










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.7 n.2 São Paulo dez. 2005
ARTIGOS
Estudo de caso: a reutilização de células de ninho abandonado de Polistes (Aphanilopterus) simillimus Zikán, 1951 (Hymenoptera: Vespidae, Polistinae) por Tetrapedia (Tetrapedia) diversipes Klug, 1810 (Hymenoptera: Apidae, Apinae)
Case study: the re-utilization of abandoned nest cells of Polistes (Aphanilopterus) simillimus Zikán, 1951 (Hymenoptera: Vespidae, Polistinae) by Tetrapedia (Tetrapedia) diversipes Klug, 1810 (Hymenoptera: Apidae, Apinae)
Nozor Paulo Outeiro Pinto1
Universidade Estadual Paulista, Campus de Rio Claro
RESUMO
Espécies de abelhas e vespas solitárias podem reutilizar as cavidades de ninhos antigos de Hymenoptera, assim como células construídas de fibras vegetais ou barro em ninhos abandonados, adaptando-os à geração de sua descendência. Este é um método simples e eficiente de nidificação porque implica em menor esforço, economia de tempo e menor risco de predação à fêmea reutilizadora. A vespa social Polistes simillimus constrói ninhos de fibras vegetais que são fixados a um substrato por um pedicelo ficando os favos expostos. Ninhos da abelha solitária Tetrapedia diversipes tem sido encontrados em cavidades em madeira, nas cavidades dos ninhos armadilhas e podem estar associados às células de ninhos antigos de outros himenópteros. Foi encontrado em ninho abandonado de P. simillimus células reutilizadas por T. (T.) diversipes para nidificação. Esta observação ainda não havia sido relatada.
Descritores: Nidificação, Vespas, Abelhas, Polistes simillimus, Tetrapedia (Tetrapedia) diversipes.
ABSTRACT
Species of solitary wasps and bees may re-utilize holes of old nests of Hymenoptera as well as cells made of vegetable fibers or mud in abandoned nests, and adjust them to their own offsprings. This simple and efficient nesting strategy involves smaller effort, time and risk of predation to reutilizer female. The social wasp Polistes simillimus makes nests of vegetable fibers which are attached to a substract by a pedicel leaving the combs exposed. Nests of the solitary bee Tetrapedia diversipes have been found in holes in wood, trap nests and their nests may be associated to cells of old nests of others Hymenoptera. In a abandoned nest of P. simillimus nest cells re-utilized for nesting T. (T.) diversipes was found. This is the first report of XXXX reutilization.
Index terms: Nesting behavior, Wasps, Bees, Polistes simillimus, Tetrapedia (Tetrapedia) diversipes.
O comportamento de nidificação da maioria das espécies de vespas é efetuado solitariamente (Evans & Eberhard, 1970; Spradbery, 1973) o mesmo ocorrendo com as espécies de abelhas (Alves-dos-Santos, 2000, 2002; Ayasse, Paxton, & Tengö, 2001; Buchmann, 1987; Michener, 1974, 2000; Roubik, 1989; Spradbery, 1973). Na maioria das espécies de abelhas cada fêmea faz o seu próprio ninho, ou algumas vezes vários deles, sem buscar as locações dos outros ninhos da mesma espécie (Michener, 1974).
Ninhos abandonados de vespas e abelhas, dependendo dos locais que os abriguem, podem permanecer em bom estado de conservação após serem abandonados e este fato, pode, as vezes, permitir que sejam reusadas, com adaptações, pelas gerações seguintes das construtoras e mesmo por outras fêmeas solitárias de outras espécies de himenópteros. As vespas inquilinas e as que habitam em buracos estreitos são oportunistas e ocupam ampla variedade de cavidades preexistentes, normalmente buracos vazios usados por outros insetos em madeira seca, como ocorre com as vespas xilófilas (Cowan, 1991), podendo, também, ocupar as células vazias de ninhos construídos por outras espécies de vespas (Iwata, 1976, citado por Cowan, 1991). As reutilizações podem ocorrer em células de ninhos ativos e em células de ninhos abandonados.
