











 (pdf)
(pdf)




 Permalink
PermalinkRevista de Etologia
ISSN 1517-2805
Rev. etol. vol.9 no.2 São Paulo dez. 2010
ARTIGO
Comportamentos de hierarquia e dominância em grupos de bugios-ruivos, Alouatta guariba clamitans (Cabrera, 1940), em cativeiro
Hierarchy and dominance behaviors in brown howler monkey groups Alouatta guariba clamitans (Cabrera, 1940) in captivity
Alice Copetti DalmasoI; Thais Leiroz CodenottiII
I Universidade Federal de Santa Maria
II Associação para Conservação da Vida Silvestre
RESUMO
O estudo que segue foi realizado com dois grupos de bugios-ruivos (Alouatta guariba clamitans) em cativeiro, no Centro de Acolhimento de Primatas e Aves (PRIMAVES) com o objetivo de identificar comportamentos de hierarquia e dominância, através da quantificação e duração das condutas, comparando os dados obtidos com dados já descritos para o gênero Alouatta. Foram realizadas 172 horas de observação e a partir da quantificação das condutas catar (categoria social não-agonística), vocalizar (categoria social agonística), vigiar (categoria alerta) e comer (categoria alimentação). Ambos os grupos mostraram uma evidente dominância entre os indivíduos, com os machos juvenis apresentando certo grau de centralidade espacial, demonstrado através dos comportamentos de vocalização e vigiar. As fêmeas mostraram-se ativas nas funções de vigia e catação, evidenciando o seu importante papel na contribuição da estabilidade social, proporcionando ao grupo que vive em cativeiro uma relação apaziguadora entre os indivíduos.
Palavras-chave: Bugio-ruivo. Alouatta guariba clamitans. Hierarquia. Dominância.
ABSTRACT
This study was done with two groups of brown howler monkeys in captivity, at the Centro de Acolhimento de Primatas e Aves (PRIMAVES). The aim was to identify hierarchy and dominance behaviors through quantification and duration of conducts, comparing the data with information already described for Alouatta genus. A total of 172 hours of observations were carried out, and from the quantification of grooming (non-agonistic social category), roaring (agonistic social category), vigilance (alert category) and eating (feeding category), both groups showed an evident dominance between the individuals, with the youthful males performing a certain degree of spatial centralization, showed through roaring and vigilance behaviors. The females are active in vigilance and grooming, showing an important role in the maintenance of social stability, providing to the group that lives in captivity an appeasing relation between the individuals.
Keywords: Brown howler monkey. Alouatta guariba clamitans. Hierarchy. Dominance.
Os animais exercem padrões de comportamento que são "uma mistura inseparável de componentes herdados ou específicos da espécie e, de adquiridos ou aprendidos" (Kindlovits, 1999, p. 203) que por sua vez contribuem, diretamente, no sucesso reprodutivo e na sobrevivência dos indivíduos (Krebs & Davies, 1996). Em adição a isso, Wilson (1975, apud Jones, 1980) afirma que hierarquias de dominância são características em muitas sociedades de animais e são praticamente universais em indivíduos que vivem em grupo, entre eles, os primatas. Por conseguinte, a vida em grupo e a presença de uma hierarquia e dominância em bandos de bugios facilitam, na vida in situ, a diminuição da predação, maior eficiência em encontrar alimento, além de troca de informações, acesso a parceiros para acasalamento, termorregulação e, inclusive, transmissão de doenças ( Marques, 2001).
Em estudo de dominância em grupos de primatas, Jones (1981) afirma que o comportamento agressivo em bugios pela busca de recursos, associado com o tamanho do corpo e força, gerou a dominância de ranking mais alto de hierarquia, composto por indivíduos adultos jovens, sendo estes, portanto, mais capacitados à competição que indivíduos mais velhos. Esse tipo de organização social, no qual a idade está correlacionada negativamente com o ranking de hierarquia para ambos os sexos, é encontrado em Alouatta palliata (Jones, 1979, 1980). Porém, na maioria dos outros grupos de primatas, machos são dominantes sobre fêmeas e um sistema linear de hierarquia é evidente (Jones, 1983; Oliveira & Ades, 1998; Gruener, Machado & Hirano, 2000; Silveira & Codenotti, 2001; Wang & Milton, 2003; Domingo- Balcells, 2005).
