










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTIIndicadores
Compartilhar

 Permalink
PermalinkTemas em Psicologia
versão impressa ISSN 1413-389X
Temas psicol. vol.5 no.3 Ribeirão Preto dez. 1997
PROCESSOS SOCIAIS E DESENVOLVIMENTO
Uma aproximação às vocalizações do muriqui (Brachyteles arachnoides)(1)
César Ades; Francisco Dyonísio Cardoso Mendes
Departamento de Psicologia Experimental USP
O comportamento de comunicação dos primatas oferece informações de especial importância sobre os seus processos cognitivos e sobre sua organização social. Estudos bem conhecidos (Premack 1983; Gardner e Gardner, 1994; Gardner, Garder & Van Cantfort, 1989; Savage-Rumbaugh 1990; entre outros) nos prepararam a aceitar a idéia de que primatas, pelo menos chimpanzés, são capazes de adquirir o uso flexível e semântico de um sistema arbitrário de sinais. Permanece a questão de saber quão complexa, flexível e ajustada às circunstâncias é a comunicação em condições naturais, quando os primatas vivem por sua própria conta, expostos aos perigos e pressionados pela necessidade de obter seu alimento. Não há contradição numa proposta que una a preocupação pelo cognitivo e pela adaptação ao habitat natural. Os processos de percepção, memória, raciocínio etc. ganham em ser tomados numa perspectiva psicoetológica, isto é, levando-se em conta os contextos naturais em desempenham uma função adaptativa (Ades, 1993).
Os estudos sobre comunicação em primatas concentraram-se, inicialmente, a partir da obra de Darwin (1872), na análise de expressões faciais, gestos e posturas. Maestripieri (1996), num estudo que pertence a esta tradição, mostrou recentemente que, no caso de Macaca nemestrina, expressões como o "pucker" (uma forma de comprimir e esticar os lábios para a frente, com retraimento das sobrancelhas e das orelhas), o estalar da língua, a exibição dos dentes, comunicam estados emocionais e intenções, sinalizam posições de dominância ou de submissão e constituem um contexto para a redução da distância social entre os indivíduos. Especialmente interessante é o uso do pucker em intercâmbios entre animais: o pucker de um indivíduo é freqüentemente seguido de puckers de outros indivíduos, como se estivesse ocorrendo uma "conversa".
A partir do final da década de 1970, com o progresso nas técnicas de registro e análise sonográfíca das vocalizações, ganhou impulso o estudo da comunicação vocal. Tornou-se possível obter uma representação bastante precisa das características acústicas dos sons produzidos pelos animais, um passo fundamental para sua análise em termos de notas ou elementos (que seriam como os fonemas, na análise lingüística da fala humana), e para o estudo de uma possível "gramática", ou seja, das dependências seqüenciais destes elementos ou grupos de elementos e das regras de sua produção.
Também despertaram entusiasmo resultados que indicavam que, além de seus componentes emocionais ou motivacionais, a comunicação vocal dos primatas possuía características "semânticas". O macaco vervet (Cercopithecus aethiops) tem, em seu repertório vocal, chamados de alarme distintos para predadores aéreos como a águia; predadores terrestres como o leopardo; ou para cobras. Numa pesquisa clássica, em que usaram reproduções em playback, essenciais como controle sobre a informação efetivamente transmitida, Seyfarth, Cheney e Marler (1980) demonstraram que vervets adultos respondiam de maneira diferenciada diante dos chamados de águia, onça ou cobra mesmo que não pudessem ver o predador ou o comportamento do emissor. Após um chamado de leopardo, os membros do grupo procuravam a extremidade de galhos em árvores vizinhas; após um chamado de águia, refugiavam-se em arbustos terrestres e, após o chamado da cobra, permaneciam alertas, sobre as patas traseiras, inspecionando o chão. Cheney e Seyfarth (1990) levantam a possibilidade de que estes chamados possam ser usados de modo enganador, como quando, no meio de uma luta entre dois bandos de vervets, um dos membros do grupo perdedor lança um grito de leopardo, dispersando todos e permitindo uma recuperação de vantagem.
Dentre as muitas funções que podem ser atendidas através da vocalização intragrupal está a de indicar a presença de fontes de alimento ou de alimentos abundantes (Chapman e Lefebre, 1990; Dittus, 1984), a de sinalizar o início e direção do movimento grupai (Boinski, 1993), a de promover ou manter laços afiliativos entre os indivíduos (Smith, Newman e Symmes, 1982), e a de regular o espaçamento entre os indivíduos durante as suas atividades de forrageio (Mendes e Ades, 1995; Palombit, 1992; Robinson, 1982).