O reuso de células é um método simples, porém efetivo para aumentar o potencial reprodutivo e evoluiu em diversos ramos filéticos de Aculeata (Michener, 1964, citado por Sakagami, Gobbi, & Zucchi, 1990).
Em ninhos ativos de vespas sociais cujos favos são expostos, como ocorre com espécies de Polistes, o reuso de células é freqüente. Gobbi & Zucchi (1985) observaram em Ribeirão Preto, SP, que Polistes versicolor Olivier, reutilizando células para a emergência até a 4a geração. Em Rio Claro, SP, Gobbi, Fowler, Chaud Netto e Nazareth (1993) encontraram, através da contagem de mecônios das células, mais de três reutilizações em ninhos de P. versicolor e duas reutilizações em algumas células de ninhos de P. simillimus. Prezoto (2001) e Colas-Rosas, Pinto e Gobbi (2002) também encontraram, em ninhos de P. simillimus, algumas células usadas em três ocasiões. O método da contagem de mecônios foi empregado uma vez que essas duas espécies, ao que se sabe, não removem o mecônio depositado no fundo das células quando da reutilização.
O ninho de P. simillimus é do tipo estelocitaro (com um pedicelo), gimnodomo (células sem o envoltório protetor), construído de polpa de celulose, liso, de cor cinza escuro, e segundo a descrição de um deles, feita por Richards (1978), as células maiores medem 17,0mm de comprimento e 4,5mm de diâmetro.
P. simillimus é uma vespa social encontrada no Brasil desde o Rio Grande do Sul até a Bahia e, também, na Bolívia e no Paraguai; apresenta o tórax e o propodeo em tons ferruginosos claros e ocasionalmente escuros com as pernas e gaster nos mesmos tons e com manchas amarelas sendo que as manchas do gaster em P. simillimus são arredondadas em menor número do que aquelas encontradas em P. versicolor, uma espécie próxima (Richards, 1978).
Os substratos usados para a fixação dos ninhos são as construções de alvenaria, mais comumente os beirais de madeira, luminárias, galhos (observações no Biotério do I.B. UNESP, Rio Claro, SP).
A contribuição de P. simillimus no controle de diversas pragas agrícolas (Prezoto, 1996, 2001; Prezoto, Giannotti, & Machado, 1994; Prezoto & Machado, 1999) é importante.
As abelhas Tetrapediini estão incluídas em dois gêneros: Tetrapedia Klug e Coelioxoides Cresson considerados grupos irmãos (Roig- Alsina, 1990). Coelioxoides são cleptoparasitas e Tetrapedia coletadoras de óleos florais (Alves-dos- Santos, Melo, & Rozen Jr., 2002; Garófalo, (DATA) citado por Silveira, Melo & Almeida, 2002; Roig-Alsina, 1990).
O habitat das espécies desses gêneros são as regiões tropicais das Américas sendo que as espécies de Tetrapedia são mais diversificadas em áreas de cerrado (Zanella, 2002). A fauna de abelhas do cerrado caracteriza-se pela grande representatividade dos gêneros da família Anthophoridae (=Apidae) coletores de óleos, como Centris, Epicharis, Paratetrapedia, Tetrapediae mais Meliponinae (Silveira & Campos, 1995).
Quatro espécies de Coelioxoides (uma não nominada) e oito espécies de Tetrapedia (duas não nominadas) (Pedro & Camargo, 1997) estão. Uma dessas espécies é Tetrapedia diversipes, estudada por Moure (1941) com exemplares coletados em várias regiões do Brasil, nordeste, sudeste e sul, inclusive Rio Claro, SP (Silveira et al, 2002).
Em Cajuru, SP, em trabalho objetivando verificar a abundância e sazonalidade de vespas e abelhas que nidificam em cavidades pré-existentes (ninhos armadilhas utilizando gomos de bambu), T. diversipes teve maior presença em áreas em que existiam construções abandonadas, circundadas por algumas culturas (cana-de-açúcar, milho, arroz e sorgo) e glebas de Matas Mesófica Semidecídua e Galeria (Camillo, Garófalo, Serrano, & Muccillo, 1995). T. diversipes foi, também, uma das espécies mais abundantes encontrada em Guaribas, Paraíba, em área representando um mosaico entre savanas abertas e florestas fechadas (Aguiar & Martins, 2002).
