











 (pdf)
(pdf)




 Permalink
PermalinkRevista de Etologia
ISSN 1517-2805 ISSN 2175-3636
Rev. etol. v.6 n.1 São Paulo jun. 2004
ARTIGOS
Female social dominance does not establish mating priority in Nile tilapia*
A dominância social da fêmea não promove prioridade de acasalamento em fêmeas de tilápia-do-Nilo
Eliane Gonçalves-de-Freitas**; Aline Chimello Ferreira**
Universidade Estadual Paulista, São José do Rio Preto
ABSTRACT
Female dominance and size are important features for mating choices in fishes. As dominants are also the largest females in some species, we tested whether dominance per se favours mating priority in Nile tilapia females. Three matched-size adult females (n = 13) were grouped in a 140 l-aquarium during 3 days for hierarchical settlement. On the 4th day a male larger than the females was introduced in the group and kept there for 12 consecutive days until reproduction. Spawning occurred in 8 out of 13 replicates, and the alpha female was the first to mate in only 50% of the cases. Males may thus choose females according to size and not according to rank, as occurs in other cichlid species. As confrontations and the establishment of hierarchical dominance involve high energetic expenditure in Nile tilapias, the reason why females invest in such behaviors is still a question to be elucidated.
Index terms: Reproductive behavior, Mate choice, Nile tilapias. Oreochromis niloticus.
RESUMO
A dominância e o tamanho da fêmea são características importantes para a escolha do parceiro sexual em peixes. Como as fêmeas dominantes também são as maiores em várias espécies, procuramos verificar se a dominância em si promove a prioridade de acasalamento em fêmeas de tilápia-do-Nilo. 13 grupos de 3 fêmeas adultas de tamanhos similares foram mantidas em aquários de 140 l para o estabelecimento da hierárquia entre cada 3 indivíduos. No quarto dia, um macho adulto foi introduzido no grupo e mantido lá por 12 dias para reprodução. Ocorreu desova em 8 réplicas, sendo que a fêmea alfa somente foi a primeira a acasalar em 4 destes casos. É possível que os machos de tilápia do Nilo escolham as fêmeas usando o tamanho como critério, como em outros ciclídeos, e não pela posição social. Uma vez que os confrontos envolvem um elevado gasto energético para a tilápia-do-Nilo, o motivo pelo qual as fêmeas investem nesse comportamento ainda é uma questão a ser resolvida.
Descritores: Comportamento reprodutivo, Escolha do parceiro, Tilápia-do-nilo. Oreochromis niloticus.
Social dominance is a widespread feature among vertebrates. In fish, dominance establishment is less complex than it is in so-called higher vertebrates, but it also envolves aggressive displays, physical confrontations and ritualized fights (e.g. Baerends & Baerends-Van Roon, 1950; Frier & Iles, 1971; Sloman & Armstrong, 2002).
Dominant status gives priority for reproduction in male cichlid fishes. The animals defend a territory where, according to the species, spawning or also broodcare takes place (Baerends & Baerends-Van Roon, 1950; Frier & Iles, 1972; Mackaye, 1986). Dominance also gives advantage to females, which can reproduce first in a reproductive season (Brandtman, Scandura & Trillmich, 1999; Weber & Weber, 1976). For some species, subordinate males (Borowsky, 1987) or females (Rosenqvist, 1990) have their reproduction inhibited, an indication that social status can also affect the reproductive physiology of fishes.
Male or female mate preference may be modulated by several factors which signal possible genetical or non genetical benefits to the brood, as size, fecundity, bright color, territory, status etc. (Turner, 1996). Selection for female fecundity or increased probability of paternity are also immediate benefits for males, leading them (not always) to choose large females (Barlow, 1986; Mackaye, 1986; Rosenqvist, 1990). Both sexes may, moreover, choose as mates those which are more efficient protectors of young (Forsgreen, 1997; Turner, 1996). Status also may be a feature which determines mate choice, as it may give information about the partner's condition and capacity to broodcare, to compete for environmental resources or to simply survive (e.g. Berglund & Rosenqvist, 2001).
Nile tilapia, Oreochromis niloticus (L.), is a cichlid species with social dominance hierarchies and a polygynic-polyandric mating system, in which dominant males have priority for mating (Gonçalves-de-Freitas & Nishida, 1998). According to Gonçalves-de-Freitas and Nishida (1998), females are also aggressive and fight for a high position in the group. Reproduction is not inhibited in subordinate males or females (Gonçalves-de-Freitas & Nishida, 1998; Gonçalves-de-Freitas, 1999) but, though the advantage of being dominant is clear for males, this has not be shown to be the case for females.