Em trabalhos recentes, Alouatta guariba clamitans recebeu atenção quanto à ecologia populacional (Jardim, 2005; Miranda et al., 2006; Miranda & Passos, 2005), ao comportamento de beber (Almeida-Silva et al., 2005; Miranda et al., 2005), ao comportamento social (Miranda et al., 2005; Silveira & Codenotti, 2001), à dieta (Miranda & Passos, 2004; Silveira & Codenotti, 2001a), à funcionalidade das chamadas de longo alcance (Cunha & Jalles-Filho, 2007; Steinmetz, 2005), à morfologia do aparelho reprodutor feminino (Veras, Miglino & Silva, 2006), ao comportamento antipredatório (Miranda et al., 2006), ao registro de simpatria entre A caraya e A. guariba clamitans (Codenotti et al., 2002; Aguiar et al., 2006), à verificação anormal da coloração de pelagem em população isolada (Fortes & Bicca-Marques, 2007), e, ainda, ao estudo molecular de localização cromossômica de sequência telomérica (Mudry, Nieves & Bolzán, 2007).
Por ser descrita como uma espécie em risco de extinção em médio prazo de tempo pela IUCN Red List of Threatened Species (2007), e como vulnerável na Lista das Espécies da Fauna Ameaçadas de Extinção no Rio Grande do Sul (2002), o presente estudo com A. guariba clamitans em cativeiro teve por finalidade contribuir com o bem-estar e a conservação dos indivíduos nesse meio, considerado uma das alternativas para conservação da espécie. A pesquisa realizada teve por objetivo identificar comportamentos de hierarquia e dominância em grupos de bugios-ruivos jovens e infantes, em cativeiro, comparando os resultados com dados comportamentais já descritos na literatura, para o gênero Alouatta.
Método
Área de estudo
A pesquisa foi desenvolvida no Criadouro Conservacionista "Centro de Acolhimento de Primatas e Aves" (PRIMAVES), (28° 10' 7'' S; 52° 32' 9'' W; elevação: 638 m), no distrito de Bela Vista, município de Passo Fundo, Rio Grande do Sul, Brasil.
Procedimentos
Entre março de 2007 e janeiro de 2008 foram estudados dois grupos de bugios-ruivos com as seguintes composições: G1: três fêmeas infantes, dois machos infantes e um macho juvenil; G2: dois machos juvenis e duas fêmeas juvenis, sendo que ambos os grupos não possuem indivíduos aparentados. Os bandos foram observados em quatro etapas bimestrais de 10 dias cada, com duas observações diárias por grupo, uma pela manhã e outra à tarde (das 8h às 11h30 e das 14h às 18h30 ), com duração de 1 hora para cada observação (4 horas de observação diárias). O método de conduta (Martin & Bateson, 1991) foi utilizado para favorecer a quantificação das frequências (acrescentando-se as durações) no aparecimento dos comportamentos de hierarquia e dominância e as interações sociais entre os membros dos grupos, e o método ad libitum como ferramenta qualitativa, auxiliando na observação, identificação e descrição comportamental. Os comportamentos observados e analisados foram: catar (categoria social nãoagonística), vocalizar (categoria social agonística), vigiar (categoria alerta) e comer (categoria alimentação).
Para os registros de interações, os indivíduos foram identificados por nome e para as análises de hierarquia e dominância, os mesmos foram caracterizados por categoria sexoetária, tamanho corporal e coloração.
Os dados quantitativos foram somados e analisados por dia de observação, através do teste nãoparamétrico de Friedman, pela aplicação do Qui- Quadrado (para as interações sociais) e da estatística descritiva (média e desvio padrão).
Resultados
Obeteve-se um registro de 160 observações, totalizando 172 horas, com exatamente 86 horas de registro para cada grupo. O grupo G2, constituído por quatro indivíduos jovens (Raí, Raísa, Bebel e Greg) realizou várias interações sociais, apresentando diferenças estatisticamente significativas, demonstradas pelo teste Qui-quadrado, com respeito ao comportamento Catar. (Tabela 1).
