










Serviços Personalizados
Journal

artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkPsicologia: teoria e prática
versão impressa ISSN 1516-3687
Psicol. teor. prat. vol.22 no.2 São Paulo maio/ago. 2020
https://doi.org/10.5935/1980-6906/psicologia.v22n2p83-104
ARTICLES
HUMAN DEVELOPMENT
Review and update of ultrasonic vocalization in animals: correlation with autism spectrum disorder experimental models?
Revisión y actualización de la vocalización ultrasónica en animales: ¿correlación con el modelo del trastorno del espectro autista?
Juciara da C. SilvaI ; Miriam O. RibeiroII; Déborah M. dos SantosI; Carlos Alberto A. PenattiI
; Miriam O. RibeiroII; Déborah M. dos SantosI; Carlos Alberto A. PenattiI
INove de Julho University (UNJ), São Paulo, SP, Brazil
IIMackenzie Presbyterian University (UPM), São Paulo, SP, Brazil
ABSTRACT
The ability to understand and to communicate is essential for the adapted social and cultural development of children. Autism spectrum disorders (ASD) are characterized by social interaction deficiencies in the verbal language domain and may incur in repetitive, stereotyped, and restricted gestural behaviors. Regarding animal models in ASD, many studies focus on gene expression, cortical neuronal migration and cell maturation, and neural network deficits. However, very few studies directly link animal oral communication and gene expression in cortical areas of language. The object of our study aimed to review and to correlate genes involved in ASD and those related to ultrasonic communication in animal model studies of language-based social behavior at the PubMed database. The PubMed search yielded 25 selected papers. Some findings showed critical genes, which modulate several aspects of verbal communication. Most of these genes can be markers of communication deficits.
Keywords: communication; ultrasound vocalization; ASD; genetics; neurodevelopment.
RESUMEN
La capacidad de comprender y comunicarse es esencial para el desarrollo social y cultural adaptado de los niños. El trastorno del espectro autista (TEA) se caracterizan por deficiencias de interacción social en el dominio del lenguaje verbal y pueden incurrir en comportamientos gestuales repetitivos, estereotipados y restringidos. Con respecto a los modelos animales en TEA, muchos estudios se centran en la expresión génica, la migración y maduración neuronal cortical y los déficits de la red neuronal, pero muy pocos son los estudios que relacionen directamente la comunicación oral animal y la expresión génica en áreas corticales del lenguaje. El objetivo de nuestro estudio tuvo como objetivo revisar y correlacionar genes involucrados en TEA y aquellos relacionados con la comunicación ultrasónica en estudios con modelos animales de comportamiento social basado en el lenguaje en la base de datos PubMed. La búsqueda en PubMed arrojó 25 artículos seleccionados. Algunos hallazgos mostraron genes críticos, que modulan varios aspectos de la comunicación verbal. Esta evidencia puede ofrecer la oportunidad como biomarcadores genéticos para los déficits de comunicación en los desórdenes en el neurodesarrollo.
Palabras clave: comunicación; vocalización por ultrasonido; TEA; genética; neurodesarrollo.
1. Introduction
The development and sophistication of human language are essential characteristics for our species. Although many animals have varied and elaborated communication systems, human communicability allows an array of almost infinite construction of oral sound phrases and meanings, not only to share needs and desires but also to establish an even more complex personal relationship (Werker & Hensch, 2015).
Vocal comprehension and expression comprise development in the broadest sense employing verbal and non-verbal assessments. The delays and interruptions in the acquisition of phases that are necessary for the maturation of communication may generate consequences that affect a child's cognitive, social, behavioral, and emotional functions and impact his/her progression into adulthood (Liu, Zahrt, & Simms, 2018).
Communication disorders in children include: delay only in expressive language or errors in the production of simple sounds (articulation); multiple errors in the production of sounds with some degree of deficit in motor planning or delay in expressive language; and mixed receptive language with difficulty in complex language processing (Liu, Zahrt, & Simms, 2018).
During healthy development, a series of communicative acts emerges and matures as the structural language develops. The maintenance of the conversational topic appears in interactions with adults, and the adequacy of responses increases. In the past decade, the communicative, social, and emotional aspects of language have followed a qualitative distribution in 27 observable behaviors. These are classified into five relevant domains for children aged between 5 and 11 years. The five domains are defined by 1. introduction and responsiveness (the ability to introduce communication and to be receptive to the communication of others); 2. non-verbal communication (usage and understanding of gestures, facial expressions, body postures and proximity between speakers); 3. socio-emotional attunement (interpreting the emotional reactions of others and demonstrating appropriate responses); 4. executive function (given the interactions and flexibility in the planning of the communicative content); and 5. negotiation (cooperating and negotiating properly with communicative partners). For children with communication deficits who develop verbal language, the previously described pragmatic difficulties persist and evolve into language deficits, which include unskilled attempts to initiate communication, narrower intervals of communication acts and difficulties in the production of communication of new language (Parsons, Cordier, Munro, Joosten, & Speyer, 2017).
