus tricolor, Vanellus miles (Dann, 1981) e Vanellus crassirostris, Vanellus armatus (Walters, 1980), Vanellus lugubris, Vanellus melanopterus (Ward, 1989) são monogâmicos. Vanellus coronatus é monogâmico (Walters, 1980) e poligâmico com duas fêmeas e um macho permanecendo com eles e com os filhotes (Ward, 1989).
O quero-quero interage com muitos outros animais, em especial com as aves joão-de-barro, chupim e sabiá-laranjeira. Dentre os mamíferos, destacam-se ovelhas, cães, gatos, cavalos e vacas. Essa convivência está associada ao fato dos quero-queros habitarem ambientes rurais e urbanos. É comum observar-se Vanellus sp. com outras espécies como Pluvialis apricaria e Larus ridibundus. (Barnard & Stephens, 1981; Thompson & Barnard, 1983, 1984). Nas áreas urbanas, os Vanellus chilensis estão constantemente submetidos a influências externas e à circulação de um elevado número de pessoas, carros e máquinas.
O ambiente ocupado pelos quero-queros deixa-os vulneráveis à predação, desenvolvendo-se neles habilidades que os levam a detectar e reagir aos predadores. Estas estratégias defensivas foram observadas em Vanellus coronatus, Vanellus melanopterus e Vanellus lugubris (Ward, 1989), Vanellus armatus (Thomas, 1983), Vanellus crassirostris (Walters, 1979), Vanellus indicus (Kalsi & Khera, 1987) e Vanellus spinosus (Walters, 1990). O tipo de resposta utilizado por Vanellus vanellus é determinado pelo perigo que o predador representa (Elliot, 1985a).
As respostas defensivas de Vanellus chilensis são mais freqüentes no período reprodutivo, durante a proteção dos ovos e dos filhotes, manifestando-se então por perseguições e ataques aos predadores (Costa, 1985, 1994b; Myers, 1978, Oniki, 1986; Walters, 1982, 1980, 1990; Zuberbhühler, 1973). Entre os principais predadores naturais de Vanellus chilensis estão o gavião, Caracara plancus, o urubu, Coragyps atratus, a coruja, Speotyto cunicularia. No ambiente urbano, a presença humana tornou-se uma ameaça para os quero-queros, através da predação dos ovos, perseguições e através dos ataques a adultos e filhotes.
Os comportamentos de defesa de Vanellus chilensis são complexos: o repertório de agressão interespecífica difere do repertório intraespecífico. As únicas categorias comportamentais presentes em ambas os casos são as posturas de alerta e de advertência, descritas por Costa (1985).
Entre os quero-queros, o comportamento interespecífico foi mais estudado em Vanellus vanellus (Barnard & Stephens, 1981; Beintema & Visser, 1989a, 1989b; Buitron, 1983; Elliot, 1985a, 1985b; Galbraith, 1989; Grisser, 1990; Hesp & Barnard, 1989; Iversen, 1986; Kirby & Green, 1991; Mcmahon, 1996; Thompson, 1983, 1986; Thompson & Barnard, 1983, 1984; Thompson & Lendren, 1985), em Vanellus coronatus, Vanellus melanopterus, Vanellus lugubris (Ward, 1989), em Vanellus armatus (Thomas, 1983), Vanellus crassirostris (Walters, 1979) e Vanellus indicus (Kalsi & Khera, 1987).
Os registros em áreas urbanas mostram que a ocupação de espaço pelos quero-queros se dá em parques, praças, campus universitários, campos de futebol, aeroportos, nos telhados de casas e nos topos dos edifícios. Poucos estudos comportamentais foram no entanto realizados nas áreas antrópicas (Costa, 1985, 1999; Oniki, 1986; Ruszczyk et al., 1987).
Na tentativa de se conhecer um pouco mais a respeito da ocupação pelos quero-queros de áreas urbanas, buscou-se na presente pesquisa observar como as aves reagem às invasões e perseguições nos períodos reprodutivo e não reprodutivo. Levando-se em conta que os quero-queros também são encontrados em ambientes rurais e no cativeiro, buscou-se coletar informações sobre o comportamento interespecífico exibido pelos quero-queros nestes locais, para obter conhecimentos relevantes a respeito da organização social da espécie. Teve-se por objetivo específico observar o número de quero-queros que atacam e perseguem; o número de intrusos; a freqüência das categorias comportamentais diante das invasões direcionadas e não direcionadas e das perseguições, a freqüência das respostas dos quero-queros diante da distância dos intrusos nos conflitos dos períodos reprodutivo e não reprodutivo, assim como registrar os sinais de alarme dos quero-queros e analisar os tipo de ataques que exibem.
Método
As observações foram realizadas entre março de 1995 a janeiro de 1997, num total de 4500 horas de observação, no Parque Barigüi, no Jardim Botânico, no Campus da PUC PR, caracterizando-se estas como áreas urbanas do Município de Curitiba, Estado do Paraná. No ambiente rural, realizaram-se 90 horas de observações na Fazenda Experimental da Cargill, em Toledo, oeste do Estado do Paraná. No cativeiro foram efetuadas 40 horas de observações com dois quero-queros, no período não reprodutivo, no Passeio Público da cidade de Curitiba, num recinto de cativeiro de 35 x 12 x 6 metros.
Os registros das observações foram elaborados conforme Lehner (1996). Em cada confronto agonístico, foram anotados: o número e a espécie invasora, a distância do intruso aos adultos, ao ninho e aos filhotes (zero a cinco metros, cinco a 10 metros, 10 a 15 metros acima de 20 metros), o modo de aproximação do intruso e o número de quero-queros que participavam da defesa com respostas desencadeadas.
Considerou-se intruso qualquer indivíduo de outra espécie que se deslocasse na área ocupada pelos quero-queros, interagindo ou não com estes. Registrou-se como invasões direcionadas as aproximações dos intrusos feitas na direção do adulto ou do ninho e como invasões não direcionadas os casos em que o intruso apenas transitasse na área ocupada pelos quero-queros. Procurou-se, também, verificar se o invasor (presença humana) andava ou corria e ainda se as perseguições (aproximações com insistência) ocorriam em direção às aves adultas.
