










Serviços Personalizados
Journal

artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTIIndicadores
Compartilhar

 Permalink
PermalinkPsicologia: teoria e prática
versão impressa ISSN 1516-3687
Psicol. teor. prat. vol.13 no.3 São Paulo dez. 2011
ARTIGO ORIGINAL
Sensibilidade ao contraste espacial de adultos jovens para grades senoidais verticais em condições diferentes de luminância
Spatial contrast sensitivity of young adults for vertical sine-wave gratings in different conditions of luminance
La sensibilidad al contraste espacial de los adultos jóvenes a la verticales redes sinusoidales en condiciones diferentes de luminancia
Jandilson Avelino da Silva; Maria José Nunes Gadelha; Michael Jackson Oliveira de Andrade; Joenilton Saturnino Cazé da Silva; Natanael Antonio dos Santos
Universidade Federal da Paraíba, João Pessoa - PB - Brasil
RESUMO
O objetivo deste estudo foi medir a função de sensibilidade ao contraste (FSC) de adultos jovens para frequências espaciais de grades senoidais verticais em condições de luminância fotópica e escotópica. Utilizou-se o método psicofísico da escolha forçada para estimativa da sensibilidade ao contraste para as frequências de 0,6, 2,5 e 5,0 cpg. Todos os participantes apresentavam acuidade visual normal ou corrigida e estavam livres de doenças oculares identificáveis. As análises estatísticas mostraram que houve diferenças entre as condições de luminância e que os adultos foram mais sensíveis às frequências média e alta na condição de luminância fotópica. Esses resultados indicam que as características do sistema visual humano para detectar frequências espaciais em condições de luminância fotópica e escotópica passam por mecanismos distintos que tornam possível a diferenciação de contraste.
Palavras-chave: sensibilidade ao contraste; grade senoidal vertical; método psicofísico; luminância fotópica; luminância escotópica.
ABSTRACT
The aim of this study was to measure the contrast sensitivity function (CSF) of young adults for spatial frequencies of vertical sine-wave gratings in terms of photopic and scotopic luminance. We used the forced-choice psychophysical method to estimate contrast sensitivity for frequencies of 0.6, 2.5 and 5.0 cpd. All participants had normal or corrected acuity and they were free of ocular pathologies. Statistical analysis showed that there were differences between the conditions of luminance, where adults were more sensitive to middle and high frequencies in the photopic luminance condition. These results indicate that the characteristics of human visual system to detect spatial frequencies in photopic conditions of luminance and scotopic pass through distinct mechanisms that make possible the differentiation of contrast.
Keywords: contrast sensitivity; vertical sine-wave gratings; psychophysical method; photopic luminance, scotopic luminance.
RESUMEN
El objetivo de este estudio fue medir la función de sensibilidad al contraste (FSC) de los adultos jóvenes para frecuencias espaciales de las verticales redes sinusoidales en condiciones de luminosidad fotópica y escotópica. Se utilizó el método psicofísico de elección forzada para estimar la sensibilidad al contraste para las frecuencias de 0,6, 2,5 y 5,0 cpg. Todos los participantes presentaban acuidad visual normal o corigida y estaban libres de patología ocular. El análisis estadístico mostró que hay diferencias entre las condiciones de luminancia, y que los adultos son más sensibles a las frecuencias medias y altas en la condición de luminancia fotópica. Estos resultados indican que las características del sistema visual humano para la detección de frecuencias espaciales en condiciones de luminancia fotópica y escotópica pasan por diferentes mecanismos que hacen posible la diferenciación de contraste.
Palabras clave: sensibilidad al contraste; verticales redes sinusoidales; método psicofísico; luminanciafotópica, luminancia escotópica.
Introdução
A sensibilidade ao contraste espacial corresponde ao mínimo de contraste necessário à habilidade de discriminar um objeto de seu meio circunvizinho. A literatura discute que grupos de vias visuais paralelas constituídas por células parvocelulares e magnocelulares estão envolvidos nas diferenças encontradas na sensibilidade ao contraste acromático sob condições de luminância fotópica e escotópica, respectivamente (MIKI et al., 2008).