Among neurodevelopmental disorders that can affect communication, there are autism spectrum disorders (ASD). This neuropsychological entity, which is characterized by deficiencies in expression/understanding of verbal language and social interactions with significant seclusion, unveils a series of variations demonstrated in repetitive, stereotyped behaviors and determines restricted social interests in many cases. Although there is no clinical correlation between ASD with specific mechanisms in neurobiological development per se, the proposition for early intervention with psychological follow-up and speech therapy supports improvement. In addition to specific pedagogical criteria, they have been shown to improve cognition and adaptive behavior by altering brain responses to social stimuli (Schuetze, Rohr, Dewey, McCrimmon, & Bray, 2017).
The diagnosis of ASD is based on communicative deficits that are observed in everyday social interactions. Deficits become evident in situations in which there is a divergence between verbal communication and the understanding of the literal meaning of a sentence or a phrase (in the case of irony or sarcasm, for example) and have been considered products of a primary commitment in representing mental states. Some studies suggest that communication deficits arise from the main difficulties with motivation, social attention, or cognitive flexibility (Wadge, Brewer, Bird, Toni, & Stolk, 2019).
The most recent edition of the 'Diagnostic and Statistical Manual of Mental Disorders' (DSM-5) removed language difficulties as a major condition of ASD. However, a significant proportion of children with ASD have difficulties in acquiring spoken language. The severity of these difficulties varies considerably. Most children with ASD acquire language during the preschool years, usually at the age of five. However, about 25 to 30 % do not develop any functional spoken language or remain minimally verbal. Language difficulties in children with ASD can result in several adverse sequelae, which include behavioral difficulties, deficits in adaptive functional skills, and a social relationship (Brignell et al., 2018).
Many children with ASD continue to demonstrate language skills below their peers. A great proportion of those youngsters do not obtain useful speech, which is defined as expressive language that can be used frequently, communicatively, referentially and semantically in a different way. Expressive language deficits are particularly important in light of the strong predictive relationship among the expressive language skills and social and adaptive outcomes for children with this type of disorder (McDaniel, D'Ambrose Slaboch, & Yoder, 2018).
Since the publication of the DSM-5, there has been a greater focus on social communication, particularly on how this skill should be assessed, and how it should be addressed. The assessment is challenging, because of the nature of how social communication must occur in a natural context where this ability can be observed directly and constantly in real-time (Hyter et al., 2017).
Animal experimentation research models counterbalance clinical research to tackle communication deficits in possible complementary approaches. Although they do not characterize a total exclusive or faithful state of human order, research with animal models is fundamental for the advances and better understanding in ASD clinical settings. They foster research settings that are experimentally controlled, rigorous in variables, treatable, and exploited mechanically for that type of disorder. Currently, some of the relevant experimental animal models show that rodents produce surprisingly complex ultrasonic vocalizations (USV) during their social and sexual interactions in an ecological/social adapted environment (Arriaga & Jarvis, 2013; Hanson & Hurley, 2012).
Among the species studied, mice have great potential in the study of ultrasound sounds (USVs), emulating the equivalent of a voice skill. Therefore, this form of vocal expression serves to provide a basic research system in animal communication, behavioral biology, and neuroscience and, additionally, collaborates to comprehend issues involved in biomedical sciences. Rodents produce both sound vocalizations in the human audible and ultrasonic ranges. They emit sounds in fundamental frequencies ranging from 100 up to 120,000 Hz. (Riede, Borgard, & Pasch, 2017). Mouse USVs appear to provide a variety of functions, including social recognition (individual, kinship, sex, and species recognition), rival bullying (intra-sexual selection), and partner attraction (inter-sexual selection) (Zala, Reitschmidt, Noll, Balazs, & Penn, 2017).