Na busca dos locais de nidificação, espécies de Tetrapedia procuram as cavidades encontradas na madeira (Michener, 2000; Michener, Mc Ginley, & Danforth, 1994; Silveira et al. 2002). nidifica solitariamente e os locais escolhidos são as cavidades na madeira (mesmo as já usadas) e pode nidificar junto a restos de ninhos inativos de outros himenópteros como os da abelha Anthodioctes megachiloides (Megachil T. diversipesidae) Holmberg e a casulos de vespas Trypoxylon (Crabronidae) (Alves-dos-Santos et al, 2002) e também, por indução, em ninhos armadilhas, (Aguiar & Martins, 2002; Alves-dos-Santos et al., 2002; Camillo, et al. 1995).
Pólen e óleos florais são usados como alimento pelas larvas, e a mistura de terra, óleo e, possivelmente secreções, são utilizadas na preparação das partições das células (Alves-dos-Santos et al, 2002).
T. diversipes está relacionada com a coleta de pólen e óleo em Oncidium paranaensis (Oncidiinae), em Curitiba, Paraná (Singer & Cocucci, 1999); em Opuntia (Cactaceae) e Ludwigia(Onagraceae), pólen, NE do Rio Grande do Sul, (Alves-dos-Santos, 1999). Machos de Tetrapedia também coletam óleo nas escopas (Vogel & Machado, 1991, mencionando, possivelmente, T.rugulosa Friese, em flores de Angelonia bisaccata e A. pubescens); Singer e Cocucci, 1999, mencionando T. diversipes, em flores de Oncidium paranaensis.
As flores produtoras de óleos, que são atrativos às abelhas, são encontradas em 80 gêneros de 10 famílias vegetais e nos Trópicos estão representadas, principalmente, nas famílias Malpighiaceae (40 gêneros), Orchidaceae (11 gêneros) e Scrophulariaceae (9 gêneros) (Ramalho & Silva, 2002).
Descrevemos aqui a ocorrência de caso de reutilização, de algumas células de ninho abandonado da vespa social Polistes simillimus, pela abelha solitária Tetrapédia (Tetrapedia) diversipes.
Material e Métodos
O Biotério do Instituto de Biociências da UNESP, Campus de Rio Claro, SP, (22°24S; 47°33W, 612m de altitude) é um prédio retangular de 1100m2 com um jardim interno que contém algumas plantas ornamentais, árvores frutíferas e pequenos canteiros de crucíferas. As temperaturas médias (máx./min.) em janeiro, fevereiro e março de 2003 foram de 24,9°C, 27,0°C, 24,8°C, respectivamente, e o índice pluviométrico de 411,2mm, 113,3mm e 96,9mm, respectivamente, (dados do CEAPLA, IGCE, UNESP, Rio Claro).
Durante um levantamento fenológico em 10 de fevereiro de 2003, no local, foi observado em um ninho abandonado de Polistes simillimusmuitas formigas Crematogaster sp removendo pólen do interior de algumas das células. O ninho estava fixado por um arame na madeira de um beiral e inspeção mais detalhada revelou algumas células fechadas com uma camada escura, compactada, enquanto outras estavam fechadas com barro vermelho. Este ninho havia sido transferido do distrito de Ferraz, distante aproximadamente 10 km, alguns meses antes, e as vespas permaneceram em atividades por poucos dias. Após ter ocorrido o abandono pelas vespas, o ninho foi, então, reutilizado em algumas células pela abelha, e em 10 de fevereiro foi retirado do beiral em que se encontrava e levado para o laboratório.