A organização funcional do sistema visual através de vias ou canais múltiplos e paralelos parece ser determinante na análise e no processamento de padrões de luminância espaciais pela retina e pelo córtex visual primário (CASAGRANDE; NORTON, 1991; GOODALE; MILNER, 1992; VIDYASAGAR et al., 2002). As evidências foram descritas em vias diferentes que passam pelo núcleo geniculado lateral (NGL), pelo córtex visual primário e por áreas visuais extraestriadas (MERIGAN; MAUNSELL, 1993). Entre essas vias que surgem na retina e se projetam para as áreas visuais corticais estriadas e extraestriadas através do NGL, destacam-se as vias parvocelular e magnocelular (BULLIER; HENRY, 1980; CASAGRANDE; NORTON, 1991; CASAGRANDE; KAAS, 1994; DE YOE; VAN-ESSEN, 1985; LIVINGSTONE; HUBEL, 1987; MAUNSELL; NEWSOME, 1987; SHIPP; ZEKI, 1985). Estudos mostram uma convergência dessas vias em áreas extraestriadas (DE YOE; VAN-ESSEN, 1985; KIPER; FENSTEMAKER; GEGENFURTNER, 1997; MAUNSELL; NEALY; DEPRIEST, 1990; MERIGAN; MAUNSELL, 1993; SHIPP; ZEKI, 1985).
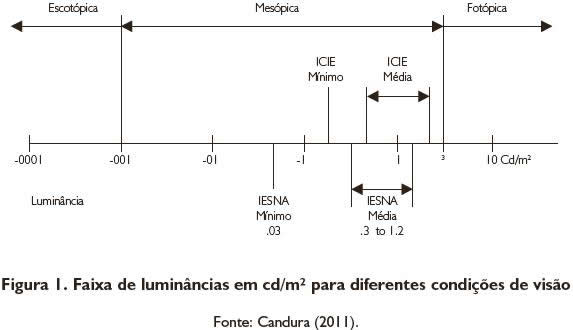
Uma maneira de medir o comportamento dessas vias no processamento de padrões formados por distribuições espaciais de luminância é utilizar a sensibilidade ao contraste, um indicador dos atributos fisiológicos e psicofísicos (CAMPBELL; MAFFEI, 1974; DE VALOIS; DE VALOIS, 1988). A luminância é uma grandeza fotométrica relacionada fundamentalmente ao brilho ou à quantidade de luz refletida de uma superfície (HERNÁNDEZ-LLOREDA, 2001). Uma faixa de luminâncias em cd/m² para diferentes condições de visão pode ser observada na Figura 1.
As principais unidades fisiológicas de fototransdução do sistema visual são os fotorreceptores, classificados em cones e bastonetes. Eles são ativados em níveis diferentes de luminância como a visão fotópica ou diurna mediada pelos cones e a visão escotópica ou noturna mediada pelos bastonetes (BRADLEY; SWITKES; DE VALOIS, 1985). Nesse contexto, a via parvocelular formada predominantemente pelos cones opera melhor na percepção de padrões cromáticos e acromáticos com luminâncias médias e altas cruciais para a percepção de objetos. Já a via magnocelular formada principalmente pelos bastonetes não responde a padrões cromáticos e opera melhor na percepção de movimento e padrões com luminâncias baixas que dão as características mais globais dos objetos (MERIGAN; KATZ; MAUNSELL, 1991; MULLEN; KINGDOM, 1991; PURVES et al., 2001).
A função de sensibilidade ao contraste (FSC) é uma medida clássica utilizada para obter respostas do sistema visual humano a uma ampla faixa de frequências espaciais. Ela é o inverso da curva de limiar de contraste, ou seja, quanto menos contraste o indivíduo precisa para perceber um estímulo, maior é sua sensibilidade (GADELHA et al., 2010; SANTOS et al., 2009).