O método focal (Altmann, 1974) foi usado, sendo observados todos os quero-queros envolvidos desde o início do episódio, com a presença do intruso, até o final, com a expulsão dos intrusos ou a fuga dos quero-queros do confronto, obtendo-se assim a freqüência das categorias desencadeadas no comportamento de defesa. Utilizou-se de filmes e fotografias para a identificação e análise das categorias.
O comportamento defensivo dos quero-queros adultos relativamente aos filhotes somente foi observado no ambiente urbano. As invasões territoriais foram analisadas em cada uma das etapas do período reprodutivo (formação do grupo reprodutivo, corte, elaboração do ninho, nidificação e cuidado parental). Durante a nidificação, registrou-se as respostas executadas pelos quero-queros no momento de uma invasão pelo solo e de acordo com a distância do intruso ao ninho. Verificou-se o comportamento da ave que estava nidificando diante de uma invasão territorial .
Na elaboração do ninho e na corte, as posturas foram registradas sem que fosse diferenciado o sexo dos quero-queros envolvidos na atividade. Os quero-queros que formam o trio reprodutivo de nidificação foram chamados de aves "A", "B", e "C", uma vez que há ausência de dimorfismo sexual externo nesta espécie, e uma vez que não havia observações relacionadas com a posição de cópula. Como o método foi estritamente observacional, não envolvendo marcação individual, os quero-queros foram diferenciados pelos comportamentos que apresentavam no momento do registro. O quero-quero "A" era a ave que se encontrava nidificando ou próxima do ninho; o quero-quero "B" era o que se encontrava nos limites territoriais e que revezava o outro na incubação dos ovos. Considerou-se como a ave "C" o quero-quero que se deslocava nos limites territoriais sem participar da incubação dos ovos.
Foi realizada uma análise comparativa entre os ambientes urbano, rural e cativeiro; entre os comportamentos das aves da dupla e do trio reprodutivos; entre os modos de aproximação do intruso (direcionado, não direcionado e perseguição) e entre as categorias desencadeadas nas respostas agonísticas. Para tal foi utilizado o teste de c2, sendo aplicada a correção de Yates quando a freqüência esperada em uma das classes era menor do que cinco (os valores de c2 são, neste caso, acompanhados de um asterisco).
Resultados
Os comportamentos defensivos interespecíficos de Vanellus chilensis ocorreram durante o ano todo e eram compostos por categorias de fuga, de ameaça, de perseguição, de manobras de distração, de ataque e de alerta. O quero-quero, ao realizar a fuga, utiliza-se das posturas de fuga, de agachamento e de deslocamento por vôo. A postura de fuga foi realizada pelo quero-quero andando e/ou correndo, posicionando as asas junto ao corpo e a cabeça e o pescoço estirados para frente; é uma categoria específica do período não reprodutivo numa defesa interespecífica. O agachamento é realizado pelos quero-queros por flexão das pernas, a parte ventral do corpo encostando-se no solo, a ave permanecendo imóvel diante do intruso. (Figura 1).
Fuga

Ameaça
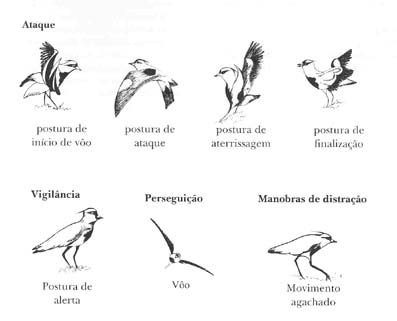
Figura 1. Comportamento de fuga e ameaça do quero-quero.
Do comportamento de ameaça fazem parte as posturas de ameaça frontal, de ameaça 1, de ameaça 2 e postura desencadeadora de ataque exclusivas do período reprodutivo (Costa, 1994a). A ameaça frontal é uma categoria executada estando o quero-quero de frente ao intruso, posicionado o corpo e a cabeça para frente. A postura de ameaça 1 foi executada em deslocamento rápido com o pescoço estirado para cima, corpo ligeiramente levantado e a cabeça com movimentos laterais. Na postura de ameaça 2 o quero-quero permanece estático, ameaçando o intruso com o corpo reto, cabeça ligeiramente estirada para frente e para baixo, com ângulo acentuado entre o corpo e a cabeça. A postura preparatória de ataque ocorre na seqüência da postura de ameaça 2, quando o quero-quero vai gradativamente encostando o corpo no solo, flexionando as pernas; preparando-se para o comportamento de ataque. (Figura 1).
Ataque

Figura 2. Comportamentos de ataque, vigilância,perseguição e manobras de distração do quero-quero.
No comportamento de perseguição, o quero-quero utiliza o vôo. Cabe destacar que há o vôo alto, baixo e rasante, presente também nos comportamentos de ataque, perseguição e de fuga (Figuras 1 e 2).
Entre as manobras de distração estão o movimento agachado e o agachamento-distração, manobras exclusivas do período reprodutivo durante o revezamento dos quero-queros na nidificação, assim como na defesa interespecífica (Costa, 1985, 1994b) (Figura 2).
O ataque é realizado em vôo através das posturas de início de vôo que antecedem o ataque, e das posturas de aterrissagem e de finalização que seguem o vôo rasante (Figura 2).
A atividade de vigilância com a postura de alerta apresentou-se estando os quero-queros parados e em deslocamento (Figura 2).
O desencadeamento das categorias comportamentais ocorreu de acordo com os períodos reprodutivo e não reprodutivo, número e espécie de invasor, modo de invasão assim como o tipo de aproximação do intruso.
Comportamento defensivo no período não-reprodutivo
Além do ser humano, os invasores da área territorial dos quero-queros foram ovelhas, cães, cavalos e vacas. Observou-se o carcará e a coruja buraqueira como predadores naturais e algumas espécies de pássaros como o sabiá-laranjeira, o joão-de-barro e o chupim que habitam e se deslocam no espaço territorial dos quero-queros.