Formigas e alguns pequenos coleópteros foram removidos do ninho e as células que estavam fechadas com a camada escura e também aquelas que continham o barro vermelho foram separadas das demais células. As células separadas foram colocadas em recipientes de vidro fechados, e mantidos sob condições de temperatura e umidade ambiente, para observações nos dias subseqüentes. Na separação foi coletada uma abelha adulta, viva, escura, não demonstrando agressividade, no interior de uma das células.
Algumas abelhas emergiram, posteriormente, no laboratório e também alguns machos foram coletados, em um agregado, nas proximidades do Biotério.
Exemplares foram enviados ao Prof. Dr. João M. F. Camargo e Profa. Dra. Silvia Regina de Menezes Pedro, da USP, Ribeirão Preto, SP, que os identificaram como Tetrapedia (Tetrapedia) diversipes Klug, 1810.
Os demais exemplares coletados no trabalho serão depositados na Coleção de Espécies do Departamento de Zoologia do Instituto de Biociências, UNESP, Rio Claro, São Paulo.
Resultados
O ninho continha 860 células e destas, três continham uma camada escura de terra com resina ou óleo fechando externamente as células e cinco apresentavam apenas a parte mediana fechada com o mesmo material, ficando visível o pólen que estava sendo armazenado na parte anterior, antes do ninho ter sido coletado.
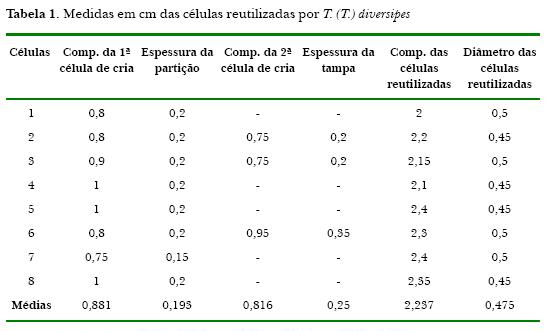
Encontramos para as oito células reutilizadas no ninho de P. simillimus o comprimento médio de 2,237cm (Tabela 1). Uma célula reutilizada no ninho de P. simillimus pode ser vista (Fig. 1), na qual existe uma partição de terra e óleo a qual secciona a célula: a parte posterior da célula (a qual denominamos de 1a célula de cria) apresentou o comprimento médio de 0,881cm (o espaço disponível após a partição até atingir a parte posterior em que se localizava o mecônio), e a anterior (a 2a célula de cria) o comprimento médio de 0,816cm (o espaço disponível após o fechamento da entrada da célula reutilizada até atingir a partição). Apenas três células reutilizadas estavam totalmente fechadas na parte anterior e continham duas células de crias, e cinco das células reutilizadas continham apenas a 1ª célula, ficando aberta à parte anterior. O total de células de crias de T. (T.) diversipes encontradas foi onze.

Das três células reutilizadas que continham as duas células de crias em uma a tampa (a camada que fechava a entrada da célula) estava rente à entrada, e nas outras duas, um pouco mais para o interior.
O diâmetro das células reutilizadas (a entrada hexagonal das células que adquire uma forma mais arredondada quando da reutilização) que encontramos foi de 0,475cm. A espessura média da tampa que continha as duas células de crias foi de 0,25cm e a espessura média da partição que dividia as duas células de crias foi de 0,193cm.
Três das células reutilizadas continham no fundo o mecônio de P. simillimus e em quatro células a análise não pode ser conclusiva devido o fundo já estar bastante danificados, pelas formigas ou coleópteros, ou ambos; e uma das células não continha o mecônio. Orifícios menores de 2mm e outros, um pouco maiores, foram observados também nas laterais das células.
Emergiram no laboratório três fêmeas e um imaturo foi encontrado em uma das células; dois machos foram coletados próximos ao local, em um agregado.