A FSC possibilita que os aspectos fundamentais da resolução espacial sejam analisados através das vias visuais a partir da caracterização da sensibilidade ao contraste em diversas condições de luminância (CAMPBELL; ROBSON, 1968; CAMPBELL; MAFFEI, 1974; CORNSWEET, 1970; HERNÁNDEZ-LLOREDA, 2001). Dessa forma, a redução ou o aumento da intensidade de contraste necessário para detectar um estímulo pode ser um indicador de alterações nos mecanismos que processam o contraste (CAMPBELL; ROBSON, 1968; SANTOS; SIMAS, 2002).
As frequências espaciais são usadas como padrões periódicos externos no espaço. Elas podem ser unidimensionais (faixas claras ou escuras, por exemplo) e são especificadas pela unidade de medida ciclos por grau (cpg) de ângulo visual (COSTA et al., 2009; SANTOS; SIMAS, 2001).
De Valois e De Valois (1988) elaboraram uma representação das frequências espaciais baixas, médias e altas por meio da FSC. A via parvocelular está envolvida na resolução espacial de frequências espaciais médias e altas, e a via magnocelular responde principalmente às frequências espaciais baixas (BERNINGER et al., 1989; KULIKOWSKI, 1991; LEE et al., 2000; KULIKOWSKI; MURRAY; PARRIS, 1989; MERIGAN; KATZ; MAUNSEL, 1991; MIKI et al., 2008; SUTTLE; HARDING, 1999; SOUZA et al., 2007).
Estudos já demonstram a existência de mecanismo de resposta funcional relacionado à sensibilidade ao contraste em recém-nascidos e em crianças, nas condições de luminância fotópica e escotópica (PEIPER, 1926; TRINCKER; TRINCKER, 1955). Além disso, foi mostrado que a sensibilidade ao contraste nas células M é geralmente muito mais alta do que nas células P (DERRINGTON; LENNIE, 1984; KAPLAN; SHAPLEY, 1982).
Outros estudos determinaram o limiar de contraste acromático a partir de grades senoidais registrando atividades nas células magno e parvocelular (DERRINGTON; LENNIE, 1984). Resultados relacionados à idade mostraram perda na informação espacial em condição escotópica para frequências espaciais baixas e um declínio para as frequências altas (JACKSON; OWSLEY, 2000; SCHEFRIN et al., 1998, 1999).
Ainda que esses achados da literatura apresentem resultados acerca da sensibilidade ao contraste relativos às principais vias visuais, em sua grande maioria, eles são apresentados em apenas um único nível de luminância, geralmente fotópica. Desse modo, acreditando que luminâncias diferentes possam interferir na sensibilidade ao contraste, o objetivo deste estudo foi medir a FSC de adultos jovens para frequências espaciais de grades senoidais verticais em condições de luminância fotópica e escotópica.
Métodos
Participantes
Participaram deste estudo nove voluntários (seis mulheres e três homens), na faixa etária de 19 a 24 anos (M = 21,11; DP = 2,02). Todos os participantes apresentavam acuidade visual normal ou corrigida, avaliados pela cartela de optótipos "E" de Rasquim, e estavam livres de qualquer doença ocular ou neuropsicológica identificável. A participação aconteceu de forma voluntária por meio de um Termo de Consentimento Livre e Esclarecido devidamente assinado, de acordo com a Resolução no 196/96 do Conselho Nacional de Saúde (Ministério da Saúde), que trata das diretrizes e normas de pesquisa envolvendo seres humanos, submetido e aprovado pelo Comitê de Ética e Pesquisa do Hospital Universitário Lauro Wanderley sob Protocolo no 76/10.
Equipamentos e estímulos
As medidas de sensibilidade ao contraste foram realizadas em um monitor de vídeo colorido LG/RCT (Cathodic Ray Tube), com tela plana de 19 polegadas. A tela do monitor apresentava resolução de 1.024 x 768 pixels e taxa de atualização de 70 Hz. O monitor era controlado por um microcomputador com uma placa de vídeo de entrada VGA e DVI, conectado ao hardware Bits++ (Cambridge Research Systems, Rochester, Kent, England). A função do Bits++ era aumentar a resolução da tela do monitor de 8 para 14 bits, possibilitando a geração de estímulos com melhor definição.