O etólogo que se propõe entender o significado dos chamados de uma determinada espécie encontra-se numa situação análoga à imaginada - e usada filosoficamente - por Quine (1960), a do antropólogo que visita uma certa cultura e tenta traduzir as palavras dessa cultura sem a ajuda de um intérprete. Uma tradução radical (o termo é de Quine) nos parece menos árdua no caso da passagem de uma língua para outra, do que no da interpretação de vocalizações. Os aspectos transculturais do comportamento humano, as constâncias no uso de símbolos lingüísticos (acreditar nestas constâncias não significa necessáriamente aderir à posição chomskyana) fornecem pistas para o antropólogo/tradutor. No caso dos chamados de primatas, lançados na floresta, a incerteza é maior: como decifrar o significado de seqüências sonoras sem antes entender o modo de vida do animal, sua reação aos predadores e às presas ou ao alimento em geral, suas interações afiliativas e agonísticas, os rituais de cortejo e acasalamento, e outros comportamentos ainda? Além do estudo contextual, é preciso definir critérios, os mais "naturais"possíveis, para o recorte dos registros sonográficos em suas sílabas ou elementos básicos. Como lembram Nowicki e Nelson (1990) "não se pode, tão facilmente quanto no caso do estudo de pessoas, colocar perguntas aos animais para verificar nossas opiniões a respeito do que constitui uma categoria natural" (p. 90). Não há um critério definitivo para a categorização das unidades vocais. Nowicki e Nelson citam (e comparam) três: uma ordenação multidimensional, baseada em correlações de espectrogramas; a aplicação de técnicas de conglomerados e uma forma mais intuitiva que consiste no julgamento, por observadores humanos, dos sonogramas, a partir de suas características visuais. E interessante que este último critério não tenha se revelado inferior aos outros dois.
Um guia possível, na tentativa de tradução, pode ser encontrado nas abordagens da lingüística e da psicolingüística. Consiste em rejeitar a dicotomia "entre sistemas de comunicação simples, estereotipados e fixos nos animais e sistemas de comunicação complexos, variáveis e abertos no ser humano" (Snowdon, 1982, p. 211). Snowdon tem sido o defensor de uma abordagem à comunicação vocal em primatas que leve em conta sua variabilidade, complexidade e modulação por aprendizagem social e que se apoie - heurísticamente ou mesmo em nível de modelos - no estudo da linguagem. A percepção categórica de certos sinais (os animais receptores reagem a faixas de um estímulo que varia gradualmente como se fossem categorias distintas), a variabilidade de indivíduo para indivíduo (havendo, muitas vezes, a presença de uma "assinatura vocal", ou seja, de aspectos da vocalização de um indivíduo discrimináveis por outros), os dialetos que surgem em regiões geograficamente distintas, mudanças ontogenéticas, os indícios de sintaxe etc. são todos aspectos em que o paralelismo com a fala humana pode ser propício como fonte de hipóteses.
No presente artigo, para exemplificar as estratégias e os resultados possíveis de uma análise da vocalização em ambiente natural, expomos alguns dos resultados de um programa de pesquisa sobre a vocalização do muriqui (Brachyteles arachnoides), em curso em nosso laboratório de comunicação acústica, na USP. Karen Strier, pioneira das pesquisas sobre o comportamento do muriqui, notou uma vez que era "difícil saber o que os muriquis dizem uns aos outros, e o que entendem " (1992, p. 79). Nossa pesquisa é uma tentativa de saber o que os muriquis dizem uns aos outros, uma aproximação ao muriquês.
O MURIQUI
O muriqui ou mono-carvoeiro, o maior primata neotropical (pode alcançar 15 kilos de peso), é endêmico da Mata Atlântica e se apresenta em duas sub-espécies, Brachyteles arachnoides arachnoides, encontrada nos estados do Rio e de São Paulo, e Brachyteles arachnoides hypoxanthus, que se localiza em Minas Gerais e no Espírito Santo. Dotado de uma cauda preênsil que lhe proporciona um quinto ponto de apoio (além das mãos), ele se desloca com agilidade pelos galhos a uma boa altura do chão. Percorre a floresta em busca de frutos, mas também está especializado em alimentar-se de folhas, principalmente nas épocas em que os frutos estão escassos.