Discussão
Alves-dos-Santos et al. (2002) verificaram que os ninhos de T. diversipes consistiam de uma a sete células por ninho, em ninhos armadilhas (normalmente cinco a sete células em cavidades de 10-12cm de comprimento, linearmente dispostas) e com o comprimento de cada célula variando de 10-15mm. T. (T.) diversipes (Fig. 2) nidificou em cavidades de comprimentos menores (2,0- 2,4cm) nas células do ninho de P. simillimus (embora estas sendo um pouco maiores do que aquelas mencionadas por Richards (1978), que foi de 17mm), adaptou uma ou duas células de crias por célula reutilizada e repetiu o procedimento em outras células vazias, vizinhas. Com a ressalva do pequeno número de células reutilizáveis encontradas, neste caso, a abelha desenvolveu sua prole em espaços, comparativamente pequenos (0,816 e 0,88cm, médias). O comprimento médio das abelhas fêmeas, emergidas em cativeiro (n=3), em vista lateral, foi de 7,52mm e do macho coletado, 8,5mm. O tempo de desenvolvimento das abelhas é influenciado pela temperatura, e indivíduos menores tendem a ser produzidos em temperaturas mais altas (Garófalo, 1985; Kamm, 1974; Roubik, 1989; Tepedino & Parker, 1986), o que pode estar relacionado com a temperatura constatada no local em fevereiro de 2003 (média de 27,0?C). Acreditamos que a escolha por T. (T.) diversipesde cavidades de comprimento reduzidos necessita de mais investigações.

O diâmetro das células que encontramos (0,475cm) foi, também, comparativamente menor do que os utilizados por T. diversipes em 70% dos ninhos armadilhas observados por Aguiar e Martins, (2002) que foi de 0,6cm e os observados por Alves-dos-Santos et al. (2002) para Tetrapedia, em São Paulo, variaram de 3-10mm com possível preferência por aqueles com 4- 6mm.
A interferência das predadoras formigas Crematogaster pode ser considerada como um nítido obstáculo no desenvolvimento de T. (T.) diversipes, tanto na predação do pólen como das suas formas imaturas. Alguns coleópteros encontrados (menores de 5mm), inclusive no interior das células de crias, podem ter interferido no desenvolvimento da prole de T. (T.) diversipes, porém, não foi possível inferir a maneira como ocorreu e as suas conseqüências.
Células parcialmente revestidas com barro vermelho em seus interiores foram encontradas vizinhas àquelas reutilizadas por T. (T.) diversipes indicando que outra reutilizadora também deu início na adaptação de células no ninho abandonado. Este comportamento não é incomum em ninhos de vespídeos sociais e, no mesmo local, foi constatado o reuso de células de ninho abandonado de Mischocyttarus Saussure (Hymenoptera: Vespoidea, Polistinae, Mischocyttarini) por Pison duckei Menke (1968) (Hymenoptera: Crabronidae) (Pinto, Fowler, Digiampietri, & Gobbi, 2002) bem como foi observado exemplares de Pachodynerus sp (Hymenoptera: Vespidae, Eumeninae) emergidos de ninho antigo de P. simillimus, (Nozor Paulo Outeiro Pinto, observação pessoal, abril de 2003).
Abelhas e vespas solitárias têm afinidades em seus comportamentos de nidificação (Camillo et al., 1995; Jayasingh & Freeman, 1980), ocorrendo similaridades no emprego de materiais usados para as construções, ou adaptações, dos ninhos (como o uso de barro, fibras vegetais, folhas, resinas) bem como a escolha dos locais em que nidificarão, quer sejam sob o solo ou em substratos acima do solo.
Na maioria das espécies de Sphecidae (Hymenoptera; Apoidea) o comportamento de nidificação é solitário e os ninhos destas vespas são colocados em duas categorias dependendo da locação: 1) aqueles construídos sob o solo (terrícolas) e, 2) e os construídos acima do solo. No segundo grupo estão incluídos os ninhos feitos nos ramos, os desenvolvidos nas fendas, e os de barro, encontrados junto às construções feitas pelo homem. Nem todas estas vespas, porém, constroem um buraco original e aquelas que fazem os ninhos nos ramos podem tomar túneis abandonados já usados por abelhas, bem como podem adotar e remodelar ninhos de barro (Bohart & Menke, 1976). Nos Eumeninae (Hymenoptera: Vespoidea) na maior parte das espécies, o comportamento de nidificação também é solitário e algumas espécies podem escavar seus ninhos no solo, vertical ou horizontalmente, e a maioria nidifica em cavidades pré-existentes, como buracos em troncos, orifícios feitos e usados por besouros, ninhos abandonados de outros himenópteros e mesmo conchas vazias de caracol (Carpenter & Marques, 2001).