Intricate language patterns are unlikely to be instinctive for rodents. The complexity of ultrasonic vocalization patterns contains important communicative information. Quantitative assessment of the value of the media reward is not yet available for rodents or individuals with ASD (Mony, Hong, & Lee, 2018). Although there are some established models to assess or to work with ASD symptoms, newer and improved models are needed. Designing a behavioral model specifically relevant to human mental disorders presents a great challenge, as there is a discontinuity between human social interactions and those exhibited by non-primate mammals, such as rodents. Besides, ASD-related symptoms are unique, but inherently variable in humans.
Regarding the genetic approach, rodent research models that incorporate a growing list of changes in possible genes involved in language have been used to detail and deepen the understanding of the effects of genetic mutations on cellular/tissue formation, connectivity, physiology and neuronal behavior (Lazaro & Golshani, 2015).
The importance of the communicability of individuals considered as ASD prompts us to organize and highlight the main studies within basic research that use animal models focusing on ultrasonic vocalization. Specifically, we aim to assess the importance of these studies and future perspectives for the neurobehavioral canvas in developmental communication deficits.
2. Methodology
We searched for articles that used and analyzed the theme of ultrasonic vocalization in animal experimentation models correlated to basic ASD research.
2.1 Study inclusion and exclusion criteria
Experimental (original) and bibliographic review studies that presented results regarding the impact of ultrasonic vocalization on animal models and their possible interaction for a better understanding of ASD were included. The other inclusion criterion was: date of publication of the study from October 2017 until October 2019. The remaining articles were excluded from the period not described and/or outside the theme of ultrasonic vocalization as the objective of the experimental study.
2.2 Search strategy
The bibliographic search of the articles was carried out in the PubMed database, using the descriptors "autism ultrasonic vocalization," being conducted by two researchers independently. The abstracts read in full were selected, using the inclusion criteria adopted in the study.
The initial assessment, based on the titles and abstracts, was instrumental in rejecting those not in accordance with the inclusion criteria. After evaluating the feasibility, the article was read in its entirety.
3. Results and discussion
Communication deficits are characteristic of ASD. Depending on the individual's intellectual capacity, communication disarrangements can manifest as lack of speech, language delay, use of odd prosody and intonation, stereotyped speech, persevering phrases, and difficulties with language pragmatics, such as those involved in initiation and maintenance of appropriate language and meaningful conversations. Rodents emit complex ultrasonic vocalizations (USVs) during social meetings with their peers. Leading researchers assume that the USV emission features as a sign of potential complex communication. It is believed that USVs function as a means of social recognition and courtship behavior, which may indicate to provoke an approaching behavior and to communicate aggression. USVs vary between spectrotemporal parameters, such as frequency, duration, and intensity. These resources are used to analyze USVs in different categories by the researchers. USVs have become a model commonly used to emulate and, thus, to add to the understanding of human communication (Screven & Dent, 2019).
Currently, different types of USVs are recognized in rodents: emitted by isolated pups; produced by youngsters in interaction; vocalized by adult males exposed to females. The emission of USVs is used to study the characteristics related to ASD. For example, the puppies emit USVs causing a search behavior during the separation of the mother (Scattoni, Crawley, & Ricceri, 2009).
The data suggest that there are several models for a better understanding of ASD (e.g., Fmr1 for Fragile X syndrome, Mecp2 for Rett syndrome, and Tsc1/2 for tuberous sclerosis, synaptic molecule genes like neurexin, neuroligin, and Shank) and other risk genes. Most models of ASD were based on symptoms similar to human behavior (Takumi et al., 2019).