As formas de aproximação dos intrusos (direcionada, não direcionada e perseguição) tiveram freqüências diferentes nos ambientes urbano (c2(2) = 67,42, p < 0,01), rural (c2(2) = 35,38, p < 0,01) e de cativeiro (c2(2) = 6,34, p < 0,01). Prevaleceram a perseguição na área urbana e na área rural e o direcionamento no cativeiro.
Os quero-queros, no período não-reprodutivo, utilizaram como forma defensiva os comportamentos de ameaça e de fuga e os comportamentos associados (alerta e fuga, manobras e alerta, alerta e agachado). O comportamento de fuga (n = 90, 64,4%) com a postura de fuga (n = 109, 44,9%) foi o mais freqüente em resposta a todas as formas de aproximação dos intrusos, em todos os ambientes. O comportamento de alerta ocorreu isolado (n = 90, 7,8%) ou associado ao comportamento de fuga (n = 90, 13,3%). O agachamento não foi a primeira resposta apresentada e teve baixa freqüência (n = 109, 8,2 %).
Houve freqüências similares das posturas de fuga e vôo-fuga nas várias formas de aproximação de intrusos. A postura de alerta, contudo, foi mais freqüentemente exibida nas aproximações não direcionadas (c2*(1) = 3,6, p < 0,05) e nas perseguições (c2(1) = 12,8, p < 0,01) do que nas aproximações direcionadas. As freqüências das posturas de alerta, fuga e vôo-fuga não diferiram entre ambientes.
Os quero-queros, quando se deparavam com um intruso, apresentavam uma categoria defensiva acompanhada ou não de vocalização. Não houve diferenças significativas nas freqüências das respostas acompanhadas ou não de gritos, nos três ambientes. Não ocorreram no cativeiro respostas vocalizadas e, no ambiente rural, respostas com grito foram mais freqüentes do que no ambiente urbano (c2*(1) = 25,1, p < 0,01).
Foi mais freqüente a emissão de som, na primeira resposta dos quero-queros a uma invasão, no caso das aproximações não direcionadas. No caso da perseguição, predominaram as respostas sem vocalização (c2(1) = 4,48, p < 0,05). Não houve diferença na freqüência das primeiras respostas sem vocalização
entre cativeiro e ambiente urbano. As categorias mais freqüentes, no ambiente urbano, foram as de fuga (c2(1) = 39,89, p < 0,01) e de alerta (c2(1) = 6,76, p < 0,01); no cativeiro, a categoria de fuga (c2(1) = 24,06, p < 0,01).
Comparando-se as respostas de acordo com a distância do invasor, verificou-se que a postura de agachamento-fuga foi mais freqüente na distância de 15-20 m do que na de 0-5 m (c2*(1) = 16,1, p < 0,01) e a postura de alerta foi mais freqüente a distâncias de 6-10 m do que a distâncias de 0-5 m (c2(1) = 3,5, p < 0,01). Estando o invasor a uma distância de 0-5 m, os quero-queros responderam com freqüência similar de fuga, vôo-fuga, alerta e agachamento, nos ambientes urbano e rural.
Durante o período não reprodutivo, as defesas com ataque, realizadas pelo indivíduo ou pelo grupo, foram raras (16 casos em 228 confrontos observados). Seu alvo foram cães e aves que atravessavam a área ocupada pelos quero-queros.
Comportamento defensivo no período reprodutivo
Ritual de acasalamento
O ritual de acasalamento compreende a construção do ninho, a cópula e a postura dos ovos. Nesta fase, a maioria dos quero-queros já tem uma área territorial, permanecendo nela em grupos de dois ou três (dupla ou trio reprodutivo, Costa, 1999). Na área rural foram observadas duplas reprodutivas e, na urbana, duplas e trios reprodutivos.
A aproximação não direcionada de intrusos, durante o período reprodutivo, predominou no caso das duplas rurais (c2(1) = 15,1, p < 0,01), e a perseguição, nas duplas urbanas (c2(1) = 17,12, p < 0,01). A aproximação direcionada foi mais freqüente nos trios rurais (c2(1) = 4,54, p < 0,05), mas a aproximação não direcionada e a perseguição não diferiram em freqüência, em comparação com as duplas rurais.
A primeira resposta dos quero-queros, frente a uma invasão, foi, na maioria das vezes, executada sem vocalização (85 ocorrências em 110 respostas). Respostas acompanhadas de vocalização foram mais freqüentes em duplas rurais do que em duplas urbanas (c2*(1) = 72,25, p < 0,01). Nos trios urbanos, as respostas acompanhadas de vocalização ocorreram com maior freqüência (c2*(1) = 69,4, p < 0,01).
A freqüência das posturas desempenhadas pelas duplas urbanas não diferiu das executadas pelas duplas rurais. Nos trios urbanos, o movimento agachado foi a postura menos freqüente (c2*(1) = 5,4, p < 0,01) e a de advertência a mais freqüente (c2*(1) = 11,5, p < 0,01). A postura de alerta foi vista apenas nos trios reprodutivos.
Nas duplas urbanas, diferiram as freqüências das categorias (c2 (4) = 14,8, p < 0,05): a postura de movimento agachado foi a mais freqüente (c2*(1) = 6, p < 0,01), não tendo sido observada a postura de alerta. Não diferiram em freqüência as posturas de fuga, agachamento-incubação, advertência e agachamento-distração.
Nos trios urbanos, também foi encontrada uma diferença significativa na freqüência das posturas (c2(4) = 16,9, p < 0,05), com predomínio da postura de fuga (c2*(1) = 11,1, p < 0,01), não tendo sido observada a postura de agachamento-distração.
Nas duplas rurais, não foi encontrada diferença significativa na freqüência das categorias comportamentais (c2*(3) = 6,76, p >0,05).