Social dominance allows mating priority for females of other cichlid species, but dominant females in such species are also the biggest females in the group (Brandtman et al., 1999; Weber & Weber, 1976): choice might then depend on size rather than on dominance per se, or jointly on size and dominance. Size plays an important role in male mate choices in fishes (e.g. Brandtman et al., 1999; Rosenqvist, 1990). In the present experiment, we intended to examine whether dominance as such is sufficient to influence mating preference in the female Nile tilapia.
Method
The animals were acquired from a commercial supplier and maintained in water from an artesian well supply (with no chlorine) at room temperature for 30 days before experimentation. Thirteen groups of 3 females were formed in order to determine a hierarchical rank between them. Their behavior was video-recorded during three consecutive days (12 min/day). A male was then introduced into each group and maintained there for 12 days to mate. The groups were kept in 140 L-aquaria with aerated and no chlorinated water, and with a layer of gravel to allow nest building by the male. Biological filters were used to keep the water in good quality and, moreover, the aquarium was siphoned every 4 days to remove faeces and food leftovers from the bottom. Water temperature was kept at 27°C and a 12L:12D photoperiod was used. The fish were daily fed to satiation with commercial food for tropical fish. The mean LS and weight of the females was 10.65 ± 1.29 cm and 45.63 ± 17.13 g (n = 39) and of the males 12.92 ± 1.21 cm and 77.79 ± 25.20 g (mean ± SD, n = 13) respectively. Fish sex was identified by means of inspection of genital papillae stained with methylen blue (Afonso & Lebout, 1993) and each individual was recognized by having a small piece of the caudal fin cut, according to Fernandes and Volpato (1993). Fish were anesthetized with benzocaine (2:5,000) before manipulation.
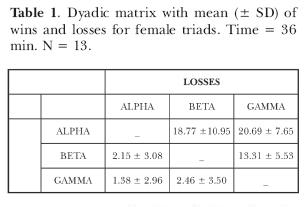
The record of agonistic interaction was based on the ethogram described by Alvarenga and Volpato (1995) for this species. The dominance was inferred from a dyadic interaction matrix during the three days of observation (Lehner, 1995), and according to rules by Boyd and Silk (1983) for the determination of dominant-subordinate relationships.
The rules established to identify winners and losers in each contest and to build matrices were as follows: A fish wins if it (1) chases another fish that flees; (2) attacks and is not attacked; (3) spreads its dorsal fin causing flight or another subordinate display in the opponent fish. A fish loses if it (1) flies from attacks or chases; (2) does not fight back when attacked; (3) withdraws its dorsal fin when attacked; (4) changes the body or eyes color pattern to dark.
Rank was inferred from the sum of contests in the 3 sessions of observation (36 min). Results were put in a matrix to identify the alpha, beta and gamma female (Lehner, 1996). After the introduction of the male, the groups were examined daily in order to check for mouthbrooding females. At the end of the experiment, fish were killed with an overdose of benzocaine and dissected for gonad analysis to confirm their sex and maturity, which was inferred from macroscopic analysis as described in Paiva, Verani, Mainardes-Pinto and Tabata (1988).
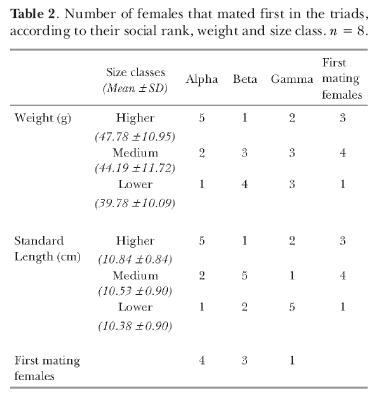
Although triads were formed according to female size equivalence, we examined whether slight differences in size and weight could determine social rank. Female were classified into higher, medium and lower size (or weight) and the frequency of females in each class was compared using a Chi-Square Test (Zar, 1999). Association among social rank and first spawning followed the same procedure.
Results and discussion
Females displayed a linear hierarchy (Table 1). Lack of association between social dominance and small differences in size and weight (p > 0.05, Table 2) shows that alpha position was settled by other factors and indicates that the control of fish size was adequate. Moreover, there was no association between social dominance and priority for mating (p > 0.05, Table 2) since in only 4 cases, alpha females reproduced first. Beta female mated first in 3 cases and gamma females in 1 case.