O grupo de muriquis compõe-se de vários machos e fêmeas adultas, de jovens e infantes, podendo alcançar mais de 50 indivíduos. As fêmeas não se distinguem dos machos, pelo tamanho do corpo ou dos caninos, não existe a dominância dos machos típica de outros primatas que vivem em grupos multimacho-multifêmea. Mesmo assim, notam-se diferenças: os muriquis machos são mais coesos espacialmente, interagem mais entre si de forma afíliativa (Mendes, 1990). Isso talvez se origine no fato de serem as fêmeas, nesta espécie, as que migram para outros grupos, como parte da estratégia reprodutiva; os machos permanecem no grupo e sua coesão talvez tenha a ver com o parentesco.
Há muito pouco conflito ou agressão aberta dentro do grupo de muriquis. Em anos de pesquisa de campo, foram poucos os episódios observados que pudessem inequivocamente ser classificados como agonísticos. Os relacionamentos sociais não seguem o modelo hierárquico de dominância/subordinação tão claro em outras espécies, os machos não monopolizam sítios de dormida, fontes de alimento ou mesmo acesso às fêmeas; a tolerância em relação aos juvenis é generalizada. Se há hierarquia, entre os muriquis, é uma hierarquia afíliativa, que pode ser constatada registrando-se a taxa de abraços e toques e o número de vezes que cada indivíduo se senta perto de outros (Mendes, 1990). O agonismo aparece nos confrontos entre grupos, marcado por fortes vocalizações, por comportamentos de aproximação e de apoio nos membros de cada grupo. E ritualizado e sem corpo a corpo.
A busca das fontes de alimento é uma atividade primordial, na qual o muriqui gasta boa parte de seu tempo de vigília. Começando mais cedo no verão, mais tarde no inverno, interrompida por pausas que são maiores no meio do dia, o forrageamento se dá geralmente através de uma locomoção do grupo como um todo, sem coordenação rígida, os indivíduos avançando em formações variáveis, dispersos na mata, muitas vezes sem contato visual. Em grandes árvores, quando é abundante a oferta, o grupo todo ou quase todo pode ser visto se alimentando, ou acampado nas proximidades (Strier, 1992). Para que esta estratégia de coleta de alimento seja eficiente, ela deve satisfazer duas condições: a primeira é que os indivíduos possam dispersar-se o suficiente para a localização e o uso de fontes também dispersas; a segunda é que possa ser mantida uma direção coletiva, com coordenação do deslocamento dos indivíduos e com a possibilidade de reunião, após afastamentos temporários. A vocalização permite que estas características do forrageio possam ser implementadas, aumenta a distância pela qual cada membro ou subgrupo de membros pode se afastar por conta própria, sem perigo de perder-se em relação ao resto do grupo e propicia a reunificação, quando necessário. Numa mata densa, a informação sobre quem está vocalizando e onde é essencial.
O REPERTÓRIO DE VOCALIZAÇÕES DO MURIQUI
Os muriquis dispõem de um sistema muito rico de vocalizações. Muitas destas ocorrem em contextos específicos, e são produzidas por categorias específicas de indivíduos, sendo relativamente mais fácil a interpretação de sua função. Foi possível descrever latidos, gritos e bafejos de alarme; chamados usados durante episódios de brincadeira; trinados e guinchos de filhotes; piados e trinados de mães de infantes; trinados, gritos e guinchos de fêmeas receptivas; gargarejos, grunhidos, guinchos e bramidos de ameaça, e diversas formas de intercâmbios antifonais, ou seja, produções de sons em que dois ou mais indivíduos se alternam na emissão de sinais de estrutura relativamente semelhante (Mendes, no prelo).
Focalizaremos aqui um tipo muito especial e muito freqüente de vocalização dos muriquis, que se caracteriza pela ocorrência de chamados compostos de relinchos e gritos, emitidos em sucessão por indivíduos distintos. Tipicamente, um indivíduo começa vocalizando e, em menos de 10 segundos, um outro membro do grupo, situado em local próximo ou distante, vocaliza por sua vez, e depois outro e assim por diante, como se cada um respondesse, indicando talvez sua identidade e localização. Chamamos este tipo de interação vocal, geralmente emitido por adultos e subadultos mais velhos, intercâmbio seqüencial.
Assim se refere Strier (1992) aos chamados de intercâmbio seqüencial:
"Seqüências de relinchos são sempre produzidas quando os muriquis acordam, como se cada qual anunciasse a sua presença e a sua localização. À medida em que aumenta o número de indivíduos ativos, os relinchos aumentam em freqüência, parecendo-se cada mais com uma conversa. Discussões semelhantes podem ser ouvidas logo antes de os animais pousarem para uma sesta, à tarde; e à noitinha quando alcançaram o local onde passarão a noite" (p. 79).