Incubação dos ovos
Durante o período de nidificação, os quero-queros defenderam a região do ninho. Nesta etapa do período reprodutivo, a primeira resposta emitida por duplas de quero-queros urbanos, diante de uma invasão, foi o ataque em vôo (21 vezes em 83 ocorrências) e a postura de movimento agachado (16 vezes em 83 ocorrências). Os trios responderam com o movimento agachado (52 vezes em 182 ocorrênci
as), alerta (42 vezes em 182 ocorrências) e ataque em vôo (33 vezes em 182 ocorrências).
As posturas de alerta (c2(1) = 67,3, p < 0,01), de movimento agachado (c2(1) = 9,6, p < 0,01) e de fuga (c2(1) = 6,6, p < 0,01) foram mais freqüentes nos trios reprodutivos do que nas duplas. As posturas de advertência (c2(1) = 11,4, p < 0,01), agachamento no ninho (c2(1) = 5,8, p < 0,01) e ameaça frontal (c2(1) = 8,4, p < 0,01) foram menos freqüentes. Não houve diferenças significativas na freqüência de aparecimento das demais posturas: vôo-ataque, alerta, agachamento-distração, fuga e deslocamento por vôo.
Os quero-queros em duplas responderam inicialmente com vocalizações, durante as invasões. Não diferiam em freqüência as posturas de alerta, agachamento-distração, advertência, fuga, e agachamento-ninho, havendo predomínio da postura de fuga (c2(1) = 10,7 p < 0,01). As posturas de vôo-ataque, vôo-deslocamento e movimento agachado ocorreram sem produção de vocalizações.
Não diferiram em freqüência as categorias comportamentais dos trios, acompanhadas de vocalização (movimento agachado, advertência e fuga). Agachamento-distração (c2(1) = 7,8, p < 0,01) e o vôo-deslocamento (c2(1) = 36,3, p < 0,01) foram as mais freqüentes, enquanto que a postura de alerta foi pouco freqüente e o vôo-ataque não ocorreu.
As duplas do ambiente rural que estavam no ninho durante uma invasão não diferiram na freqüência de exibição das posturas de alerta, movimento agachado, agachamento-ninho e vôo-deslocamento. Os trios urbanos que estavam no ninho não tiveram diferenças entre as posturas de alerta, movimento agachado e agachamento no ninho. O deslocamento por vôo foi pouco freqüente e as posturas de alerta, movimento agachado, agachamento no ninho e deslocamento por vôo não diferiram nos quero-queros A das duplas em comparação com os quero-queros A dos trios urbanos. A ave A de uma dupla permanecia no ninho quando o parceiro realizava o ataque como primeira resposta.
Quando invasões foram direcionadas ao ninho, a ave A respondeu com agachamento-manobra, alerta ou movimento agachado (11 vezes em 75 observações), a ave B executou o alerta (12 vezes em 48 observações) ou o ataque em vôo (10 vezes em 48 observações); e a ave C realizou ataque em vôo (12 vezes em 40 observações) ou a advertência (7 vezes em 40 observações).
Quando as invasões não foram direcionadas ao ninho, a ave A realizou o movimento agachado (15 vezes), o alerta (14 vezes) ou permanecia no ninho (13 vezes) em 40 observações; a ave C exibiu o alerta (12 vezes) e o ataque em vôo (9 vezes) em 56 ocorrências. O quero-quero B fez manobras de distração, participou de revezamento no ninho e realizou poucos ataques. O comportamento de ataque foi realizado pela ave C que não participavas da incubação dos ovos e permanecia sempre nas áreas mais afastadas do território.
Quando perseguidos, os quero-queros escaparam através do movimento agachado. Independentemente da distância do intruso, não houve diferença significativa na freqüência das posturas das aves B e C _ comparadas às da ave A. Cabe ressaltar que apenas as aves C executaram ameaça frontal a um intruso distante de apenas cinco metros. O ataque foi realizado pelos três quero-queros com um intruso situado até 10 metros de distância; acima desta distância não realizaram mais ataque em vôo. O movimento agachado foi observado nos quero-queros A e B.
Os quero-queros da dupla reprodutiva responderam com as seguintes categorias comportamentais: no caso das invasões direcionadas ao ninho, a ave A realizou advertência (9 vezes), alerta e ataque em vôo (6 vezes) e as manobras de distração (5 vezes) em 45 observações. Sua permanência no ninho ocorreu quando a ave B atacou o intruso. A ave B respondeu com ataques por vôo (22 vezes) além de realizar a postura de advertência (10 vezes) em 68 observações.
No caso das invasões não direcionadas ao ninho, a ave A respondeu com alerta e movimento agachado (4 vezes em 15 observações),
a ave B com ameaça frontal (7 vezes) e ataque em vôo e movimento agachado (4 vezes) em 29 observações. Neste tipo de invasão, o ataque foi um comportamento exclusivo da ave B.
Quando perseguidos, as aves A (n = 34) responderam com posturas de movimento agachado (12 vezes) e agachamento-distração (7 vezes). As aves B (n = 52) fugiram em movimento-agachado (12 vezes), advertência (8 vezes) e agachamento-distração (6 vezes).
Cuidado parental
No cuidado parental, não houve diferença na freqüência da postura de movimento-agachado entre as aves A e B, na defesa dos filhotes, independentemente da idade destes. A freqüência do agachamento-distração nas aves B foi mais freqüente do que nas aves A (c2(1) = 16,3, p < 0,01). Os quero-queros B adaptaram a postura de agachamento-distração durante o cuidado ao filhote recém-eclodido. A postura de advertência não foi observada nas aves A, durante o cuidado ao filhote recém-eclodido, nem nas aves B, durante a defesa dos filhotes de duas semanas. A postura de ameaça frontal somente foi observada no cuidado ao filhote recém-eclodido pelas aves A e B. O ataque em vôo ocorreu como defesa aos filhotes até a segunda semana e a postura de ameaça frontal desempenhada na proteção dos filhotes de uma semana.