A luminância e a correção gama do monitor foram controladas com um programa LightScan e um fotômetro OptiCAL (Cambridge Research Systems, Rochester, Kent, England). O valor da luminância média do monitor na condição fotópica foi de 42,6, e na condição escotópica, de 0,09 cd/m2. Utilizou-se um programa em linguagem C++ para gerar os estímulos e rodar os experimentos, desenvolvido pelo Laboratório de Percepção, Neurociências e Comportamento (LPNeC-UFPB) em conjunto com o Laboratório de Percepção Visual (LabVis-UFPE).
O ambiente experimental tinha dimensões de 2,5 por 2,0 metros e era iluminado por uma lâmpada fluorescente Philips de 20 W na condição fotópica e escuro na condição escotópica. Uma cadeira foi fixada à distância de 150 cm do monitor. A resposta dos participantes foi registrada por meio de um mouse.
Utilizaram-se estímulos visuais acromáticos do tipo grade senoidal vertical com frequências espaciais de 0,6, 2,5 e 5,0 cpg, e um estímulo neutro cinza homogêneo com luminância média. Todos os estímulos eram circulares com diâmetro de aproximadamente 7,2 graus de ângulo visual, gerados no centro da tela do monitor. Exemplos dos estímulos podem ser encontrados na Figura 2.

Procedimento
Utilizou-se um delineamento experimental com medidas repetidas, no qual o "N" estatístico foi obtido a partir do número de reversões ou valores de máximos e mínimos de contraste, como é peculiar às pesquisas em percepção visual com métodos psicofísicos. O delineamento foi composto por duas variáveis independentes (condição de luminância e frequência espacial) e uma variável dependente, o valor de limiar ao contraste. Todos os participantes passaram por todas as condições em momentos e ordem diferentes. O efeito de ordem foi controlado pela técnica de contrabalanceamento.
Mediu-se o limiar de contraste com o método psicofísico da escolha forçada entre duas alternativas temporais (SANTOS et al., 2006), no qual o participante teve que escolher, entre dois estímulos, qual continha a frequência de teste. Esse método se baseia na probabilidade de acertos consecutivos por parte do voluntário, ou seja, em cerca de 100-150 apresentações de escolhas entre os dois estímulos, o estímulo de teste é percebido, por exemplo, em 79% das vezes.
Nas duas condições, o procedimento para medir o limiar para cada frequência espacial consistiu na apresentação sucessiva simples de pares de estímulos, sendo um deles uma frequência de teste e o outro um estímulo neutro de padrão homogêneo com luminância média. O critério adotado para variar o contraste do estímulo foi o de três acertos consecutivos para diminuir o contraste em 20% e um erro para aumentar o contraste na mesma unidade. Cada estímulo foi apresentado por 2 s, com intervalo de 1 s entre eles e de 3 s entre tentativas. Todas as medidas foram realizadas com visão binocular e pupila natural.
Antes do início de cada sessão experimental, os voluntários foram orientados a pressionar o botão esquerdo do mouse quando o estímulo de teste fosse apresentado primeiro, e o botão direito do mouse quando o estímulo de teste fosse apresentado em segundo lugar. Os voluntários receberam a seguinte instrução: "Serão apresentados pares de estímulos, um com listras claro-escuras e outro totalmente cinza. Você deverá escolher sempre o estímulo que contém as listras, pressionando o botão do lado esquerdo (botão número 1) do mouse quando o estímulo com listras for apresentado primeiro, e o botão do lado direito (botão número 2) quando for apresentado em segundo lugar (após o estímulo cinza)". Cada sessão foi iniciada com o estímulo teste com contraste em nível supralimiar, e os experimentos só começaram quando o experimentador certificou-se de que os participantes entenderam e responderam conforme as instruções.