A PESQUISA NA ESTAÇÃO BIOLÓGICA DE CARATINGA
A pesquisa sobre a qual se baseia nosso artigo foi realizada na Estação Biológica de Caratinga, município de Montes Claros, Minas Gerais. Trata-se de um fragmento de Mata Atlântica, de aproximadamente 900 hectares, circundada por plantações, com um relevo formado de morros e vales pelos quais o observador tem de se locomover, seguindo trilhas, atrás dos muriquis que dispõem de um meio de locomoção bem mais ágil. Os dados foram obtidos com um dos grupos que vivem em Caratinga, o grupo do Matão. Os membros deste grupo - observado originalmente por Strier (1986) e seguido desde então por várias equipes de pesquisadores - foram todos identificados individualmente, recebendo nomes: Arlene, Bess, Cher, Mona etc, as fêmeas; Cutlip, Irv, Preto, Clyde, Sony etc, os machos. Todos estavam habituados à presença humana.
A coleta sistemática ocorreu entre setembro de 1990 e agosto de 1991. Empregou-se o método animal focal, ou animal focal de registro contínuo (Altmann, 1974) para a obtenção de amostras dos chamados de membros adultos do grupo. Seguia-se cada animal amostrado por 10 minutos, mantendose o equipamento de gravação ligado em permanência, e o microfone na direção do animal focal. Garantia-se, assim, o registro de todas as vocalizações, inclusive as curtas e imprevisíveis. Gravações oportunísticas (ad libitum) foram usadas para a obtenção de sinais vocais raros, e das categorias utilizadas por indivíduos imaturos. Os registros de vocalizações foram efetuados com um gravador Marantz PMD-430 e um microfone cardióide direcional Sennheiser ME88 + módulo K3N. As vocalizações foram editadas e analisadas através de um sistema digital Signal/RTS de tempo contínuo.
Um registro dos dados contextuais era mantido incluindo, entre outras coisas: (1) a atividade e espaçamento do grupo (forrageio numa única fonte ou durante mudança de sítio, grupo espalhado, acampados, inativos), (2) o número total de chamados no intercâmbio (um intercâmbio era considerado completo depois de um intervalo de 10 segundos posterior à última vocalização); (3) a distância máxima e mínima dos outros participantes no intercâmbio em relação ao emissor (0 a 20 metros; 20 a 50 metros; 50 a 100 metros, mais de 100 metros); (4) a presença de possíveis referentes externos (predadores em potencial, membros de outro grupo de muriquis, grandes fontes de alimento); (5) comportamentos sociais e não-sociais do emissor; (6) sua posição em relação ao resto do grupo (isolado, periférico etc.). Não sendo possível, de antemão, saber que aspectos do contexto serão relevantes para entender a função comunicativa dos chamados, impõe-se manter muito ampla a gama dos registros contextuais.
OS ELEMENTOS DA VOCALIZAÇÃO
Os chamados que compõem os intercâmbios seqüenciais têm uma variabilidade considerável. A Figura 1 mostra o registro sonográfíco de vocalizações emitidas por quatro indivíduos, no mesmo intercâmbio. Nota-se que elas diferem na duração, na amplitude máxima, nas notas que os compõem e na combinação destas. Estamos muito longe da estereotipia e da simplicidade estrutural de outros sistemas de comunicação. A combinação de notas e a intensidade de certos chamados os tornam semelhantes a relinchos de potros; outros chamados soam como os gritos roucos de outras espécies de animais. A variabilidade estrutural não se manifesta apenas nas diferenças entre indivíduos, os registros mostram que um mesmo indivíduo pode gerar vocalizações heterogêneas.
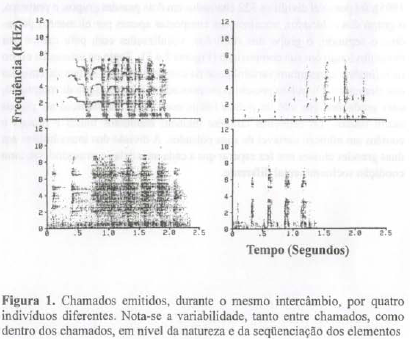
Numa primeira etapa, em que foram usados 322 dentre os 400 primeiros chamados registrados (foram descartados 78 chamados devido à presença de ruídos intensos), procuramos isolar as unidades mínimas de vocalização, os elementos (numa abordagem lingüística, corresponderiam a fonemas). Esta análise preliminar permitiu distinguir 14 elementos básicos. Cinco dentre eles são elementos pulsados (curtos) com duração inferior a 100 milisegundos: p, t, h,r, tri; nove, elementos longos: ra, gm, ar, al, nh, zz, zm, zx, e gt.