Tabela 1. Valores significativos do X2 para a conduta catar entre os membros do grupo G2 (p< 0,05).
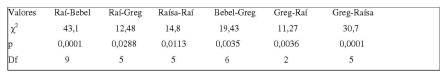
Para esta conduta, o macho juvenil Raí (de maior tamanho que Greg e coloração avermelhada) foi o que mais realizou a ação (f= 56), e o Greg o que menos catou (f= 17). Entre as duas fêmeas juvenis, ambas realizaram a ação igualmente (f= 48); porém, Raísa, mais jovem que Bebel, foi a mais catada (f= 44). Houve diferenças de catação entre as duas fêmeas e o macho dominante, com Raí realizando mais a ação com Raísa, catando-a numa frequência (f) de 32 interações e ela catando-o 19 vezes. O Teste X2 , porém, não obteve valores estatisticamente significativos para essas interações, mostrando apenas que ele não catou as duas fêmeas de forma igual.
Aplicando o Teste Qui-quadrado, os resultados não foram estatisticamente significativos para o comportamento de interação alimentar nesse grupo, que teve anotada a frequência com que um indivíduo roubava o alimento diretamente das mãos ou da boca de outro indivíduo. Analisados os atos comportamentais Vigiar (alerta) e Vocalizar (social não-agonístico), o Teste de Friedman mostrou diferenças de conjunto altamente significativas para as frequências (X2corr.= 97,04; p= 0,0001; X2corr.= 33,0; p= 0,0001, respectivamente) e igualmente para a durações (X2corr.= 91,52; p= 0,0001; X2corr.= 34,2; p= 0,0001, respectivamente). A tabela 2 mostra os dados obtidos pela aplicação da estatística descritiva.
Tabela 2. Frequência e duração média dos comportamentos realizados por cada indivíduo do grupo G2 (Vocalizar e Vigiar).
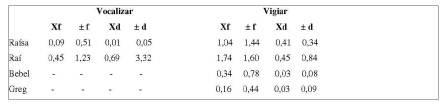
No comportamento Vocalizar, Raí apresentou a maior frequência (f= 36) e, igualmente para a conduta Vigiar, foi o indivíduo que mais realizou a ação (f= 134), seguido de Raísa (f= 84), Bebel (f= 27) e Greg, com a menor frequência de vigia (f= 13).
No grupo G1, composto por seis indivíduos (Yago, Drica, Jully, Jack, Jasper e Cindy), o Teste Quiquadrado apresentou diferenças significativas com relação às interações do comportamento catar (tabela 3).
Tabela 3. Valores significativos do X2 para a conduta catar entre os membros do grupo (p< 0,05).
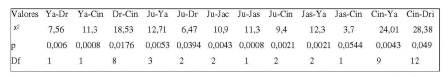
Para a interação alimentar, o teste não apresentou diferenças estatisticamente significativas. Neste grupo, em todas as etapas de observação, a frequência das condutas Vocalizar (f= 18) e Vigiar (f= 24) foram realizadas somente pelo macho juvenil Yago (Xfvo= 2,00; ±f= 2,26; Xdvo= 0,94; ±d= 2,08; Xfal= 0,59; ±f= 0,71; Xda= 0,50; ±d= 1,25).