As sessões experimentais tinham duração de 5 a 10 minutos, e cada frequência foi medida duas vezes, aleatoriamente, em dias diferentes. Nesse tipo de procedimento, o número de apresentações necessárias para determinar o limiar de contraste varia de acordo com os acertos e erros dos voluntários, bem como do número de máximos e mínimos ou reversões que se quer obter. A apresentação dos estímulos para os participantes em condição escotópica ocorreu depois de uma adaptação de 30 minutos na ausência de luz. Assim, a sessão experimental terminou automaticamente assim que três valores máximos e três mínimos de contrastes (ou seis reversões) foram obtidos.
Resultados
Os valores de máximos e mínimos do limiar de contraste obtidos a partir das seis reversões foram dispostos em planilhas, de acordo com a condição de luminância, e em seguida transformados em sensibilidade ao contraste para a realização do tratamento estatístico. A Figura 3 apresenta os valores de sensibilidade ao contraste para as frequências de 0,6, 2,5 e 5 cpg nas condições de luminância fotópica e escotópica. O erro padrão das médias está representado pelas barras verticais (Figura 3) e possui um intervalo de confiança de 95%, corrigido para o tamanho da amostra pelo teste estatístico t-Student.

Na Figura 3, pode-se observar que a máxima sensibilidade ocorreu em pontos diferentes para cada condição de luminância. Na condição escotópica, o pico da curva se localiza na frequência baixa (0,6 cpg). Já na condição fotópica, o ápice da sensibilidade ao contraste ocorreu para a frequência média (2,5 cpg). Ambas as curvas tiveram reduções na sensibilidade para a frequência alta (5,0 cpg).
A Anova para medidas repetidas mostrou que houve um efeito principal entre as frequências espaciais e as condições de luminância com F (4, 211) = 73,40, (p < 0,05). Já o teste post hoc Tukey HSD mostrou diferenças entre as frequências de 2,5 e 5,0 (p < 0,05), onde os voluntários demonstraram uma sensibilidade cerca de três vezes maior na condição de luminância fotópica.
Os resultados mostram que o desempenho da FSC depende da condição de luminância, sugerindo que o processamento visual é realizado por vias que processam informações distintas, a via dos cones e a dos bastonetes.
Discussão
O modelo de canais múltiplos assume que cada faixa estreita de frequência espacial é conduzida por um canal independente. A função de sensibilidade ao contraste é o envelope de canais para as diversas propriedades visuais, sendo cada um sensível a faixas diferentes de frequências espaciais (BRADDICK; CAMPBELL; ATKINSON, 1978; GRAHAM; NACHMIAS, 1971; SANTOS; SIMAS, 2001).
As propriedades espaciais da sensibilidade ao contraste apresentam mecanismos sensório-visuais diferenciados. Os neurônios corticais são altamente seletivos para frequências espaciais, tornando possível que as diferenças de respostas associadas com frequências espaciais baixas, médias e altas reflitam o papel funcional das vias visuais magnocelular e parvocelular (JONHSON; HAWKEN; SHAPLEY, 2001; SKOTTUN; SKOYLES, 2007).
Como já era esperado, de acordo com estudos anteriores (KULIKOWSKI; MURRAY; PARRIS, 1989; BERNINGER et al., 1989; KULIKOWSKI, 1991; SUTTLE; HARDING, 1999; MERIGAN; KATZ; MAUNSEL, 1991; MIKI et al., 2008; SKOTTUN; SKOYLES, 2007), neste trabalho os neurônios do sistema magnocelular tenderam a responder preferencialmente a estímulos de baixa frequência espacial (0,6 cpg). Já o sistema parvocelular teve maior sensibilidade nas frequências espaciais média e alta (2,4 e 5 cpg).
Foi possível observar, no referido estudo, que a ativação dos bastonetes em condições de luminância escotópica representa uma curva de sensibilidade ao contraste com efeito decrescente, apresentando maior valor de sensibilidade ao contraste da frequência espacial mais baixa para a mais alta (0,6 = 77,2; 2,5 = 61,4; 5 = 39,4). Observou-se ainda que o contrário ocorreu com as medidas de sensibilidade ao contraste em condições de luminância fotópica, nas quais a ativação dos cones aconteceu com um acréscimo na sensibilidade de acordo com o aumento das frequências espaciais (0,6 = 75,3; 2,5 = 181,6; 5 = 121,4). Esses achados são semelhantes aos encontrados por Ellemberg et al. (2001) e corroboram o estudo de Bradley, Switkes e De Valois (1985), na medida em que ambos sugerem que as sensibilidades ao contraste possuem respostas distintas para os níveis diferentes de luminância.