A extração de elementos, a partir dos registros acústicos, permite uma análise em termos de teoria da informação, uma descrição quantitativa da redundância e das dependências seqüenciais a partir de análises markovianas. Pode-se chegar perto de especificar a "sintaxe" de produção dos sons elementares, ou seja, a ordem preferencial em que são emitidos, as regras de passagem de uns para os outros.
ESTACADOS E RELINCHOS
A partir da inspeção visual e de uma análise de conglomerados (Mendes, 1995), foi possível dividir os 322 chamados em dois grandes grupos, o primeiro, o grupo dos estacados, vocalizações compostas apenas por elementos pulsados; o segundo, o grupo dos relinchos, vocalizações com pelo menos um elemento longo em sua composição (Figuras 2 e 3). Tanto os estacados como os relinchos apresentam variabilidade na composição e na ordem de emissão dos elementos. Os pulsos presentes em estacados variam no grau de rouquidão, com gradações que vão de pulsos tonais com claros harmônicos até pulsos muito roucos. Os relinchos também incluem elementos tonais e roucos e contêm um número variável de sons pulsados. A divisão dos intercâmbios em duas grandes classes nos fez esperar que a cada uma delas correspondesse uma condição socioambiental diferente.
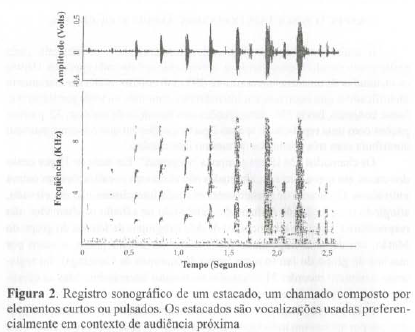
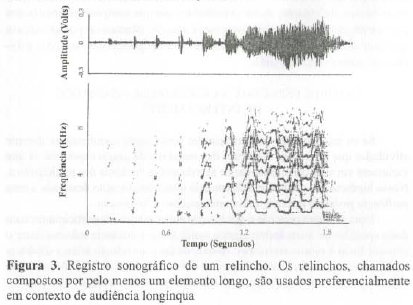
ASPECTOS SOCIAIS DO INTERCÂMBIO SEQÜENCIAL
É muito interessante notar que, nos intercâmbios seqüenciais, cada participante vocaliza, quase sempre, apenas uma vez em cada episódio. Dentre os chamados da amostra, encontramos 293 cujo emissor podia ser claramente identificado e que ocorriam em intercâmbios com dois ou mais participantes. Neste conjunto, havia 255 participações sem repetição de emissor, 32 participações com uma repetição, e apenas 2 participações em que o mesmo muriqui contribuía com três chamados no mesmo intercâmbio.
Os chamados não levam sempre a "respostas". Em mais de 25 por cento dos casos, em nossa amostra, chamados não eliciavam vocalizações em outros indivíduos. O número de participantes em cada intercâmbio não era elevado, atingindo uma média de 4 chamados (incluindo no cálculo os chamados não respondidos). Durante um encontro de dois subgrupos de fêmeas do grupo do Matão, um deles acompanhado por machos do mesmo grupo, o outro por machos do grupo do Jaó (o outro grupo de muriquis de Caratinga), foi registrado o número recorde: 31 chamados no mesmo intercâmbio. Mas as condições eram então muito especiais.
Pode parecer inapropriado incluir, nos intercâmbios, vocalizações produzidas por apenas um indivíduo. As características acústicas destes chamados os tornavam, no entanto, muito semelhantes aos que compunham seqüências com vários participantes. A não-resposta é uma das alternativas plausíveis para qualquer sinal de comunicação; palavras também podem não despertar palavras em outros seres humanos.
FATORES ESPACIAIS NA EMISSÃO DE CHAMADOS DE INTERCÂMBIO
Se os intercâmbios seqüenciais têm uma função coordenadora durante atividades que envolvam dispersão dos membros do grupo esperar-se-ia que variassem em suas características de acordo com a distância dos participantes. Nossa hipótese era de que se diferenciaria uma comunicação destinada a uma audiência próxima da destinada a uma audiência longínqua.