Espécies de abelhas que nidificam solitariamente e incluídas nas famílias Apidae, Anthophoridae (=Apidae), Megachilidae, Halictidae, Colletidae, exibem, também, uma grande diversidade quanto aos locais escolhidos para nidificar, quer sejam em substratos sólidos (buracos abertos no solo, ninhos de térmitas, ninhos de formigas, ninhos expostos, os feitosnos galhos e nas folhagens) bem como em cavidades preexistentes (buracos em árvores, tocas de besouros, covas, subterrâneos, orifícios e ninhos usados geralmente por outros himenópteros) (Roubik, 1989).
No contexto ecológico, um aspecto positivo do reuso de células para espécies de vespas e abelhas solitárias, é o da diminuição dos custos advindos dos fatores tempo, esforço, risco de predação, oferecendo, por conseguinte, uma possibilidade maior de sucesso no potencial reprodutivo à espécie reutilizadora.
A Apidae T. (T.) diversipes parece demonstrar, quanto a escolha dos locais para desenvolver sua prole, a plasticidade no comportamento de nidificação observado nas espécies de vespas e abelhas solitárias, não aquelas construtoras de um específico padrão de ninho, e sim, as (re)utilizadoras e adaptadoras de diversificadas cavidades preexistentes, buscando, inclusive, as encontradas nas células de ninho abandonado da vespa social P. simillimus.
Referências
Aguiar, A. J. C., & Martins, C. F. (2002). Abelhas e vespas solitárias em ninhos-armadilha na Reserva Biológica Guaribas (Mamanguape, Paraíba, Brasil). Revista Brasileira de Zoologia, 19(Supl. 1), 101-116. [ Links ]
Alves-dos-Santos, I. (1999). Abelhas e plantas melíferas da mata atlântica, restinga e dunas do litoral norte do Estado do Rio Grande do Sul, Brasil. Revista Brasileira de Entomologia, 43(3/4), 191-223. [ Links ]
Alves-dos-Santos, I. (2002). A vida de uma abelha. Ciência Hoje, 30(179), 60-62. [ Links ]
Alves-dos-Santos, I., Melo, A. R., & Rozen Jr, J. C. (2002). Biology and immature stages of the bee Tribe Tetrapediini (Hymenoptera: Apidae). American Museum Novitates, 3377, 1-45. [ Links ]
Ayasse, M., Paxton, R. J., & Tengö, J. (2001). Mating behavior and chemical communication in order Hymenoptera. Annual Review of Entomology, 46, 31-78. [ Links ]
Bohard, R. M., & Menke A. S. (1976). Sphecid wasps of the world. A generic revision. Berkeley, CA: University of California Press. [ Links ]
Buchmann, S. L. (1987). The ecology of oil flowers and their bees. Annual Review of Ecology and Systematics, 18, 343-69. [ Links ]
Camillo, E., Garófalo, C. A., Serrano, J. C., & Muccillo, G. (1995). Diversidade e abundância sazonal de abelhas e vespas solitárias em ninhos armadilhas (Hymenoptera, Apocrita, Aculeata). Revista Brasileira de Entomologia, 39(2), 459-470. [ Links ]
Carpenter, J. M., & Marques, O. M. (2001). Contribuição ao estudo dos vespídeos do Brasil (Insecta, Hymenoptera, Vespoidea, Vespidae). Dissertação de Mestrado, Departamento de Fitotecnia, Escola de Agronomia, Universidade Federal da Bahia, Cruz das Almas, BA. [CD-ROM]. Série Publicações Digitais, 2. [ Links ]
Colas-Rosas, P. F., Pinto, N. P. O., & Gobbi, N. (2002). Comparação da produtividade em ninhos de tamanho médio com um ninho de tamanho acima da média em vespas Polistes simillimus Zikán (1951). Em Livro de Resumos. 5 Simpósio de Iniciação Científica (p. 12). UNESP, Rio Claro, SP. [ Links ]