Respostas puramente magno ou parvocelular ainda não são tão claras e não se sabe ao certo como essas vias interagem na prática fora do laboratório (SKOTTUN; SKOYLES, 2007). A inexistência de diferenças significantes para as frequências baixas, verificada neste estudo, pode ser relacionada às propriedades das vias parvo e magnocelular, uma vez que, dependendo das condições de luminância, tanto a via parvocelular quanto a magnocelular podem estar sendo ativadas em conjunto ou com certa predominância de uma delas. Isso pode deslocar a sensibilidade para as frequências mais baixas ou mais altas, podendo chegar a um ponto em que a sensibilidade em condições fotópica e escotópica pode coincidir em algum faixa de frequência.
Em conclusão, esses resultados demonstram que as características do sistema visual humano para detectar frequências espaciais em condições de luminância fotópica e escotópica passam por mecanismos distintos que tornam possível a diferenciação de contraste. As vias visuais respondem de forma específica a cada informação de luminância, permitindo entender como a FSC de adultos jovens se comporta ao processar padrões de imagens espaciais.
Considerações finais
Com base nos resultados deste estudo, podemos inferir que existem grupos de neurônios sintonizados (ou canais) para faixas estreitas de frequências espaciais diferentes em condições de luminância fotópica e escotópica. A função de sensibilidade ao contraste pode ser uma ferramenta importante para descrever e avaliar o funcionamento desses canais em voluntários saudáveis e com comprometimento dos sistemas visual e nervoso decorrente de patologias. Ainda assim, é necessário realizar novas pesquisas, usando outros tipos de estímulos e condições para tentar descrever de forma mais sistemática o papel das vias magnocelular e parvocelular no processamento de padrões espaciais em condições de luminância diferente.
Referências
BERNINGER, T. A. et al. Separable evoked retinal and cortical potentials from each major visual pathway: preliminary results. British Journal of Ophthalmology, v. 73, p. 502-511, 1989. [ Links ]
BRADDICK, O.; CAMPBELL, F. W.; ATKINSON, J. Channels in vision: basic aspects. In: HELD, R.; LEIBOWITZ, H. W.; TEUBER, H. L. (Ed.). Handbook of sensory physiology. New York: Springer-Verlag, 1978. v. 5, p. 3-38. [ Links ]
BRADLEY, A.; SWITKES, E.; DE VALOIS, K. K. Orientation and spatial frequency selectivity of adaptation to isoluminant color patterns. Ophthalmologic Visual Science, v. 26, p. 182, 1985. [ Links ]
BULLIER, J.; HERING, G. H. Ordinal position and afferent input of neurons in monkey striate cortex. Journal Comparative Neurologic, v. 42, p. 1271-1281, 1980. [ Links ]
CAMPBELL, F. W.; MAFFEI, L. Contrast and spatial frequency. Scientific American, v. 231, p. 106-114, 1974. [ Links ]
CAMPBELL, F. W.; ROBSON, F. G. Application of the Fourier analysis to the visibility of gratings. Journal of Physiology, v. 197, p. 551-566, 1968. [ Links ]
CANDURA, P. Visão humana. Disponível em: <http://www.lume.com.br/pdf/ed03/ed_03_Ilum.pdf>. Acesso em: 21 jul. 2011. [ Links ]
CASAGRANDE, V. A.; KAAS, J. H. Cerebral cortex. New York: Plenum Press, 1994. p. 201-259. [ Links ]
CASAGRANDE, V. A.; NORTON, T. T. The neural basis of vision function: vision and visual dysfunction. London: Macmillan Press, 1991. p. 41-84. [ Links ]