Discussão
A conduta grooming (catar) foi relatada na literatura por Wilson (1975, apud Jones, 1979) como um comportamento apaziguador de indivíduos num grupo, que além de reduzir a probabilidade de agressão entre eles, pode ser interpretado como um comportamento altruístico descrito por Seyfarth (1977, apud Schino, 2007), no qual indivíduos que catam outros indivíduos de alto ranking de hierarquia seriam retribuídos pelo beneficio de conduta em troca de apoio agonístico. Esta conduta não foi aplicável neste caso para o grupo G2, pois o indivíduo mais velho do grupo, considerado previamente o indivíduo dominante (Raí), foi quem mais procurou realizar a ação, e o macho submisso (Greg) foi o que menos realizou a conduta no grupo. Não seguindo, portanto, os padrões comportamentais do sexo com relação à catação. Nossa análise corroborou com os resultados de Oliveira e Ades (1998) e de Silveira e Codenotti (2001) com relação às fêmeas do grupo terem sido as maiores realizadoras da conduta. A única hipótese para explicar a alta reciprocidade de catação entre o macho Raí e a fêmea mais jovem Raísa é que cresceram juntos, desde os primeiros meses de vida, e mantiveram sempre um estreito laço de amizade e uma grande afinidade. Raísa só teve filhote na última etapa do estudo, em janeiro de 2008, não havendo procura maior por parte do macho em realizar ou receber a ação entre as etapas (o nascimento do infante de Raísa em janeiro de 2008 fez com que o macho Raí expulsasse Greg do grupo, comprovando ao observador a dominância deste indivíduo e o comportamento próprio da espécie). Com relação à fêmea Bebel, mesmo havendo uma baixa frequência de interações entre os dois (Raí-Bebel: f= 14; Bebel-Raí: f= 17), percebeu-se que quando a fêmea ganhou um filhote em agosto de 2007, o macho procurou aproximar-se da mesma para receber a catação.
A presença de infantes num dos períodos do estudo fez com que houvesse variações nas taxas de relacionamento afiliativo entre Raísa e Bebel, pois o filhote desta promoveu uma aproximação e cuidado maior de Raísa em relação à fêmea mais velha e ao infante, resultados estes similares ao estudo de Zucker e Clarke (1998) com fêmeas de Alouatta palliata. Ambas as fêmeas cataram igualmente; porém, a fêmea sexualmente madura (Bebel) foi a que menos sofreu a ação entre elas. Diante desses resultados e do fato de que o comportamento social catar deixa patente a hierarquia existente entre os membros do grupo (Silveira & Codenotti, 2001), consideramos que a fêmea Raísa ocupa posição mais elevada que a fêmea Bebel, compartilhando, portanto, com os resultados obtidos por Jones (1983) com fêmeas de A. caraya. Entretanto, as baixas taxas do comportamento social catar neste grupo comprovam o que Southwick (1963, apud Jones, 1979) afirma sobre bugios, primatas de não contato.
Em relação à vocalização de bugios, Wilson (1975, apud Jones, 1998) afirma que a comunicação destes animais seria primariamente vocal, uso de sinal e de comunicação não danosa. Além disso, é consistente afirmar que primatas não humanos, entre eles os do gênero Alouatta, emitem as altas chamadas por sons de baixa frequência para facilitar sua transmissão por longas distâncias (Mitani & Stuht, 1998), permitir comunicação intragrupal (Steinmetz, 2005), ter relação com encontros intergrupais (Oliveira & Ades, 1996), defesa ativa de bordas (Cunha & Jalles- Filho, 2007) e regulação do uso do espaço (Cunha & Birne, 2006).
Raí (G2) foi identificado como o macho alfa pelo seu maior número de atividade de vocalização, comparado com os outros indivíduos do bando, que pouco ou nada realizaram esse comportamento (Tabela 2). As vocalizações não se intensificaram nos períodos em que havia presença de infantes, ao contrário dos resultados de Kitchen (2004), em que os machos alfas de A. pigra respondiam mais rapidamente e por mais tempo na presença de filhotes nos grupos. Este resultado pode ser explicado pelo fato de que o cativeiro apresenta movimentação de funcionários e, principalmente, pelo comportamento dos outros grupos de primatas, entre eles, bugios que ficam expostos à visão dos animais em seus recintos. Por isso, a taxa de frequência e duração de vocalização para bugios cativos depende, principalmente, do comportamento de vocalização dos bugios de outros grupos do cativeiro, de possíveis interferências (como ruídos externos e visitantes) que o macho pode considerar uma iminente ameaça, e ainda de mudanças no tempo, como previsão de chuva (Horwich & Gebhard, 1983). Diante do exposto, contudo, é necessário ressaltar que Raí, mesmo identificado como o dominante do grupo é um macho jovem, por isso não tendo desenvolvido por completo seu potencial efetivo de macho alfa, o qual zela pela segurança do grupo.