Cowan, D. P. (1991). The solitary and presocial vespidae. Em K. G. Ross & R. W. Matthews (Eds.), The social biology of wasps (pp. 33-73). Ithaca, NY: Comstock Publishing Associates, Cornell University Press. [ Links ]
Evans, H. E., & Eberhard, M. J. W. (1970). The wasps. Ann Arbor, MI: University of Michigan Press. [ Links ]
Garófalo, C. A. (1985). Social structure of Euglossa cordata nests (Hymenoptera: Apidae: Euglossini). Entomology General, 11, 77-83. [ Links ]
Gobbi, N., & Zucchi, R. (1985). On the ecology of Polistes versicolor versicolor (Olivier) in southern Brazil. 2. Colony productivity. Naturalia, 10, 21-25. [ Links ]
Gobbi. N., Fowler, H. G., Chaud Netto, J., & Nazareth, S. L. (1993). Comparative colony productivity of Polistes simillimus and the evolution of paragyny in the Polistinae. Zool. J. B. Physiol, 97, 239-242. [ Links ]
Jayasingh, D. B., & Freeman, B. E. (1980). The comparative population dynamics of eight solitary bees and wasps (Aculeata; Apocrita; Hymenoptera) trap-nested in Jamaica. Biotropica, 12(3), 214-219. [ Links ]
Kamm, D. (1974). Effects of temperature, day length, and number of adults on the sizes of cells and offspring in a primitively social bee (Hymenoptera: Halictidae). Journal of the Kansas Entomology Society, 47, 8-18. [ Links ]
Michener, C. D. (1974). The social behavior of the bee – A comparative study. Harvard: MA: Belknap Press of Harvard University Press. [ Links ]
Michener, C. D., Mc Ginley, R. J., & Danforth, B. N. (1994). The bee genera of north and Central America. Washington, DC: Smithsonian Institution Press. [ Links ]
Michener, C. D. (2000). The bees of the world. Baltimore: Johns Hopkins. [ Links ]
Moure, Pe. J. S. (1941). Notas sobre abelhas do grupo Tetrapedia. Revista de Entomologia, 12, 515-521. [ Links ]
Pedro, S. R. de M., & Camargo, J. M. F. de (1997). Biodiversidade do Estado de São Paulo, Brasil. Em C. A. Joly & C. E. Bicudo (Orgs.). Apoidea Apiformes (Vol. 5, pp. 194-211). São Paulo: FAPESP. [ Links ]
Pinto, N. P. O., Fowler, H. G., Digiampietri Filho, E. A., & Gobbi, N. (2002). Notes on three species of Pison Jurine, 1808 (Hymenoptera: Crabronidae) collected in Rio Claro (SP), Brazil. Arquivos do Instituto Biológico, 69(1), 89-91. http://w.w.w.biologico.sp.gov.br/arquivos/V69-1/pinto.pdf [ Links ]
Prezoto, F., Giannotti, E., & Machado, V. L. L. (1994). Atividade forrageadora e material coletado pela vespa social Polistes simillimus Zikán, 1951 (Hymenoptera, Vespidae). Insecta, 3(1), 11-19. [ Links ]
Prezoto, F. (1996). Ação de Polistes (Aphanilopterus) simillimus Zikán, 1951 (Hymenoptera, Vespidae) no combate às pragas de Zea mays L. Dissertação de Mestrado, Instituto de Biociências, Universidade Estadual Paulista, UNESP, Rio Claro, SP. [ Links ]
Prezoto, F., & Machado, V. L. L. (1999). Ação de Polistes (Aphanilopterus) simillimus Zikán (Hymenoptera, Vespidae) no controle de Spodoptera frugiperda (Smith) (Lepidoptera, Noctuidae). Revista Brasileira de Zoologia, 16(3), 841-850. [ Links ]