Uma primeira previsão é que haveria maior número de participantes num dado episódio de intercâmbio quanto maior fosse a distância máxima entre o emissor focal e outros emissores. Houve, de fato, correlação entre a distância máxima estimada (em quatro categorias: de 0 a 20 metros, de 20 a 50 metros, de 50 a 100 metros e mais de 100 metros) e o número de chamados do intercâmbio.
Uma segunda previsão partia da constatação de que estacados (chamados compostos apenas por elementos pulsados) são em geral menos intensos do que os relinchos (chamados que contêm elementos longos). Aqueles seriam então mais apropriados para uma comunicação de perto, estes, para uma comunicação de longe. Os dados foram coerentes com a hipótese: foi maior a freqüência de estacados em intercâmbios com um número pequeno de participantes (portanto, próximos) do que em intercâmbios com número maior de participantes. Também diferiu significativamente a distribuição dos estacados e dos relinchos de acordo com a distância estimada. Os primeiros ocorriam principalmente entre 0 e 50 metros de distância máxima, os segundos, a distâncias máximas maiores do que 50 metros.
Sinais acústicos graves são, em geral, menos suscetíveis à difração e à absorção causadas por uma vegetação densa. Representam uma das maneiras de ser minimizada a atenuação excessiva do som em matas tropicais e aumentar o alcance da vocalização. Cinco relinchos graves, em nossa amostra, foram todos emitidos quando os muriquis se encontravam subdivididos e distantes. Dois deles foram emitidos por Bess e Fernanda quando, de volta de uma excursão a uma fonte de frutos longínqua, encontraram o grupo disperso e reagiram ao chamado de uma fêmea não identificada situada a mais de 100 metros.
Como explicar, a partir da hipótese de dois subsistemas de comunicação, que estacados tenham também ocorrido em intercâmbios de longo alcance ? Por que dispender energia em sons altos em interações de curto alcance ? Poder-seia argumentar que, pelo menos em parte, certos chamados em intercâmbios reflitam estados motivacionais dos emissores, não tendo a "intencionalidade" social suposta. Poder-se-ia também pensar, e esta alternativa nos parece razoável, que os vocalizadores produzem sons direcionados a ouvintes potenciais em distâncias diferentes.
RELINCHOS DE FÊMEAS RECEPTIVAS
Três tipos de relinchos foram produzidos quase exclusivamente por fêmeas receptivas: relinchos formados apenas por longas emissões tonais, relinchos formados pela repetição de um elemento modulado com alguma rouquidão e relinchos que incluíam uma sucessão de pulsos rápidos, parecendo trinados. Estes chamados possuem, muito provavelmente, uma função de "divulgação" do estado reprodutivo da fêmeas.
VOCALIZAÇÕES LIGADAS À PRESENÇA DE ALIMENTO
Muriquis na floresta são muitas vezes surpreendidos produzindo relinchos ao chegarem a fontes importantes de alimentação e estes relinchos são muitas vezes seguidos da aproximação de outros indivíduos do grupo. José Rimoli (1994), durante sua pesquisa de mestrado, observou o grupo do Matão chegando na cercania de duas árvores de canafístula (Cássia leiandra) passando perto da árvore, sem que nenhum animal se alimentasse. No final deste mesmo dia e no dia seguinte, os animais efetuaram uma longa volta, retornando à canafístula. No momento da entrada dos primeiros animais na árvore, houve fortes relinchos que talvez desempenhassem uma função de food call agregando o resto dos animais.
Maurício Talebi-Gomes (comunicação pessoal) também verificou, durante sua pesquisa de mestrado com muriquis, no Parque Estadual de Carlos Botelho, vocalizações ligadas à descoberta de alimento. Curiosamente, notou vocalizações semelhantes em muriquis mantidos em cativeiro, no Centro de Primatologia do Rio de Janeiro, de manhã cedo, quando era trazido o alimento. Aos relinchos emitidos num dos recintos respondiam muriquis situados em recinto distante.