CORNSWEET, T. N. Vision perception. New York: Academic Press, 1970. [ Links ]
COSTA, T. L. et al. Envelhecimento humano e sensibilidade ao contraste fotópica para frequências angulares. Psicologia: teoria e pesquisa, v. 25, p. 589-593, 2009. [ Links ]
DE VALOIS, R. L.; DE VALOIS, K. K. Spatial vision. New York: Oxford University Press, 1988. [ Links ]
DE YOE, E. A.; VAN-ESSEN, D. C. Concurrent processing streams in monkey visual cortex. Trends in Neuroscience, v. 11, p. 219-226, 1985. [ Links ]
DERRINGTON, A. M.; LENNIE, P. Spatial and temporal contrast sensitivities of neurons in lateral geniculate nucleus of Macaque. Journal of Physiology, v. 357, p. 219-240, 1984. [ Links ]
ELLEMBERG, D. et al. Contrast dependency of VEPs as a function of spatial frequency: the parvocellular and magnocellular contributions to human VEPs. Spatial Vision, v. 15, p. 99-111, 2001. [ Links ]
GADELHA, M. J. N. et al. Sensibilidade ao contraste acromático para grades senoidais verticais em adolescentes e adultos. Psicologia: teoria e prática, v. 12, n. 1, p. 59-70, 2010. [ Links ]
GRAHAM, N.; NACHMIAS, J. Detection of grating patterns containing two spatial frequencies: a comparison of single-channel and multiple channel models. Vision Research, v. 11, p. 251-259, 1971. [ Links ]
GOODALE, M. A.; MILNER, A. D. Separate visual pathways for perception and action. Trends in Neurosciences, v. 15, p. 20-25, 1992. [ Links ]
HERNÁNDEZ-LLOREDA, M. J. Mecanismos de detección del contraste cromático y de luminancia en el sistema visual humano. Anales de Psicologia, v. 17, p. 219-233, 2001. [ Links ]
JACKSON, G. R.; OWSLEY, C. Scotopic sensitivity during adulthood. Vision Research, v. 40, p. 2467-2473, 2000. [ Links ]
JONHSON, E. N.; HAWKEN, M. J.; SHAPLEY, R. The spatial transformation of color in the primary visual cortex of the macaque monkey. Nature Neuroscience, v. 4, n. 1, p. 409-416, 2001. [ Links ]
KAPLAN, E.; SHAPLEY, R. M. X and Y cells in the lateral geniculate nucleus of macaque monkeys. Journal of Physiology, v. 330, p. 125-143, 1982. [ Links ]
KIPER, D. C.; FENSTEMAKER, S. B.; GEGENFURTNER, K. R. Chromatic properties of neurons in macaque area V2. Visual Neuroscience, v. 14, p. 1061-1072, 1997. [ Links ]
KULIKOWSKI, J. J. On the nature of visual evoked potentials, unit responses and psychophysics. In: VALBERG, A.; LEE, B. B. From pigments to perception. New York: Plenum Press, 1991. p. 197-208. [ Links ]
KULIKOWSKI, J. J.; MURRAY, I. J.; PARRIS, N. R. A. Electrophysiological correlates of chromatic-opponent and achromatic stimulation in man. In: DRUM, B.; VERRIEST, E. Colour vision deficiencies IX. Dordrechr: Academic, 1989. p. 145-153. [ Links ]
LEE, B. B. et al. Visual responses of ganglion cells of a New-World primate, them capuchin monkey. Cebus Paella, v. 528, n. 3, p. 573-590, 2000. [ Links ]
LIVINGSTONE, M. S.; HUBEL, D. H. Psychophysical evidence for separate channels for the perception of form, color, movement, and depth. Journal of Neuroscience, v. 7, p. 3416-3468, 1987. [ Links ]
MAUNSELL, J. H.; NEALY, T. A.; DEPRIEST, D. D. Magnocellular and parvocellular contributions to responses in the middle temporal visual area (MT) of the macaque monkey. Journal of Neuroscience, v. 10, p. 3323-3334, 1990. [ Links ]
MAUNSELL, H. R.; NEWSOME, W.T. Visual processing in the monkey extra striate cortex. Annual Review of Neuroscience, v. 10, p. 363-401, 1987. [ Links ]