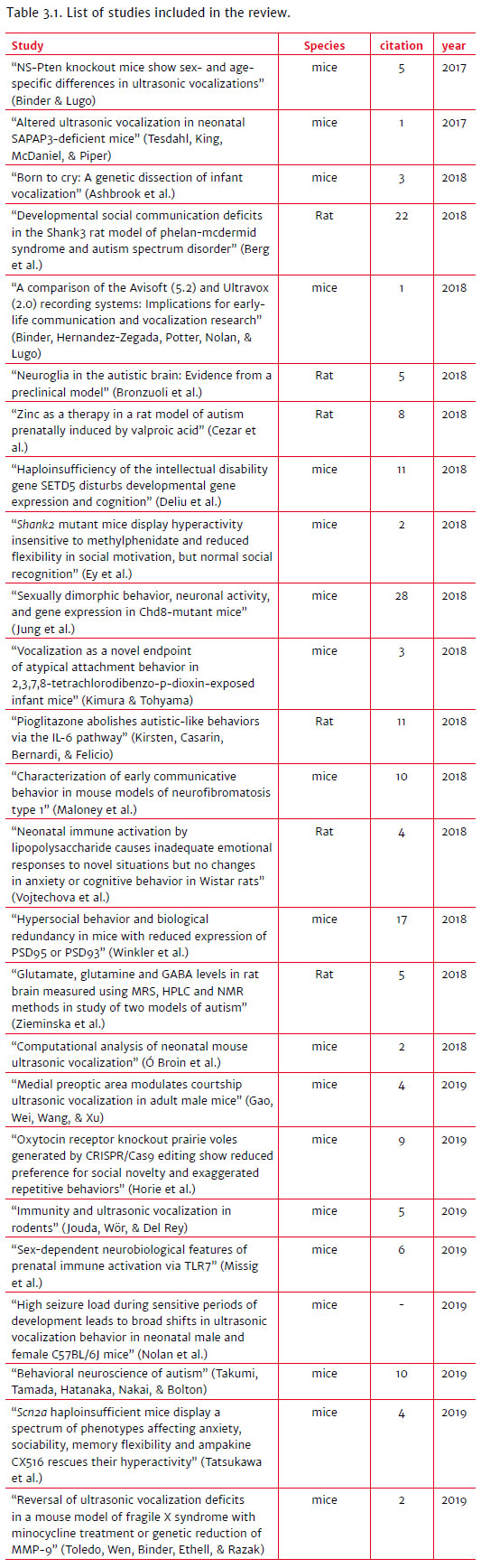
The research design (Figure 3.1) found 119 articles, 25 of which were selected for the preparation of this manuscript (Table 3.1). The described species are murine animal models, highlighting Mus musculus, Wistar rat, and one species of the genus Microtus.
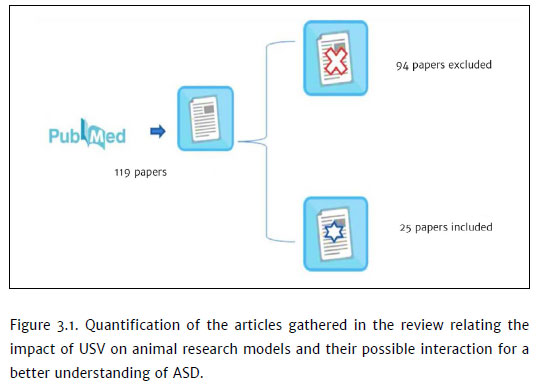
The data show 23 experimental studies and two bibliographic reviews. Studies with mice represent the majority of research, adding up to 19 studies and five studies conducted with rats, whereas one work taking advantage of the genus Microtus.
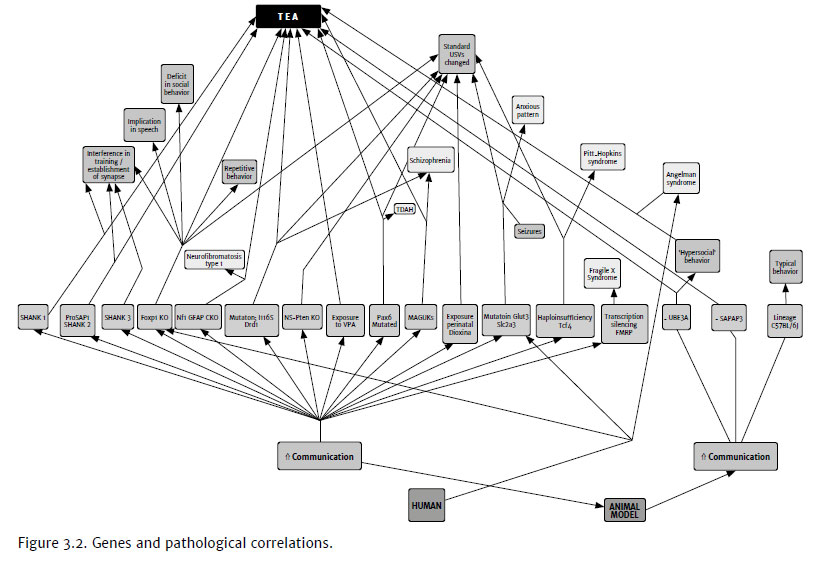
The findings in our search review corroborate the literature. In basic research, the main model used in the study of ultrasonic vocalization is the transgenic mice. The used researched literature shows that animal experimental models exist for almost all known human gene disruption. The coding regions of the genome are especially well preserved and easier to model in mice, while non-coding DNA, including regulatory elements, shows reduced conservation among various species (St. Clair & Johnstone, 2018).