ALGUMAS IMPLICAÇÕES TEÓRICAS
A tradução - mesmo que não radical (Quine, 1960) - das vocalizações de um animal como o muriqui é um empreendimento que custa horas de permanência no campo, horas de decodifícação de registros magnéticos e muito esforço de análise, tanto para a definição de elementos, para a elaboração de modelos mais abrangentes e para a descoberta dos determinantes contextuais. Não se obtém, não é este o objetivo, um dicionário de sílabas ou frases em muriquês, em que cada categoria de vocalização se liga a um "significado" distinto, mas sim uma compreensão do papel que diversos tipos de chamados desempenham na economia comportamental do grupo de animais. O que instiga e inspira é verificar a complexidade dos sistemas de comunicação do muriqui. De um lado, a variabilidade impressionante dos modos de combinação de elementos; de outro, o caráter socioespacial e sociocognitivo dos chamados seqüenciais.
Entre os atributos mais relevantes, do ponto de vista de uma comparação entre os sistemas de comunicação vocal de primatas e a linguagem humana, está o uso de regras gramaticais e sintáticas. Várias aves e primatas - e o muriqui se enquadra evidentemente nesta categoria - geram vocalizações diferentes por meio da recombinação de elementos acústicos recorrentes. Será que estão em jogo regras generativas? Essas regras estabelecem como um número limitado de elementos acústicos deve ser combinado na produção de diferentes seqüências vocais. No caso dos idiomas humanos, essas regras estabelecem as seqüências de palavras ou sentenças gramaticalmente corretas. No caso dos animais, determinariam seqüências partilhadas pelos membros da espécie. Apesar do interesse, poucos exemplos de processos generativos têm sido descritos em animais. Hailman, Ficken e Ficken (1985), por exemplo, mostraram como Parus atricapillus, uma ave norte-americana, recombina os quatro elementos acústicos presentes em seu canto, em centenas de diferentes seqüências vocais, através de poucas regras do tipo "o elemento a é sempre seguido de outro a ou de d". Quais são as regras de recombinação presentes nos intercâmbios dos muriquis é uma pergunta à qual estamos atualmente dedicando atenção e que talvez traga respostas surpreendentes.
Que animais na floresta produzam em seqüência seus chamados, com pouca repetição individual dentro de cada episódio e de modo diferenciado de acordo com a distância dos outros indica a existência de um conhecimento social e de uma avaliação da circunstância externa. Os animais não "conversam", como pode sugerir a imagem usada por Karen Strier; se conversassem, haveria a alternância emissor/receptor que se dá quando duas pessoas batem papo. Mas estão fornecendo coordenadas importantes para a progressão do grupo e para a permanência de cada indivíduo a uma distância razoável. É possível que cada chamado de muriqui carregue sua marca individual, sua assinatura. Pesquisadores com grande tempo de permanência no campo, em Caratinga, relatam reconhecer determinados muriquis pelo seu relincho, ao longe. Os macacos também podem ser capazes deste reconhecimento.
Os sub-sistemas vocais "para perto" e "para longe", o uso, por indivíduos, de chamados que favoreçam a localização de uma fonte alimentar pelo restante ou por alguns outros refletem a flexibilidade do comportamento espacial dos muriquis. Todos os membros do grupo podem coletar alimento ou descansar numa única árvore, também podem dispersar-se em uma área extensa da floresta, ou formar subgrupos que utilizem diferentes regiões de montes e vales, um indivíduo pode permanecer periférico, mesmo quando o restante do grupo está coeso etc. Como o sistema de vocalização representa esta dinâmica espacial e como funciona para que haja coordenação do grupo é uma pergunta etológica, mas também uma maneira de abordar o estudo da cognição animal, inserindo-a no contexto ecológico.