Comportamento de ataque
A freqüência de ataques efetuados por duplas de quero-queros, mas não a de ataques efetuados por trios, foi inferior à freqüência dos ataques individuais (c2(2) = 154,5, p < 0,01). A freqüência dos ataques de trios não diferiu significativamente da freqüência de ataques de duplas. O número de intrusos atacados por quero-queros variou (c2(4) = 8,6, p < 0,01), sendo o ataque a um só mais comum e menos freqüentes os dirigidos a dois, três, quatro e cinco intrusos. O número de intrusos atacados diferiu, nos ataques de duplas de quero-queros (c2(4) = 15,7, p < 0,01), sendo o ataque a um intruso o mais freqüente e a três o menos freqüente. Os ataques dos trios reprodutivos foram geralmente direcionados a um intruso, não sendo observados ataques a mais de três intrusos.
Durante o período reprodutivo, os quero-queros atacaram pessoas, aves, predadores e cães. Os ataques realizados pelas duplas reprodutivas não diferiram dos realizados individualmente. Os cães não foram atacados por duplas ou trios de aves. A freqüência dos ataques de trios diferiu da dos ataques individuais (c2(2) = 267,1, p < 0,01). Nos ataques individuais, os quero-queros não atacaram igualmente aves, pessoas, predadores e cães (c2(3) = 165,23, p < 0,01). O número de pessoas e de aves atacadas foi mais freqüente o de predadores e cães, menos freqüente.
Discussão
No período não reprodutivo, os quero-queros apresentaram agrupamentos que variavam de quatro a oitenta aves, sendo o comportamento defensivo exclusivamente individual, em contraste com o período reprodutivo em que, além do comportamento de defesa individual e territorial, houve proteção aos ovos e aos filhotes.
Ao se defenderem, no período não reprodutivo, os quero-queros utilizaram-se basicamente dos comportamentos de fuga e de alerta, independentemente do modo de aproximação e da distância do intruso, tanto no ambiente urbano como no ambiente rural. Através da fuga, distanciavam-se do intruso e do local que ocupavam antes da invasão. A baixa freqüência do grito de alarme demonstrou que, no período não reprodutivo, a resposta era individual, servindo como autodefesa, sem implicação coletiva. Os demais integrantes do grupo escapavam quando, por sua vez, eram ameaçados pelo intruso. Esse aspecto de reação individual foi bem evidenciado nas aproximações de cães e de pessoas: os quero-queros escapavam à medida que eram ameaçados. Gritos eram emitidos durante a fuga por vôo, que se seguia à fuga pelo solo.
As aproximações não direcionadas nos ambientes urbano e rural, embora menos freqüentes que as perseguições, ocorreram em função da alta freqüência de fatores externos como carros, máquinas, pessoas e animais [cães, ovelhas, patos e gansos]. O comportamento de alerta foi exibido mesmo diante de aproximações não direcionadas. Sabe-se que convivência em grupo reduz o risco de predação e o tempo de vigilância, aumentando a disponibilidade de cada membro do grupo para forragear (Caraco, Martindale, & Pulliam, 1980).
Walters e Walters (1980) observaram que o comportamento antipredador de Vanellus chilensis, no período não reprodutivo, está relacionado à freqüência de ataques de carcarás e às ameaças com pedras do ser humano em relação às aves adultas. Os indivíduos de Vanellus chilensis observados no ambiente urbano não foram contudo atacados, mas perseguidos pelo ser humano, enquanto o ataque de predadores ocorreu em baixa freqüência.
O agachamento, embora seja uma resposta de fuga para Charadriiformes (Grisser, 1990; Simmons, 1955) e embora observado em filhotes dos quero-queros (Costa, 1985, 1999), foi uma conduta pouco freqüente nos adultos, no período não reprodutivo N = 109 (8,2%). É possível que represente uma atividade típica do estágio imaturo desencadeada nas primeiras semanas de vida e gradativamente substituída pelas posturas de fuga (andando, correndo) e de vôo. Simmons (1952) afirma que nos Charadriiformes o agachamento é um sinal de que a ave está alarmada: ela suspende as outras atividades, enquanto fica observando a fonte de perigo, embora não esteja suficientemente estimulada para se movimentar.
O agachamento diante de um intruso parece ser utilizado pelos quero-queros como tentativa de se esconder, sendo uma resposta de defesa primária críptica pouco utilizada pelos adultos. Quando o escape foi realizado em grupo, as aves sobrevoaram a área de ocupação, e ao aterrissarem, voltaram às atividades de vigilância ou de manutenção.
No cativeiro, a perseguição foi pouco freqüente, provavelmente por serem sempre as mesmas aves que habitavam o local: Eudocimos ruber (guará-rubra), Tigrisoma lineatum (socó-boi), Casmerodius albus (garça-branca), Euxenura maguari (maguari), Balearica pavonina (grou coroado), Aix sponsa (marreca carolina), Amazonetta brasiliensis (marreca ananaí), Threskiornis aethiofica (ibis sagrado), Larus dominicanus (gaivotão), Sterna hirundinacea (trinta réis), Phoenicopterus ruber roseus (flamingo), Aix galericulata (marreco mandarim). O resultado sugere a existência de um processo de habituação nos quero-queros. Foi apenas durante a distribuição do alimento realizado pelos tratadores que as aves, disputando o alimento, perseguiram-se umas às outras.
O comportamento de fuga foi gradativamente substituído, com o início do período reprodutivo, pelo comportamento de manobras de distração (categoria de movimento agachado) exclusivo do período reprodutivo (Figura 3). Estas manobras realizaram-se no limiar entre a fuga, no sentido de auto proteção, e a tática de desviar o intruso, embora não estivesse presente, durante a etapa de acasalamento, estimulação específica do ninho e dos filhotes (Costa, 1985, 1994b). O comportamento de manobra foi estudado em outras espécies de Charadriiformes por Armstrong (1949a, 1949b), Simmons (1952, 1955) e por Willianson (1948a, 1948b, 1952).