Genetically modified mice (Figure 3.2) as knockouts/knock-ins were generated and carry mutations with human correlation. Genetic associations include allelic variants such as fragile X, MET tyrosine kinase, Mecp2, neurexine, and tuberosclerosis susceptibility complexes (Bishop & Lahvis, 2011).
The reduction in communication was associated with the most diverse models, and, within them, we highlight some studies such as Binder and Lugo (2017), Berg et al. (2018), Missig et al. (2019), and some genes such as the SHANK family. In recent years, deletions and coding mutations in the SHANK1 SHANK2 and SHANK3 genes have been identified in individuals with ASD. The three SHANK genes encode many variants of mRNA splicing and generate multiple protein isoforms grouped into a large molecular cluster related to the postsynaptic density at excitatory glutamatergic terminals. As precursor proteins, they bind glutamatergic receptors in the molecular assembly pathway of the actin cytoskeleton and postsynaptic signaling. Therefore, SHANK genes are strongly involved in several synaptic functions, including terminal morphogenesis, synapse formation, transport of glutamate receptors, and neuronal activation by neurotransmission (Sungur et al., 2017).
Another gene of interest to improve the understanding of ASD molecular physiopathology is Foxp1. Individuals with Foxp1 mutations are diagnosed with ASD or intellectual disability and have global developmental delay, motor deficit, and severe speech impairment. However, detailed studies of loss of gene function that investigate the contributions of Foxp1 to phenotypes relevant to neurogenesis in the developing brain have yet to be performed. Some rodent models of Foxp1 have begun to elucidate their function mainly in the adult brain (Usui et al., 2017). In contrast, some studies report an increase in USV (Tesdahl, King, McDaniel, & Pieper, 2017; Winkler et al., 2018; Nolan et al., 2019; Toledo, Wen, Binder, Ethell, & Razak, 2019).
4. Final considerations and perspectives
The acquisition of language is marked by maturation in fine aspects of brain development throughout infanthood and adolescence. This process involves different language dimensions for the development and expression of sounds, words, grammar, diction, elocution, narratives, and conversation.
It is evident, from the findings reported here, the crucial role of certain genes in modulating aspects of communication, the vast majority being markers of communication deficits. Most of the rodents of transgenic origin belong to three major groups: postsynaptic protein, synaptic cell adhesion molecules (neurexin, neuroligin, and contactin-associated protein), and genes associated with the regulation of oxytocin release.
Therefore, gene therapy based on the use of recombinant DNA can be used to repair, replace, increase, or silence essentially any gene of interest in a target cell. The advancement of techniques in proteomic research will contribute to the development of new classes and the segmentation of drugs aiming at a specific action. Other advantages of gene therapy versus small molecules include obtaining long-term clinical benefits from a single treatment.
Finally, it should be noticed that all the advances and attempts to determine more individualized and complete cellular and molecular routes in the knowledge of the pathophysiology of ASD should not mitigate the enormous success of social inclusion and pedagogical and psychotherapeutic support for individuals (in special children and adolescents) with some form of ASD. The recognition of integrative resources in several areas of health and education already defines an era of humane and social inclusion for people with neurodevelopmental communication deficits.
References
Arriaga, G., & Jarvis, E. D. (2013). Mouse vocal communication system: are ultrasounds learned or innate? Brain Lang, 124(1),96–116. doi:10.1016/j.bandl.2012.10.002