Machos subadultos e fêmeas tendem a participar do coro de vocalização. Porém, a única que participou, embora pouco significativamente, foi Raísa, a fêmea mais jovem do bando, que o fez provavelmente como suporte na vocalização do macho alfa do grupo. Embora não tenham sido analisados separadamente dos rugidos, os latidos (barks), de acordo com Oliveira (2002, apud Oliveira & Ades, 2004), tiveram a clara função de intensificar contextos de alarme. Com isso, fica claro que a vocalização não se torna um comportamento de dominância entre os indivíduos, mas exalta o tipo de hierarquia existente num bando.
Acreditou-se, no início, que a conduta Comer (categoria alimentação) fosse, de alguma maneira, mostrar a relação de dominância entre os indivíduos através do comportamento agonístico entre eles enquanto comiam ou pela prioridade de acesso aos recursos alimentares, como fez Wang e Milton (2003) para identificação do macho alfa em A. palliata. A ação de roubar alimento do outro não nos mostrou resultados estatisticamente significativos, porém, a partir da observação ad libitum da prioridade de acesso ao alimento, levando em conta qual dos indivíduos se posicionava primeiro sobre a bandeja, e qual o último a se aproximar da mesma, encontraram-se resultados de relevância. Entre as fêmeas não houve diferença quanto a se colocar primeiro sobre a bandeja. Porém, ambas, na maioria das vezes, esperavam o macho alfa (Raí) se aproximar, pegar o alimento (a melhor porção e de melhor qualidade disponível) e retirar-se, para então se aproximar e comer. Entre os machos, Greg, além de ser geralmente o primeiro a se posicionar sobre a bandeja, não aparentava temor na presença do macho Raí, além de não ter sido presenciada conduta agonística entre os dois.
A categoria de Alerta (conduta vigiar), que dentre as condutas analisadas foi a mais realizada no grupo e por todos os indivíduos, para primatas arbóreos é definida na literatura como qualquer "correr dos olhos" direcionado para além do alcance dos braços do animal focal, excluindo olhadelas para o próprio corpo do animal, um parceiro de grooming, substrato próximo ou algum item alimentar" (Treves, Drescher & Ingrisano, 2001). Para espécies maiores de primatas como os do gênero Pan, a vigilância é descrita como um comportamento antipredatório, relacionado à evitação da predação (Stanford, 2002). As médias de frequência e de duração foram maiores para Raí, o que reforça e comprova, efetivamente, ser este o macho alfa do grupo G2. Depois do macho alfa, as fêmeas tiveram a segunda maior taxa de vigilância, com o macho subordinado (Greg) logo em seguida. O aumento da vigilância depois do nascimento do primeiro infante corrobora os dados de Treves, Drescher e Nicky (2001) com grupos de A. pigra. Isso esclarece ao observador que a presença de filhotes pode ser um dos principais fatores a provocar o comportamento de alerta no bando.
Com relação à análise do grupo G1, observouse uma alta taxa na frequência de catação (f= 189), e mesmo tratando-se de um grupo de jovens e infantes, porém estável, as fêmeas permaneceram as maiores realizadoras da conduta de catação (Jully, Cindy e Drica), sendo Drica quem mais procurou realizá-la (f= 86). Yago, o macho jovem do grupo, foi o mais catado (f= 45), seguido de Jully, a menor fêmea do grupo (f= 44), ação realizada principalmente por Drica. Porém, Cindy (segunda maior realizadora da conduta, com f= 54) e Yago foram dois indivíduos que freqüentemente realizavam outras condutas de interação social comuns entre todos os infantes, como brincar e encostar. Além disso, nas vocalizações de Yago, Cindy procurava sempre se colocar ao seu lado, eriçando o pelo (postura não observada nos outros indivíduos). Ou seja, ao contrário da literatura, que mostra que infantes de Alouatta não são ativos, e sim, passivos à conduta catar observada em vida livre (Oliveira & Ades, 1998; Silveira & Codenotti, 2001), em cativeiro, esse comportamento foi evidente entre os infantes, que influenciaram na coesão e estabilidade social do grupo.