Although the macroscopic analysis of the gonads showed the fish were mature, spawning was only observed in eight cases. Analyses were performed with such eight replicates.
It may be assumed that it is advantageous for the alpha females Nile tilapia to mate first because female performance in agonistic contests (or simply rank signals) could indicate her fitness for brood care, stimulating male mate choice. In addition, alpha females could enhance their reproductive success by being ready to mate again, after brood care (Lowe-Macconnell, 1958). The alpha female could thus be the first to return to the reproductive arena and reproduce one or more times. Our results do not confirm such assumptions.
According to Brandtmann et al. (1999), dominant females have priority for access to mates in the cichlid Lamprologus ocellatus. This was also observed in Cichlasoma nigrofasciatum (Weber & Weber, 1976). Nevertheless, in both species, dominants are also the biggest females and, in fish, the choice of the biggest female is important to males because fertility increases with size (Brandtmann et al., 1999) and enhances the reproductive success of both male and female. This was not verified, in our experiment, for slight differences in size in Nile tilapia, since association between reproduction and length or weight was also not significant (p > 0.05; table 1). Therefore, other factors, such as chemical or visual stimuli which indicate female reproductive condition (Silverman, 1978; Souza, Lucion & Wasserman, 1998), might be decisive for mating instead of hierarchical rank.
Dominance is not always the main factor determining mate choice in fishes. According to Östlund Nilsson and Nilsson (2000), dominant male stickleback prevent subordinate to reproduce but, in condition of free choice, female sticklebacks do not show preference for dominant males. In sand gobies, a fish species that exhibit paternal care, females choose good fathers instead of dominant males (Forsgren, 1997). As in such species, male mate choice in the Nile tilapia seems not to be correlated with female fighting ability, but with others features which can be accessed during courtship.
In our study, spawning was not associated neither to dominance nor to minimal differences in females' size. However, it is possible that males choose females according to perceptible differences among their sizes, as occurs to other fish species and not by means of hierarchical rank. As contests and social dominance involve an elevated energetic cost for Nile tilapia (Alvarenga & Volpato, 1995; Fernandes & Volpato, 1993), the reason why females of this species invest in such a behavior is still a question to be elucidated.
References
Afonso, L.O.B. & Lebout, E.L. (1993). Métodos para sexagem visual de alevinos de tilápia nilótica (Oreochromis niloticus). In Zimmermann, S. (ed.). Anais do IV Encontro Rio Grandense de técnicos em aquicultura (pp. 100-103). Porto Alegre, Universidade Federal do Rio Grande do Sul. [ Links ]
Alvarenga, C.M.D. & Volpato, G.L. (1995). Agonistic profile and metabolism in alevines of Nile tilapia. Physiology and.Behavior 57, 75-80. [ Links ]
Baerends, G.P., & Baerends-Van Roon, J.M. (1950). An introduction to the study of the ethology of cichlid fishes. Behaviour Supplement, 1,1-242. [ Links ]
Barlow, G.W. (1986). Mate choice in the monogamous and polychromatic Midas cichlid, Cichlasoma citrinellum. Journal of Fish Biology, 29 (Suppl. A), 123-133. [ Links ]
Berglund, A. & Rosenqvist, G. (2001). Male pipefish prefer dominant over attractive females. Behavioral Ecology, 12, 402-406. [ Links ]
Borowsky, R.L. (1987). Agonistic behavior and social inhibition of maturation in fishes of the genus Xyphophorus (Poecilidae). Copeia, 3, 792-796. [ Links ]
Boyd, R. & Silk, J.B. (1983). A method for assigning cardinal dominance ranks. Animal Behaviour, 31, 45-58. [ Links ]