Prezoto, F. (2001). Estudos biológicos e etológicos dePolistes (Aphanilopterus) simillimus Zikán, 1951. Tese de Doutorado, Instituto de Biociências, Universidade Estadual Paulista, UNESP, Rio Claro, SP. [ Links ]
Ramalho, M., & Silva, M. (2002). Flora oleífera e sua guilda de abelhas em uma comunidade de retinga tropical. Sitientibus série Ciências Biológicas 2(1/2), 34-43. <http://andromeda.uefs.br/dcbio/> [ Links ]
Richards, O. W. (1978). The social wasps of the Americas excluding the Vespinae. British Museum (Natural History). Norwich, England: Fletcher & Son. [ Links ]
Roig-Alsina, A. (1990). Coelioxoides Cresson, a parasitic genus of Tetrapediini (Hymenoptera: Apoidea). Journal of the Kansas Entomological Society. 63, 279- 287. [ Links ]
Roubik, D. W. (1989). Ecology and natural history of tropical bees. Cambridge: Cambridge University Press. [ Links ]
Silveira, F. A., & Campos, M. J. O. (1995). A melissofauna de Corumbataí (SP) e Paraopeba (MG) e uma análise da biogeografia das abelhas do cerrado brasileiro (Hymenoptera, Apoidea). Revista Brasileira de Entomologia, 39(2), 371-401. [ Links ]
Silveira, F. A., Melo, G. A. R., & Almeida, E. A. B. (2002). Abelhas brasileiras, sistemática e identificação. Belo Horizonte, MG: Ministério do Meio Ambiente, PROBIO-PNUD, Fundação Araucária. [ Links ]
Singer, R. B., & Cocucci, A. A. (1999). Pollination mechanisms in four sympatric southern Brazilian Epidendroideae orchids. Lindleyana, 14(1), 47-56. [ Links ]
Sakagami, S. F., Gobbi, N., & Zucchi, R. (1990). Nesting biology of a quasisocial Sphecid Wasps Trypoxylon fabricator. Japanese Journal of Entomology,58(4), 846-862. [ Links ]
Spradbery, J. P. (1973). An account of the biology and natural history of solitary and social wasps. Seatle: University of Washington Press. [ Links ]
Tepedino, V. J., & Parker, F. D. (1986). Effect of rearing temperature on mortality, secondgeneration emergence, and size of adult in Megachile rotundata (Hymenoptera: Megachilidae). Journal of Economic Entomology, 79, 974-977. [ Links ]
Vogel, S., & Machado, I. C. (1991). Pollination of four sympatric species of Angelonia (Scrophulariaceae) by oil-colleting bees in NE Brazil. Plant, Systematics and Evolution, 178, 153-178. [ Links ]
Zanella, F. C. V. (2002). The bees of caatinga (Hymenoptera, Apoidea, Apiformes): A species list and comparative notes regarding their distribution. Apidologie, 31(5), 579-592. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Nozor Paulo Outeiro Pinto
Departamento de Ecologia, Universidade Estadual Paulista
Av. 24A, 1515, Bela Vista,
13506-900, Rio Claro, SP, Brasil
E-mail: npopinto@rc.unesp.br
Recebido em 7 de dezembro de 2005
Aceito em: 5 de março de 2006
1Agradeço ao Prof. Dr. João M. F. Camargo e à Profa. Dra. Silvia Regina de Menezes Pedro, da F.F.C.L., USP, Ribeirão Preto, SP, pela identificação dos exemplares de Tetrapedia (Tetrapedia) diversipes Klug, 1810. À Profa. Dra. Luceli de Souza, pelos comentários críticos do texto e sugestões. Ao Sr. Luis Fernando Stocco, pelo trabalho fotográfico. Aos estudantes e técnicos de laboratório dos Departamentos de Ecologia e de Zoologia da UnB. Este trabalho foi apresentado resumidamente nos Anais do II Simpósio de Ecologia Comportamental e de Interações. Apoio financeiro: Universidade de Brasília.