A partir dos resultados obtidos, através da execução das categorias vocalização e alerta, comprovou-se que o macho juvenil mais velho (Yago), de maior tamanho corporal e de coloração ruiva, é o alfa do grupo, exercendo também dominância sobre os demais machos e fêmeas, que sendo mais jovens que os indivíduos do G2, iniciavam timidamente seus comportamentos de vocalização e vigia, sem disputa alguma com o dominante. Mesmo em um ambiente de cativeiro, onde, a priori, os animais não são aparentados, mas possuem vínculo afetivo entre si, estabelece-se uma hierarquia semelhante a bandos de bugios de vida livre, em que os juvenis exercem certo grau de dominância sobre indivíduos infantes.
Comparando os dados de frequência e duração média das condutas Vocalizar e Vigiar entre os machos dominantes dos grupos, Raí (G2) apresentou valor maior (Tabela 2) que Yago (G1) apenas para a frequência média do comportamento vigiar, fato que ocorreu provavelmente porque Raí realizava a ação somente quando se aproximava do chão ou da bandeja de alimento, que são comportamentos rápidos. Ficou nítida, para o observador, a iminente reação de Yago a ruídos externos ao recinto, que o importunaram (como barulho de máquinas, por exemplo), o que talvez explique valores maiores de freqüência e duração média de vocalização e de duração média de vigiar para o mesmo. Raí vocalizava, na maioria das vezes, somente em resposta ao coro dos outros grupos de bugios dos demais recintos ou na presença do observador.
Portanto, constatou-se que a dominância existe em ambos os grupos, tanto no grupo G2, estritamente juvenil, com Raí como o macho dominante e Raísa ocupando a posição de destaque entre as fêmeas, quanto no grupo G1, tipicamente infantojuvenil, com Yago em posição de evidente influência hierárquica sobre os demais indivíduos.
As condutas vocalizar e vigiar são comportamentos que nos permitem visualizar a hierarquia existente nos bandos, ao contrário da conduta social de catação e de interação alimentar que nos revelam a relação de dominância entre os indivíduos. Hierarquia e dominância agem juntas na estrutura e estabilidade social de bandos de bugioruivo.
Agradecimentos
Agradecemos à equipe de funcionários do Primaves pela colaboração; ao biólogo, Responsável Técnico do Centro, Luizandro Ferrari e aos seus familiares, pela preocupação e auxílio na hospedagem e alimentação da executora do projeto; à profa. dra. Nêmora Pauletti Prestes, pela revisão do manuscrito e pelas importantes contribuições científicas conferidas ao texto.
Referências
Aguiar, L. M. D., Mellek, D. M., Abreu, K. C., Boscarato, T. G., Bernardi, I. P., Miranda, J. M. D., & Passos, F. C. (2007). Sympatry between Alouatta caraya and Alouatta guariba clamitans and the rediscovery of free-ranging potential hybrids in Southern Brazil. Primates, 48. Recuperado em 18 ago. 2008: http://zoo.bio.ufpr.br/mamifero/sympatry.pdf [ Links ]
Almeida-Silva, B., Guedes, P. G., Boubli, J. P., & Strier, K. B. (2005). Deslocamento terrestre e o comportamento de beber em um grupo de barbados (Alouatta guariba clamitans) em Minas Gerais, Brasil. Neotropical Primates, 13(1), 1-3. [ Links ]
Balcells, C. D., Luna, E. R., Aliaga, M. E., Mávil, J. M., & Morales, M. M. (2005). Social interactions and dominance hierarchy in a captive group of mantled howler monkeys (Alouatta palliata mexiacana). Folia Primatologica, 76, 45-66. [ Links ]
Codenotti, T. L., Martins da Silva, V., Albuquerque, V. J., Camargo, E.W., & Martins Silveira, R.M. (2002). Distribuição e Situação atual de Conservação de Alouatta caraya (Humboldt, 1812) no Rio Grande do Sul. Neotropical Primates, 10(2), 132-141. [ Links ]
Cunha, R. G. T., & Jalles-Filho, E. (2007). The roaring of southern brown howler monkeys (Alouatta guariba clamitans) as a mechanism of active defence of borders. Folia Primatologica, 78, 259-271. [ Links ]
Marques, A. A. B. (2001). Técnicas para estudos ecológicos e comportamentos de primatas. V Congresso Brasileiro de Ecologia. [ Links ]
Marques, A. A. B., Fontana, C. S., Velez, E., Bencke, G. A., Schneider, M., & Reis, Dos Reis, R. E. (Org). (2002). Lista de Referência da Fauna Ameaçada de Extinção no Rio Grande do Sul. Decreto nº 41.672, Porto Alegre: FZB/MCT–PUCRS/PANGEA, 52p. (Publicações Avulsas FZB, 11).