Figura 3: No período não reprodutivo, os quero-queros defendem-se através da fuga (PFU), gradativamente substituída pelo movimento agachado (PMA) no início da fase de corte do estágio reprodutivo. A categoria de alerta (PAL) ocorre nos períodos não reprodutivo e reprodutivo. O ataque em vôo (PVA) e o movimento agachado (PMA) são exclusivos do período reprodutivo. C: corte, N: nidificação e CP: cuidado parental.
Mesmo na ausência de um invasor, as manobras de distração foram intercaladas com comportamentos de manutenção (alisamento da plumagem, coçar-se, alimentação, banho e repouso), alerta e deslocamento. Na área urbana, as manobras de distração ocorreram no final do período não reprodutivo, demonstrando que alguns dos subgrupos de quero-queros já apresentavam um comportamento preparatório para a reprodução (Costa, 1999).
Houve uma relação entre o período reprodutivo e a execução de determinadas categorias. Constatou-se que o terceiro quero-quero (a ave C), embora fosse um componente do trio reprodutivo, não apresentou inicialmente categorias características da reprodução, desempenhando em alta freqüência a postura de fuga típica do período não reprodutivo.
No decorrer do ritual de corte, da elaboração do ninho e da cópula, as manobras de distração permaneceram no repertório comportamental dos quero-queros, diminuindo a sua freqüência apenas durante o cuidado dos filhotes com mais de três semanas. A ausência de manobras de distração no cativeiro decorreu de uma ausência do estímulo reprodutivo.
As duplas e os trios responderam às invasões com a postura de agachamento. Esta postura foi utilizada como fuga e como manobra de distração, diferenciando-se pelo modo como se dá a abertura das asas. Na manobra de distração, as asas estão abertas, enquanto que, na fuga, as asas ficam junto ao corpo (Costa, 1994b). Gallegos (1984) observou, em Vanellus chilensis, o agachamento-manobra como "parada de falso ninho" antes da nidificação e depois do nascimento dos filhotes. Hall (1964) observou o mesmo comportamento em Haplopterus armatus (Vanellus armatus) após a eclosão dos ovos.
As manobras de distração e de vigilância continuaram a ser realizadas durante a incubação dos ovos. Neste período, ao mesmo tempo em que os quero-queros escapavam do intruso, dificultavam a localização exata do ninho. Durante uma invasão, o quero-quero que estava no ninho executava manobras de distração e alerta, tanto nas duplas como nos trios reprodutivos. O comportamento de se afastar do ninho, realizado pelos quero-queros também foi observado por Oniki (1986). Evidenciou-se que
cada quero-quero componente do trio teve uma função específica no comportamento defensivo: a ave A cuidou de desviar o invasor do ninho e as aves B e C desempenharam comportamentos de defesa. As três aves mantiveram-se em vigilância. No trio observou-se o comportamento de fuga no quero-quero C.
Elliot (1985a, 1985b) e Cramp e Simmons (Berg, 1993) observaram que tanto machos como fêmeas de Vanellus vanellus atacaram os predadores, independentemente, no período de incubação.
Vanellus chilensis possivelmente tenha aprendido como Vanellus spinosus (Walters, 1990) a identificar os tratadores que se aproximam de seus ovos. No Jardim Botânico, os funcionários eram atacados mesmo quando estavam distantes do ninho. Quando perseguidos, os quero-queros executaram o movimento agachado e o agachamento-distração. Neste comportamento, as manobras de distração funcionaram como uma atitude de escape e, ao mesmo tempo, desviaram o intruso da área territorial, permanecendo o ninho protegido.
Durante a incubação dos ovos, o ataque foi uma estratégia de defesa vinculada exclusivamente ao ninho, pois os quero-queros realizaram outras atividades como ameaças e manobras de distração sem confronto físico. As posturas de ameaça, ameaça 2, preconizadora e desencadeadora de ataque apresentaram-se em baixa freqüência. As manobras de distração (movimento agachado e agachamento) prevaleceram em todas as respostas dadas pelo trio, independentemente dos tipos ou modos de invasão. O ataque foi realizado por todos os componentes do grupo reprodutivo quando o intruso estava a 10 metros do ninho. Confirmam-se assim as observações de Elliot (1985a, 1985b) que verificou que Vanellus vanellus diminuiu o comportamento defensivo à medida que o predador se afasta do ninho. Ward (1989) observou que o ataque foi realizado estando o invasor a cinco metros (Vanellus coronatus) ou a 10 metros do ninho (Vanellus melanopterus). Ward verificou que Vanellus lugubris não atacou nem perseguiu intrusos durante o período de incubação, enquanto que um espécime de Vanellus melanopterus, nesta fase, só atacou quando o companheiro não foi capaz de expulsar o intruso. Oniki (1986) observou o quero-quero levantar-se do ninho quando o intruso estava a 150 metros de distância, na área rural, e a sete metros, no campus da Universidade Estadual Paulista em Rio Claro, São Paulo.
Os quero-queros das duplas reprodutivas mostraram respostas semelhantes aos invasores ao realizarem a postura de movimento agachado durante o cuidado parental. O ataque foi efetuado até a segunda semana de desenvolvimento dos filhotes.
As defesas com ataque foram mais freqüentes no período reprodutivo (212 ocorrências) do que no período não reprodutivo (16 ocorrências), em 228 observações. Estes resultados vêm ao encontro das observações de Walters (1979, 1982) quando comenta que a agressão das diferentes espécies de Vanellus, em direção a outras espécies de aves intrusas nos seus territórios, relacionam-se ao comportamento parental.
O comportamento de ataque iniciou-se na nidificação e prolongou-se até o que os filhotes tivessem cerca de duas a três semanas de idade (Figura 3). Os ataques ocorreram com mais freqüência quando o intruso encontrava-se a menos de cinco metros do filhote, independentemente da idade deste.
Os quero-queros em todas as situações realizaram comportamentos defensivos, sem confronto físico, apresentando uma pequena freqüência de condutas de ameaça. As manobras de distração com a postura de movimento agachado prevaleceram em todas as respostas defensivas, independentemente do modo de invasão e do tipo de grupo reprodutivo.