Ashbrook, D. G., Roy, S., Clifford, B. G., Riede, T., Scattoni, M. L., Heck, D. H., & Williams, R. W. (2018). Born to cry: A genetic dissection of infant vocalization. Frontiers in Behavioral Neuroscience, 12,250. doi:10.3389/fnbeh.2018.00250 [ Links ]
Berg, E. L., Copping, N. A., Rivera, J. K., Pride, M. C., Careaga, M., Bauman, M. D., Silverman, J. L. (2018). Developmental social communication deficits in the Shank3 rat model of phelan-mcdermid syndrome and autism spectrum disorder. Autism Research, 11(4),587–601. doi:10.1002/aur.1925
Binder, M. S., Hernandez-Zegada, C. J., Potter, C. T., Nolan, S. O., & Lugo, J. N. (2018). A comparison of the Avisoft (5.2) and Ultravox (2.0) recording systems: Implications for early-life communication and vocalization research. Journal of Neuroscience Methods, 309,6–12. doi:10.1016/j.jneumeth.2018.08.015
Binder, M. S., & Lugo, J. N. (2017). NS-Pten knockout mice show sex- and age-specific differences in ultrasonic vocalizations. Brain and Behavior, 7(11),e00857. doi:10.1002/brb3.857 [ Links ]
Bishop, S. L., & Lahvis, G. P. (2011). The autism diagnosis in translation: shared affect in children and mouse models of ASD. Autism Research, 4(5),317-335. doi:10.1002/aur.216 [ Links ]
Brignell, A., Chenausky, K. V., Song, H., Zhu, J., Suo, C., & Morgan, A. T. (2018). Communication interventions for autism spectrum disorder in minimally verbal children. Cochrane Database of Systematic Reviews, 11, Cd012324. doi:10.1002/14651858.CD012324.pub2 [ Links ]
Bronzuoli, M. R., Facchinetti, R., Ingrassia, D., Sarvadio, M., Schiavi, S., Steardo, L., & Scuderi, C. (2018). Neuroglia in the autistic brain: evidence from a preclinical model. Molecular Autism, 9,66. doi:10.1186/s13229-018-0254-0 [ Links ]
Cezar, L. C., Kirsten, T. B., da Fonseca, C. C. N., de Lima, A. P. N., Bernardi, M. M., & Felicio, L. F. (2018). Zinc as a therapy in a rat model of autism prenatally induced by valproic acid. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 84(Pt A),173–180. doi:10.1016/j.pnpbp.2018.02.008
Deliu, E., Arecco, N., Morandell, J., Dotter, C. P., Contreras, X., Girardot, C., & Novarino, G. (2018). Haploinsufficiency of the intellectual disability gene SETD5 disturbs developmental gene expression and cognition. Nature Neuroscience, 21(12),1717– 1727. doi:10.1038/s41593-018-0266-2
Ey, E., Torquet, N., de Chaumont, F., Levi-Strauss, J., Ferhat, A. T., Le Sourd, A. M., & Bourgeron, T. (2018). Shank2 Mutant Mice Display Hyperactivity Insensitive to Methylphenidate and Reduced Flexibility in Social Motivation, but Normal Social Recognition. Frontiers in Molecular Neuroscience, 11,365. doi:10.3389/fnmol.2018.00365 [ Links ]
Gao, S. C., Wei, Y. C., Wang, S. R., & Xu, X. H. (2019). Medial Preoptic Area Modulates Courtship Ultrasonic Vocalization in Adult Male Mice. Neuroscience Bulletin, 35(4),697–708. doi:10.1007/s12264-019-00365-w
Hanson, J. L., & Hurley, L. M. (2012). Female presence and estrous state influence mouse ultrasonic courtship vocalizations. PLoS One, 7(7),e40782. doi:10.1371/journal.pone.0040782 [ Links ]
Horie, K., Inoue, K., Suzuki, S., Adachi, S., Yada, S., Hirayama, T., & Nishimori, K. (2019). Oxytocin receptor knockout prairie voles generated by CRISPR/Cas9 editing show reduced preference for social novelty and exaggerated repetitive behaviors. Hormones and Behavior, 111,60–69. doi:10.1016/j.yhbeh.2018.10.011
Hyter, Y., Vogindroukas, I., Chelas, E. N., Paparizos, K., Kivrakidou, E., & Kaloudi, V. (2017). Differentiating autism from typical development: Preliminary findings of greek versions of a pragmatic language and social communication questionnaire. Folia Phoniatrica et Logopaedica, 69(1–2), 20–26. doi:10.1159/000479277
Jouda, J., Wohr, M., & Del Rey, A. (2019). Immunity and ultrasonic vocalization in rodents. Annals of the New York Academy of Sciences, 1437(1),68–82. doi:10.1111/nyas.13931