Fortes, V. B., & Bicca-Marques J. C. (2008). Abnormal Pelage Color in an Isolated Population of Alouatta guariba clamitans Cabrera, 1940 in South Brazil. International Journal Primatology, 29, 717-722. [ Links ]
Gruener, C. G., Machado, G. S., & Hirano, Z. M. B. (2000). Análise da estruturação hierárquica de um grupo com três espécimes de Alouatta fusca clamitans (Primates: Atelidae), submetidos ao regime de cativeito. Em Resumos – Hierarquia de dominância (p. 18).
Horwich, R. H., & Gebhard, K. (1983). Roaring rhythms in black howler monkeys (Alouatta pigra) of Belize. Primates, 24(2), 290-296. [ Links ]
Jardim, A. M. A. (2005). Ecologia populacional de bugiosruivos (Alouatta guariba) nos municípios de Posto Alegre e Viamão, RS, Brasil. Tese de doutorado, Universidade Estadual de Campinas, Campinas, SP. [ Links ]
Jones, C. B. (1979). Grooming in the mantled howler monkey, Alouatta palliata Gray. Primates, 20(2), 289- 292. [ Links ]
Jones, C. B. (1980). The functions of status in the mantled howler monkey, Alouatta palliata Gray: intraspecific competition for group membership in a folivorous neotropical primate. Primates, 21(3), 389-405. [ Links ]
Jones, C. B. (1981). The evolution and socioecology of dominance in primate groups: a theoretical formulation classification and assessment. Primates, 22(1), 70-83. [ Links ]
Jones, C. B. (1983). Social organization of captive black howler monkeys (Alouatta caraya): "social competition" and the use of non-damaging behavior. Primates, 24(1), 25-39. [ Links ]
Jones, C. J. (1998). A broad-band contact call by female mantled howler monkeys: implications for heterogeneous conditions. Neotropical Primates, 6(2), 38-40. [ Links ]
Kindlovits, A. (1999). Clínica e terapêutica em primatas neotropicais. Juiz de Fora, MG: EDUFJF. [ Links ]
Kitchen, D. M. (2002). Alpha male black howler monkey responses to loud calls: effect of numeric odds, male companion behaviour and reproductive investment. Animal Behaviour, 67, 125-139. [ Links ]
Krebs, J. R., & Davies, N. B. (1996). Introdução à ecologia comportamental (M. Ramalho & C. P. Machado, trads). São Paulo: Atheneu. [ Links ]
Martin, P., & Bateson, P. (1991). Measuring Behaviour: Un introductory guide. Versão espanhola. [ Links ]
Miranda, J. M. D., & Passos, F. C. (2004). Hábito alimentar de Alouatta guariba (Humboldt) (Primate, Atelidae) em Floresta de Araucária, Paraná, Brasil. Revista Brasileira de Zoologia, 21(4), 821-826. [ Links ]
Miranda, J. M. D., & Passos, F. C. (2005). Composição e dinâmica de grupos de Alouatta guariba clamitans Cabrera (Primates, Atelidae) em Floresta Ombrófila Mista no Estado do Paraná, Brasil. Revista Brasileira de Zoologia, 22(1), 99-106. [ Links ]
Miranda, J. M. D., Moro-Rios, R. F., Bernardi, I. P. & Passos, F. C. (2005). Formas não-usuais para a obtenção de água por Alouatta guariba clamitans em ambiente de Floresta com Araucária no sul do Brasil. Neotropical Primates, 13(2), 21-23. [ Links ]
Miranda, J. M. D., Aguiar, L. M. D., Ludwig, G., Moro-Rios, R. F., & Passos, F. C. (2005). The first seven months of an infant of Alouatta guariba clamitans (Humboldt) (Primates, Atelidae): interactions and the development of behavioral patterns. Revista de Zoologia, 22(4), 1191- 1195. [ Links ]