Brandtmann, G., Scandura, M. & Trillmich, F. (1999). Female-female conflict in the harem of a snail cichlid (Lamprologus ocellatus) : Behavioural interactions and fitness consequences. Behaviour, 136, 1123-1144. [ Links ]
Fernandes, M.O. & Volpato, G.L. (1993). Heterogeneous growth in the Nile tilapia: social stress and carbohydrate metabolism. Physiology and Behavior, 54, 319 - 323. [ Links ]
Forsgreen, E. (1997) Females sand gobies prefer good fathers over dominant males. Proceedings of the Royal Society of London, Series B-Biological Sciences, 246, 1283-1286. [ Links ]
Fryer, G. & Iles, T.D. (1972). The Cichlid Fishes of the Great Lakes of Africa. T.F.H. publications, Neptune City. [ Links ]
Gonçalves-de-Freitas, E. & Nishida, S. M. (1998). Sneaking behavior of the Nile tilapia. Boletim Técnico do CEPTA, 11, 71-79. [ Links ]
Gonçalves-de-Freitas, E. (1999). Investimento reprodutivo e crescimento em machos de tilápia-do-Nilo. Tese de doutorado. Instituto de Biociências, Universidade Estadual Paulista, Botucatu, SP. [ Links ]
Lehner, P.N. (1996). Handbook of Ethological Methods (2nd ed). Cambridge University Press, Cambridge, UK. [ Links ]
Lowe-McConnell, R. (1958). Observations on the biology of Tilapia nilotica Linné in east Africa waters (Pisces: Cichlidae). Revue de Zoologie et Botanique Africaines, 57, 129-170. [ Links ]
MacKaye, K.R. (1986). Mate choice and size assortative pairing by the cichlid fishes of Lake Jiloa, Nicaragua. Journal of Fish Biology, 29 (Supp. A), 135-150. [ Links ]
Óstlund Nilsson, S. & Nilsson, G.E. (2000). Free choice by female sticklebaks: lack of preference for male dominance traits. Canadian Journal of Zoology, 78, 1251-1258. [ Links ]
Paiva, P., Verani, J.R., Mainardes-Pinto, C.S.R. & Tabata, Y.A. (1988). Studies on the growth and reproduction in the cichlid Oreochromis niloticus (LINNAEUS, 1757) in a earthen pond. Boletim do Instituto de Pesca, 15, 109-114. [ Links ]
Rosenqvist, G. (1990). Male mate choice and female-female competition for mates in the pipefish Nerophis ophidion. Animal Behaviour, 39, 1110-1115. [ Links ]
Silvermann, H. (1978). Effects of different levels of sensory contact upon reproductive activity of adult male and female Sarotherodon (Tilapia) mossambicus (Peters); Pisces: Cichlidae. Animal Behavior, 26, 1081-1090. [ Links ]
Sloman, K.A. & Armstrong, J.D. Physiological affects of dominance hierarchies: laboratory artefacts or natural phenomena? Journal of fish Biology, 61, 1-23. [ Links ]
Souza, S.M.G., Lucion, A.B. & Wassermann, G.F. (1998). Influence of 17a,20b-dihidroxi-4-pregnen-3-one, injected into a post-ovulatory female on the reproductive behaviour of male Nile tilapia (Oreochromis niloticus). Comparative Biochemistry and Physiology, 119 A, 759-763. [ Links ]
Turner, G.F. (1996). Teleost mating behaviour. In Pitcher, T.J. (ed.). Behaviour of Teleost Fishes (2nd ed.). Chapman & Hall, London. [ Links ]
Weber, P.G. & Weber, S.P. (1976). The effect of female color, size, dominance and early experience upon mate selection in male convict cichlids, Cichlasoma nigrofasciatum Guenther (Pisces, Cichlidae). Behaviour, 56, 116-135. [ Links ]
Zar, J.H. (1999). Bioestatistical Analysis (4th ed.). Prentice Hall, New Jersey. [ Links ]
 Correspondence
Correspondence
Eliane Gonçalves-de-Freitas
Laboratório de Comportamento Animal, Dep. Zoologia e Botânica, IBILCE, UNESP (CAUNESP)
R. Cristóvão Colombo, 2265,
15054-000, São José do Rio Preto, SP, Brasil
E-mail: elianeg@dzb.ibilce.unesp.br
Received September 16, 2003
Revision received November 18, 2003
Accepted March, 2004
* Results were presented at the XXVIII International Ethological Conference, Brazil, 2003. The authors thank G. L. Volpato (UNESP, Botucatu) for profitable suggestions; O. Pissolato for fish donation; R. S. C. Ferreira and C. E. Souza for technical support; J.J. Gimenez, E.F.L. Freitas, A. L. S. Castro and T. C. Mariguela for assistance in collecting animals and helping in other phases of this study.
** Laboratório de Comportamento Animal, Dep. Zoologia e Botânica, IBILCE, UNESP (CAUNESP).