Referências Bibliográficas
Ades, C. (1993) Por uma história natural da memória. Psicologia USP, 4, 25-28. [ Links ]
Altmann, J. (1974) Observational study of behavior: sampling methods. Behaviour, 49,227-262. [ Links ]
Boinski, S. (1993) Vocal coordination of group movement among white-faced capuchin monkeys, Cebus capucinus. American Journal of Primatology, 30, 85-100. [ Links ]
Chapman, C. A. e Lefebre, L. (1990) Manipulating foraging group size: spider monkey food calls at fruiting trees. Animal Behaviour, 39, 739-751. [ Links ]
Cheney, D. L. e Seyfarth, R. M. (1990) In the mind of monkeys. Natural History, September, 38-47. [ Links ]
Darwin, C. (1872) The expression of the emotions in man and annuals. London: Murray. [ Links ]
Dittus, W. (1984) Toque macaque food calls: semantic communication concerning food distribution in the environment. Animal Behaviour, 32, 470-477. [ Links ]
Gardner, B.T.; Gardner, R. A. e Van Cantfort, T. E. (Org.) (1989) Teaching sign language to chimpanzees. Albany: State University of New York Press. [ Links ]
Gardner, R. A.; e Gardner, B. T. (1994) Ethological roots of language. Em: R. A. Gardner, B.T. Chiarelli & Plooij, F.X. The ethological roots of culture. Kluwer Academic Publishers. [ Links ]
Hailman, J. P.; Ficken, M. S. e Ficken, R. W. (1985) The "chick-a-dee" call of Parus atricapillus: a recombinant system of animal communication comparated with written English. Semiotica, 56, 191-224. [ Links ]
Mastripieri, D. (1996) Gestural communication and its cognitive implications in pigtail macaques (Macaca nemestrina). Behaviour. 131 997-1022. [ Links ]
Mendes, F. D. C. (1990) Afiliação e hierarquia no muriqui: o grupo Matão de Caratinga. Tese de Mestrado. Universidade de São Paulo, SP, Brasil. [ Links ]
Mendes, F. D. C. (1995) Interações vocais no muriqui (Brachyteles arachnoides). Tese de Doutorado, Universidade de São Paulo, SP, Brasil. [ Links ]
Mendes, F. D. C. (no prelo). Padrões de interação vocal do muriqui (Brachyteles arachnoides hypoxanthus). A Primatologia no Brasil. [ Links ]
Mendes, F. D. C. e Ades, C. (1995) O papel da comunicação vocal na coordenação do espaçamento intragrupal de primatas. Anais de Etologia, 13, 148-159. [ Links ]
Nowicki, S. eNelson, D. A. (1990) Defining natural categories in acoustic signals: comparison of three methods applied touchick-a-dee" call notes. Ethology, 86, 89-101. [ Links ]
Palombit, R. A. (1992) A preliminary study of vocal communication in wild long-tailed macaques (Macaca fascicularis). II. Potential of calls to regulate intragroup spacing. International Journal of Primatology, 13, 183-207. [ Links ]
Povinelli, D. J. (1993) Reconstructing the evolution of mind. American Psychologist, 48, 493-509. [ Links ]
Premack, D. (1983) The codes of man and beast. Behavioral and Brain Sciences, 6, 125-167. [ Links ]
Quine, W. V. (1960) Word and object. Cambridge, Massachusetts: MIT Press. [ Links ]
Rimoli, J. (1994) Estratégias de forrageamento de um grupo de muriquis (Brachyteles arachnoides, Primates, Cebidae) da Estação Biológica de Caratinga, MG. Dissertação de Mestrado, Instituto de Psicologia, USP, SP. [ Links ]
Robinson, J. G. (1982) Vocal systems regulating within-group spacing. Em: C.T. Snowdon, C.H. Brown & M. Peterson (Org.). Primate communication. Cambridge: Cambridge University Press. [ Links ]
Savage-rumbaugh, E. S. (1990) Language acquisition in a nonhuman species: implications for the innateness debate. Developmental Psychobiology, 23, 599-620. [ Links ]
Seyfarth, R. M; Cheney, D. L. e Marler, P. (1980) Vervet monkey alarm calls: semantic communication in a free-ranging primate. Animal Behaviour, 28, 1070-1094. [ Links ]
Smith, H. J.; Newman, J. D. e Symmes, D. (1982) Vocal concomitants of affiliative behavior in squirrel monkeys. Em: C. T. Snowdon, C. H. Brown & M. Peterson (Org.). Primate communication. Cambridge: Cambridge University Press. [ Links ]
Snowdon, C. T. (1982) Linguistic and psycholinguistic approaches to primate communication. Em: C. T. Snowdon, C. H. Brown & M. Peterson (Org.). Primate communication. Cambridge: Cambridge University Press. [ Links ]
Strier, K. B. (1986) The behavior and ecology of the wooly spider monkey or muriqui (Brachyteles arachnoides - E. Geoffroy 1806). Tese de Doutoramento, Universidade de Harvard, Cambridge, USA. [ Links ]
Strier, K. B. (1992) Faces in the forest; the endangered Muriqui monkeys of Brazil. New York: Oxford University Press. [ Links ]
(1) A pesquisa descrita, que é parte da tese de doutorado de F.D.C. Mendes, benefíciou-se com auxílios FAPESP, CNPq e FINEP. Queremos expressar nossos agradecimentos a Charles T. Snowdon e a Karen B. Strier, ambos da Universidade de Wisconsin em Madison, com os quais a maioria das idéias deste artigo foram discutidas. As análises descritas foram realizadas no laboratório do Dr. Snowdon