O ataque dos quero-queros a predadores como o carcará foi abaixo do esperado, não sendo observado ataques a corvos (Elliot, 1985a, 1985b; Kirby & Green, 1991; Mcmahon, 1996), a Milvago chimango (Zuberbühler, 1973) e o falcão (Thomas, 1983).
Oniki (1986) observou Caracara plancus e Coragyps atratus como predadores dos quero-queros. Gallegos (1984) detectou Milvago chimango, Caracara plancus, pessoas a pé e carros como intrusos visualizados pelos quero-queros a mais de 200 metros.
Galbraith (1989) detectou como predadores de Vanellus vanellus, Falco peregrinus, Accipiter nisus, Buteo buteo, Falco tinnunculus, Mustela erminea, gatos e cães. Corvus corone predam ovos e filhotes de Vanellus vanellus. Galbraith observou que Vanellus vanellus localizava-se sempre distante de árvores, como um comportamento estratégico para inviabilizar a visualização dos predadores.
Possivelmente o número de pessoas e aves atacadas tenham sido freqüentes pelo fato do ambiente antrópico, predispondo os quero-queros a estes intrusos, enquanto que o número de predadores foi pouco freqüente, pois no ambiente urbano o número de predadores naturais dos quero-queros é menor que no ambiente rural.
Os resultados obtidos permitiram, através do estudo das categorias comportamentais, um detalhamento dos tipos de respostas defensivas dos quero-queros em diferentes circunstâncias de invasão por intrusos de outras espécies, nas áreas urbana e rural. Comprovou-se a importância do comportamento de alerta através da elevada freqüência com que é realizado e das categorias relacionadas com as manobras de distração exclusivas do período reprodutivo. Estabeleceu-se que as estratégias de defesa ocorreram de acordo com a distância do intruso do ninho, dos filhotes e dos quero-queros adultos. Um resultado importante foi a comprovação que o ataque não é a estratégia de defesa interespecífica mais freqüente, ocorrendo apenas quando o intruso está a menos de cinco metros do ninho e dos filhotes. Outro aspecto relevante foi a verificação que o comportamento de fuga é característico do período não reprodutivo e que diante de um intruso os quero-queros responderam primeiramente sem vocalização.
Nosso estudo tem relevância na medida em que desvenda algumas das características básicas do modo como os quero-queros, Vanellus chilensis, reagem defensivamente em seus territórios, em função do estágio reprodutivo. Será importante coletar outras informações, por exemplo sobre as estratégias reprodutivas e sobre o cuidado parental que complementem os dados aqui apresentados e que permitam uma compreensão maior do comportamento e da organização social da espécie.
Referências
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-267. [ Links ]
Armstrong, E. A. (1949a). Diversionary display. Part 1 _ Commotation and terminology. Ibis, 91, 88-97. [ Links ]
Armstrong, E. A. (1949b). Diversionary display Part 2 _ The nature and origin of distraction display. Ibis, 91, 179-188. [ Links ]
Barnard, C. J., & Stephens, H. (1981). Prey size selection by lapwings in lapwings/gull associations. Behaviour, 77(1-2), 1-22. [ Links ]
Beintema, A. J., & Visser, G. H. (1989a). The effect of weather on time budgets and development of chicks of meadow birds. Ardea, 77(2), 181-192. [ Links ]
Beintema, A. J., & Visser, G. H. (1989b). Growth parameters in chicks of charadriiform birds. Ardea, 77(2), 169-180. [ Links ]
Berg, A. (1993). Habitat selection by monogamous and polygamous lapwings on farmland: the importance of foraging habitats and suitable nest sites. Ardea, 81(2), 99-105. [ Links ]
Buitron, D. (1983).Variability in the responses of black - billed magpies to natural predators. Behaviour, 78, 209-236. [ Links ]
Caraco, T; Martindale, S & Pulliam, R. (1980). Avian flocking in the presence of a predator. Nature, 285, 400-401. [ Links ]
Costa, L. C. M. (1985). Aspectos comportamentais de Vanellus chilensis (Wagler, 1827) (Aves, Charadriiformes) em Curitiba, Paraná. Dissertação de mestrado, Universidade Federal do Paraná, Curitiba, PR. [ Links ]
Costa, L. C. .M. (1994a). Comportamento agonístico de Vanellus chilensis (Wagler, 1827) (Aves, Charadriiformes) em Curitiba, Paraná, Brasil. Revista Acadêmica, 9, 21-30. [ Links ]
Costa, L. C. M. (1994b). Manobras de distração de Vanellus chilensis (Wagler,1827) (Charadriiformes, Charadriidae) em Curitiba, Paraná, Brasil. Estudos de Biologia, 3(36), 33-42. [ Links ]
Costa, L. C. M. (1999). Análise do comportamento agonístico de Vanellus chilensis (Molina, 1782) (Charadriiformes, Charadriidae). Tese de doutorado, Universidade Federal do Paraná, Curitiba, PR. [ Links ]
Dann, P. (1981). Breeding of the banded and masked lapwings in Southern Victoria. Emu, 81(1-2), 1-22. [ Links ]
Elliot, R. D. (1985a). The effects of predation risk and group size on the anti-predator responses of nesting lapwings Vanellus vanellus. Behaviour, 92(1-2), 168-187. [ Links ]
Elliot, R .D. (1985b). The exclusion of avian predators from aggregations of nesting lapwings (Vanellus vanellus). Animal Behaviour, 33(1), 308-314. [ Links ]
Galbraith, H. (1989). Arrival and habitat use by lapwing Vanellus vanellus in the early breeding season. Ibis, 131(3), 377-388. [ Links ]