Miranda, J. M. D., Bernardi, I. P., Moro-Rios, R. F., & Passos, F. C. (2006). Three years on demography of a group of Alouatta guariba clamitans Cabrera (Primates, Atelidae): growth and fragmentation. Revista de Zoologia, 23(3), 703-706. [ Links ]
Miranda, J. M. D., Bernardi, I. P., Moro-Rios, R. F., & Passos, F. C. (2006). Antipredator behavior of brown howlers attacked by black hawk-eagle in southern Brazil. International Journal of Primatology, 27(4), 1097-1101. [ Links ]
Mitani, J. C., & Stuht, J. (1998). The evolution of nonhuman primate loud calls: acoustic adaptation for long-distance transmission. Primates, 39(2), 171-182. [ Links ]
Mudry, M. D., Nieves, M., & Bolzán, A. D. (2007). Chromosomal localization of the telomeric (TTAGGG)n sequence in eight species of New World Primates (Neotropical Primates, Platyrrhini). Cytogenet Genome Research, 119, 221-224. [ Links ]
Oliveira, D. A. G., & Ades, C. (1996). Vocalizações de longo alcance e de alarme do bugio ruivo Alouatta fusca clamitans no Parque Estadual da Cantareira (São Paulo- SP). Em Resumos. XIV Encontro Anual de Etologia. Uberlândia, MG. [ Links ]
Oliveira, D. A. G., & Ades, C. (1998). Proximity and grooming interactions as indicators of the social organizations of brown howling monkeys (Alouatta fusca clamitans). Neotropical Primates, 6(4), 115-117. [ Links ]
Oliveira, D.A.G., & Ades, C. (2004). Long-distance calls in Neotropical primates. Anais da Academia Brasileira de Ciências, 76(2). Recuperado em mar. 2006, da SciELO (Scientific Electronic Library Online): http://www.scielo.br [ Links ]
Santos, E. O. (2005). Metabolismo do estresse: impactos na saúde e na produção animal. Seminário apresentado. Programa de Pós-Graduação em Ciências Veterinárias, UFRGS. Recuperado em 02 set. 2008: http://www6.ufrgs.br/bioquimica/posgrad/BTA/estresse.p df [ Links ]
Schino, G. (2007) Grooming and agonistic support: a metaanalysis of primate reciprocal altruism. Behavioral Ecology, 18, 115-120. [ Links ]
Silveira, M. M. R., & Codenotti, T. L. (2001). Interações Sociais e dieta de Bugio-ruivo, Alouatta guariba clamitans, no Parque Estadual de Itapuã, Rio Grande do Sul, Brasil. Neotropical Primates, 9(1), 15-19. [ Links ]
Steinmetz, S. (2005). Vocalizações de longo-alcance como comunicação intra-grupal nos bugios (Alouatta guariba). Neotropical Primates, 13(2), 11-15. [ Links ]
Treves, A., Drescher, A., & Nicky, I. (2001). Vigilance and aggregation in black howler monkeys (Alouatta pigra). Behaviour Ecol. Sociobiology, 50, 90-95. [ Links ]
Veras M. M., Miglino, M. A., & Silva, Z. (2007). Morfologia do aparelho reprodutor em fêmeas de bugio marrom (Alouatta guariba clamitans). Braz. J. vet. Res. Anim.. 44(1), 12-17. [ Links ]
Zucker, E. L., & Clarke, M. R. (1998). Agonistic and affiliative relationships of adult female howlers (Alouatta palliata) in Costa Rica over a 4-year period. International Journal of Primatology, 19(3), 433-449. [ Links ]
Wang, E., & Milton, K. (2003). Intragroup social relationships of male Alouatta palliata on Barro Colorado Island, Republic of Panama. International Journal of Primatology, 24(6), 1227-1243. [ Links ]
Recebido em: 01/03/2009
Aceito em: 01/02/2011

Alice Copetti Dalmaso
Universidade Federal de Santa Maria
e-mail: alicedalmaso@gmail.com
Thais Leiroz Codenotti
Associação para Conservação da Vida Silvestre.