Jung, H., Park, H., Choi, Y., Kang, H., Lee, E., Kweon, H., & Kim, E. (2018). Sexually dimorphic behavior, neuronal activity, and gene expression in Chd8-mutant mice. Nature Neuroscience, 21(9),1218–1228. doi:10.1038/s41593-018-0208-z
Kimura, E., & Tohyama, C. (2018). Vocalization as a novel endpoint of atypical attachment behavior in 2,3,7,8-tetrachlorodibenzo-p-dioxin-exposed infant mice. Archives of Toxicology, 92(5),1741–1749. doi:10.1007/s00204-018-2176-1
Kirsten, T. B., Casarin, R. C., Bernardi, M. M., & Felicio, L. F. (2018). Pioglitazone abolishes autistic-like behaviors via the IL-6 pathway. PLoS One, 13(5),e0197060. doi:10.1371/journal.pone.0197060 [ Links ]
Lazaro, M. T., & Golshani, P. (2015). The utility of rodent models of autism spectrum disorders. Current Opinion in Neurology, 28(2),103–109. doi:10.1097/wco.0000000000000183
Liu, X. L., Zahrt, D. M., & Simms, M. D. (2018). An Interprofessional Team Approach to the Differential Diagnosis of Children with Language Disorders. Pediatric Clinics of North America, 65(1),73-90. doi:10.1016/j.pcl.2017.08.022 [ Links ]
Maloney, S. E., Chandler, K. C., Anastasaki, C., Rieger, M. A., Gutmann, D. H., & Dougherty, J. D. (2018). Characterization of early communicative behavior in mouse models of neurofibromatosis type 1. Autism Research, 11(1),44-58. doi:10.1002/aur.1853 [ Links ]
McDaniel, J., D’Ambrose Slaboch, K., & Yoder, P. (2018). A meta-analysis of the association between vocalizations and expressive language in children with autism spectrum disorder. Research in Developmental Disabilities, 72,202-213. doi:10.1016/j.ridd.2017.11.010
Missig, G., Robbins, J. O., Mokler, E. L., McCullough, K. M., Bilbo, S. D., McDougle, C. J., & Carlezon, W. A., Jr. (2019). Sex-dependent neurobiological features of prenatal immune activation via TLR7. Molecular Psychiatry. doi:10.1038/s41380-018-0346-4 [ Links ]
Mony, T. J., Hong, M., & Lee, H. J. (2018). Empathy study in rodent model of Autism Spectrum Disorders. Psychiatry Investigation, 15(2),104–110. doi:10.30773/pi.2017.06.20
Nolan, S. O., Hodges, S. L., Condon, S. M., Muhammed, I. D. A., Tomac, L. A., Binder, M. S., Lugo, J. N. (2019). High seizure load during sensitive periods of development leads to broad shifts in ultrasonic vocalization behavior in neonatal male and female C57BL/6J mice. Epilepsy Behavior, 95,26–33. doi:10.1016/j.yebeh.2019.03.037
Broin, P. Ó., Beckert, M. V., Takahashi, T., Izumi, T., Ye, K., Kang, G., ... Hiroi, N. (2018). Computational analysis of neonatal mouse ultrasonic vocalization. Current Protocols in Mouse Biology, 8(2),e46. doi:10.1002/cpmo.46 [ Links ]
Parsons, L., Cordier, R., Munro, N., Joosten, A., & Speyer, R. (2017). A systematic review of pragmatic language interventions for children with autism spectrum disorder. PLoS One, 12(4),e0172242. doi:10.1371/journal.pone.0172242 [ Links ]
Riede, T., Borgard, H. L., & Pasch, B. (2017). Laryngeal airway reconstruction indicates that rodent ultrasonic vocalizations are produced by an edge-tone mechanism. Royal Society Open Science, 4(11),170976. doi:10.1098/rsos.170976 [ Links ]
Scattoni, M. L., Crawley, J., & Ricceri, L. (2009). Ultrasonic vocalizations: A tool for behavioural phenotyping of mouse models of neurodevelopmental disorders. Neuroscience & Biobehavioral Reviews, 33(4),508–515. doi:10.1016/j.neubiorev.2008.08.003
Schuetze, M., Rohr, C. S., Dewey, D., McCrimmon, A., & Bray, S. (2017). Reinforcement Learning in Autism Spectrum Disorder. Frontiers in Psychology, 8,2035. doi:10.3389/fpsyg.2017.02035 [ Links ]
Screven, L. A., & Dent, M. L. (2019). Social isolation produces no effect on ultrasonic vocalization production in adult female CBA/CaJ mice. PLoS One, 14(3),e0213068. doi:10.1371/journal.pone.0213068 [ Links ]
St Clair, D., & Johnstone, M. (2018). Using mouse transgenic and human stem cell technologies to model genetic mutations associated with schizophrenia and autism. Philosophical Transactions of the Royal Society B Biological Science, 373(1742). doi:10.1098/rstb.2017.0037 [ Links ]