Gallegos, L. D. (1984). Aspectos de la biologia reproductiva del tero comum Vanellus chilensis (Gmelin). 1: comportamiento y territorialidad. Hornero, 12(3), 150-155. [ Links ]
Grisser, P. (1990). Anti-predator postures of lapwing and stonechat. British Birds, 83(10), 425. [ Links ]
Hall, K. R. L. (1964). A study of the Blacksmith Plover (Holopterus armatus) in the Cape Town Area II. Behaviour, 35(1), 3-16. [ Links ]
Hesp, L. S., & Barnard, C. J. (1989). Gulls and plovers:age-related differences in kleptoparasitism among black headed gulls (Larus ridibundus ). Behavioral ecology and sociobiology, 24(5), 297-304. [ Links ]
Iversen, F. M. (1986). The impact of disturbance on the lapwings Vanellus vanellus incubation. Dansk Ornitologisk Forenings Tidsskrift, 80(3-4), 97-102. [ Links ]
Kalsi, S. R., & Khera, S. (1987). Agonistic and distraction behaviour of the redwattled lapwing, Vanellus indicus indicus. Pavo, 25(1-2), 43-56. [ Links ]
Kirby, J. S., & Green, R. E. (1991). Nest defence by black-tailed godwits on the Ouse Washes, Cambridgeshire, England. Wader Study Group Bulletin, 61, 71-72. [ Links ]
Lehner, P. N. (1996). Handbook of ethological methods. New York: Garland. [ Links ]
Mcmahon, N. (1996). Carrion crows killing several northern lapwings. Bristish Birds, 89(6), 278. [ Links ]
Myers, J. P. (1978). One deleterious effect of mobbing in the southern lapwing (Vanellus chilensis). Auk, 95(2), 419-420. [ Links ]
Oniky, Y. (1986). Nesting of the southern lapwing Vanellus chilensis in São Paulo, Brazil. Avicultural Magazine, 92(3), 151-156. [ Links ]
Ruszczyk, A., Rodrigues, J. J. S., Roberts, T. M. T., Bendati, M. M. A., Del-Pino, R. S., Marques, J. C. V., & Melo , M. T. Q. (1987). Distribution patterns of eight bird species in the urbanization of Porto Alegre, Brazil. Ciência e Cultura, 39(1), 14-19. [ Links ]
Sick, H. (2001). Ornitologia brasileira. Rio de Janeiro: Nova Fronteira. [ Links ]
Simmons, K. E. L. (1952). The nature of the predator reactions of breeding birds. Behaviour, 4, 161-171. [ Links ]
Simmons, K. E. L. (1955). The nature of the predator-reactions of waders towards humans; with special reference to the role of the aggressive, escape and broding-drives. Behaviour, 8, 130-173. [ Links ]
Thomas, D. H. (1983). Aposematic behaviour in the blacksmith plover. Ostrich, 54(1), 51-52. [ Links ]
Thompson, D. B. A. (1983). Prey assessment by Plovers (Charadriidae): net rate of energy intake and vilnerability to kleptoparasites. Animal Behaviour, 31, 1226-1236. [ Links ]
Thompson, D. B. A . (1986). The economics of kleptoparasitism : optimal foranging, host and prey selection by gulls (Larus ridibundus). Animal Behaviour, 34(4), 1189-1205. [ Links ]
Thompson, D. B. A. & Barnard, C. J. (1983). Anti-predator responses in mixed-species associations of lapwings (Vanellus vanellus), golden plovers (Pluvialis apricaria) and black-head gulls (Larus ridibundus). Animal Behaviour, 31(2), 585-593. [ Links ]
Thompson, D. B. A., & Barnard, C. J. (1984). Prey selection by plovers (Pluvialis apricaria): optimal foraging in mixed-species groups. Animal Behaviour, 32(2), 554-563. [ Links ]
Thompson, D. B. A., & Lendren, D. W. (1985). Gulls and plovers: host vigilance, kleptoparasite succces and a model of kleptoparasite detection. Animal Behaviour, 33(4), 1318-1324. [ Links ]
Walters, J. (1979). Interspecific aggressive behavior by long-toed lapwings (Vanellus crassirostris). Animal Behaviour, 27(4), 969-981. [ Links ]
Walters, J. R. (1980). The evolution of parental behavior in lapwings. Tese de doutorado, University of Chicago. [ Links ]
Walters, J. R. (1982). Parental behavior in lapwings (Charadriidae) and its relationships with clutch sizes and mating systems. Evolution, 36(5), 1030-1040. [ Links ]
Walters, J. R. (1990). Anti-predatory behavior of lapwings:field evidence of discriminative abilities. Wilson Bulletin, 102(1), 49-70. [ Links ]
Walters, J., & Walters, B. J. (1980). Cooperative breeding by southern lapwing in Colombia. Ibis, 122(4), 505-509. [ Links ]
Ward, D. (1989). Behaviour associated with breeding of crowned, blackwinged and lesser blackwinged plovers. Ostrich, 60(4), 141-150.
Willianson, K. (1948a). Eider duck plucking down during distraction display. Ibis, 90, 142-143. [ Links ]
Willianson, K. (1948b). Field-notes on nodification and distraction-display in the Golden Plover. Ibis, 90, 90-98. [ Links ]
Willianson, K. (1952). Regional variation in the distraction displays of the Oystecatcher. Ibis, 94, 85-96. [ Links ]
Zuberbhuler, E. A. (1973). Notas ecológicas. Hornero, 11(3), 177-192. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Leny Cristina Milléo Costa
Pontifícia Universidade Católica do Paraná
R. Imaculada Conceição, 1155 - Prado Velho
80215-901 Curitiba-PR
E-mail: lcmcosta@rla01.pucpr.br
Recebido em 22 de março de 2002
Revisão recebida em 22 de abril de 2003
Aceito em 07 de maio de 2003
*Este trabalho é parte da tese de doutorado "Análise do comportamento agonístico de Vanellus chilensis (Molina, 1782) (Charadriiformes, Charadriidae)" apresentada ao Curso de Pós-graduação em Ciências Biológicas - Zoologia da Universidade Federal do Paraná, 1999. Agradeçoaos Professores Drs. Vinalto Graf, César Ades, Paulo Nogueira Neto, Luís dos Anjos, Emygdio A. Monteiro Filho, Sebastião Laroca e Marta Luciane Fischer pelos comentários críticos.