MERIGAN, W. H.; KATZ, L. M.; MAUNSELL, J. H. The effects of parvocellular lateral geniculate lesions on the acuity and contrast sensitivity of macaque monkeys. Journal of Neuroscience, v. 11, n. 4, p. 994-1001, 1991. [ Links ]
MERIGAN, W. H.; MAUNSELL, J. H. R. How parallel are the primate visual pathways? Annual Review of Neuroscience, v. 16, p. 369-402, 1993. [ Links ]
MIKI, A. et al. Magno-and parvocellular visual cortex activation in anisometropia, as studied with functional magnetic resonance imaging. Neuroophthalmology, v. 32, p. 187-193, 2008. [ Links ]
MULLEN, K. T.; KINGDOM, F. A. A. Colour contrast in form perception. In: GOURAS, P. (Ed.). The perception of colour. Oxford: MacMilan, 1991. p. 198-217. [ Links ]
PEIPER, A. Ueber die Helligkeits und Farbenempfindungen der Fruhgeburten. Arch kinderheilk, v. 80, p. 1-20, 1926. [ Links ]
PURVES, D. et al. Neuroscience. 2. ed. Sunderland, MA: Sinauer, 2001. [ Links ]
SANTOS, N. A.; SIMAS M. L. B. Função de sensibilidade ao contraste: Indicador da percepção visual da forma e da resolução espacial. Psicologia: reflexão e crítica, v. 14, p. 589-597, 2001. [ Links ]
______. Função de sensibilidade ao contraste: indicador da percepção visual da forma e da resolução espacial. Psicologia: reflexão e crítica, v. 14, n. 3, p. 589-597, 2002. [ Links ]
SANTOS, N. A. et al. Mesopic radial frequency contrast sensitivity function for young and older adults. Brazilian Journal of Medical and Biological Research, v. 39, n. 6, p. 791-794, 2006. [ Links ]
______. Sensibilidade ao contraste mesópica de crianças surdas para estímulos radiais e grades senoidais. Psico, v. 40, n. 2, p. 227-234, 2009. [ Links ]
SCHEFRIN, B. E. et al. The area of complete scotopic spatial summation enlarges with age. Journal of the Optical Society of America, v. 15, p. 340-348, 1998. [ Links ]
SCHEFRIN, B. E. et al. Senescent changes in scotopic contrast sensitivity. Vision Research, v. 39, p. 3728-3736, 1999. [ Links ]
SHIPP, S.; ZEKI, S. Segregation of pathways leading from area V2 to areas V4 and V5 of macaque monkey visual cortex. Nature, v. 315, p. 322-325, 1985. [ Links ]
SKOTTUN, B. C.; SKOYLES, J. R. Contrast sensitivity and magnocellular functioning in schizophrenia. Vision Research, v. 47, p. 2923-2933, 2007. [ Links ]
SOUZA, G. S. et al. Spatial luminance contrast sensitivity measured with transient VEP: comparison with psychophysics and evidence of multiple mechanisms. Investigative Ophthalmology & Visual Science, v. 48, n. 7, p. 3396-3404, 2007. [ Links ]
SUTTLE, C. M.; HARDING, G. F. A. Morphology of transient VEPs to luminance and chromatic pattern onset and offset. Vision Research, v. 39, p. 1577-1584, 1999. [ Links ]
TRINCKER, D.; TRINCKER, I. Die ontogenetische Entwicklung des Helligkeits- und Farbensehens beim Menschen. I. Die Entwicklung des Helligkeitsshens. Albrecht Graefes Arch Ophthal, v. 56, p. 519-534, 1955. [ Links ]
VIDYASAGAR, T. R. et al. Convergence of parvocellular and magnocellular information channels in the primary visual cortex of the macaque. European Journal of Neuroscience, v. 16, p. 945-995, 2002. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Contato
Jandilson Avelino da Silva
e-mail: jandilsonsilva@gmail.com
Tramitação
Recebido em outubro de 2010
Aceito em junho de 2011