Sungur, A. O., Jochner, M. C. E., Harb, H., Kilic, A., Garn, H., Schwarting, R. K. W., & Wohr, M. (2017). Aberrant cognitive phenotypes and altered hippocampal BDNF expression related to epigenetic modifications in mice lacking the post-synaptic scaffolding protein SHANK1: Implications for autism spectrum disorder. Hippocampus, 27(8),906–919. doi:10.1002/hipo.22741
Takumi, T., Tamada, K., Hatanaka, F., Nakai, N., & Bolton, P. F. (2019). Behavioral neuroscience of autism. Neuroscience & Biobehavioral Reviews. doi:10.1016/j.neubiorev.2019.04.012 [ Links ]
Tatsukawa, T., Raveau, M., Ogiwara, I., Hattori, S., Miyamoto, H., Mazaki, E., ... Yamakawa, K. (2019). Scn2a haploinsufficient mice display a spectrum of phenotypes affecting anxiety, sociability, memory flexibility and ampakine CX516 rescues their hyperactivity. Molecular Autism, 10,15. doi:10.1186/s13229-019-0265-5 [ Links ]
Tesdahl, N. S., King, D. K., McDaniel, L. N., & Pieper, A. A. (2017). Altered ultrasonic vocalization in neonatal SAPAP3-deficient mice. NeuroReport, 28(17),1115–1118. doi:10.1097/wnr.0000000000000863
Toledo, M. A., Wen, T. H., Binder, D. K., Ethell, I. M., & Razak, K. A. (2019). Reversal of ultrasonic vocalization deficits in a mouse model of Fragile X Syndrome with minocycline treatment or genetic reduction of MMP-9. Behavioural Brain Research, 372,112068. doi:10.1016/j.bbr.2019.112068 [ Links ]
Usui, N., Araujo, D. J., Kulkarni, A., Co, M., Ellegood, J., Harper, M., ... Konopka, G. (2017). Foxp1 regulation of neonatal vocalizations via cortical development. Genes & Development, 31(20),2039–2055. doi:10.1101/gad.305037.117
Vojtechova, I., Petrasek, T., Maleninska, K., Brozka, H., Tejkalova, H., Horacek, J., ... Vales, K. (2018). Neonatal immune activation by lipopolysaccharide causes inadequate emotional responses to novel situations but no changes in anxiety or cognitive behavior in Wistar rats. Behavioural Brain Research, 349,42–53. doi:10.1016/j.bbr.2018.05.001
Wadge, H., Brewer, R., Bird, G., Toni, I., & Stolk, A. (2019). Communicative misalignment in Autism Spectrum Disorder. Cortex, 115,15–26. doi:10.1016/j.cortex.2019.01.003
Werker, J. F., & Hensch, T. K. (2015). Critical periods in speech perception: new directions. Annual Review of Psychology, 66,173–196. doi:10.1146/annurev-psych-010814-015104
Winkler, D., Daher, F., Wustefeld, L., Hammerschmidt, K., Poggi, G., Seelbach, A., ... Dere, E. (2018). Hypersocial behavior and biological redundancy in mice with reduced expression of PSD95 or PSD93. Behavioural Brain Research, 352,35–45. doi:10.1016/j.bbr.2017.02.011
Zala, S. M., Reitschmidt, D., Noll, A., Balazs, P., & Penn, D. J. (2017). Sex-dependent modulation of ultrasonic vocalizations in house mice (Mus musculus musculus). PLoS One, 12(12),e0188647. doi:10.1371/journal.pone.0188647 [ Links ]
Zieminska, E., Toczylowska, B., Diamandakis, D., Hilgier, W., Filipkowski, R. K., Polowy, R., ... Lazarewicz, J. W. (2018). Glutamate, Glutamine and GABA Levels in Rat Brain Measured Using MRS, HPLC and NMR Methods in Study of Two Models of Autism. Frontiers in Molecular Neuroscience, 11,418. doi:10.3389/fnmol.2018.00418 [ Links ]
 Correspondence:
Correspondence:
Carlos Alberto Avellaneda Penatti
Universidade Nove de Julho, Departamento de Pós-Graduação de Mestrado e Doutorado em Medicina
Rua Vergueiro, 235, 2º subsolo, Liberdade
São Paulo, SP, Brazil. CEP 01504-001
E-mail: carlospenatti@uni9.pro.br
Submission: 31/10/2018
Acceptance: 09/04/2020
Authors notes
Juciara da C. Silva, Postgraduate Program in Medicine, Nove de Julho University (Uninove); Miriam O. Ribeiro, Postgraduate Program in Developmental Disorders, Mackenzie Presbyterian University (UPM); Déborah M. dos Santos, Postgraduate Program in Medicine, Nove de Julho University (Uninove); Carlos Alberto A. Penatti, Postgraduate Program in Medicine, Nove de Julho University (Uninove).
We would like to thank Dr. Eliana Penatti for her important contribution in reviewing the English version of this manuscript.


{kind=link}