










Serviços Personalizados
Journal

artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkUniversitas Psychologica
versão impressa ISSN 1657-9267
Univ. Psychol. v.7 n.1 Bogotá abr. 2008
Búsqueda de alimento en hámsteres dorados (mesocricetus auratus): el efecto de la distancia entre múltiples fuentes de alimento
Foraging behavior in golden hamsters (mesocricetus auratus): effect of the distance among multiple patches
Felipe Cabrera*
Universidad de Guadalajara, México
ABSTRACT
The pattern of travel and the efficiency in foraging behavior was evaluated in four hamsters searching for food within an enclosure with multiple patches. Two different distances among patches were randomly arranged: Near-Patches (10 cm separation) and Distant-Patches (21.5 cm separation). Subjects obtained the food by mounting over the cylinders (stations) placed in the enclosure of 110 cm2. Results showed that in both, Near and Distant conditions, the distance between responses was longer in late stages of the trials then in early stages. Nonetheless, the most choices to adjacent stations were in Distant-Patches condition, while skips and diagonal-station choices were more frequently showed in the Near-Patches condition.
Key words authors: Travel pattern, Foraging behavior, Distance, Hamster.
Key words plus: Clinical protocols, Quality of life.
RESUMEN
Se evaluó el patrón de desplazamiento y la eficiencia en la búsqueda de alimento de cuatro hámsteres en un encierro con múltiples metas, variando la distancia entre ellas. La distancia entre las metas se manipuló en orden aleatorio, según dos condiciones: la de Metas Cercanas (10 cm entre ellas) y la condición de Metas Distantes (21.5 cm entre ellas). Los sujetos debían extraer el alimento colocado dentro de unos cilindros (estaciones) trepando a ellos en un encierro de 110 cm2. Los resultados mostraron, en ambas condiciones, que la distancia recorrida entre las subidas a las estaciones fue mayor en momentos más tardíos del ensayo. Por otro lado, aunque la proporción más alta de elecciones se centró en las estaciones adyacentes, éstas ocurrieron en mayor medida para la condición de Estaciones Distantes, y hubo un mayor número de elecciones a estaciones ubicadas diagonalmente o salteadas en la condición de Estaciones Cercanas.
Palabras clave autores: Patrón de desplazamiento, Búsqueda de alimento, Distancia, Hámster.
Palabras clave descriptores: Protocolos clínicos, Calidad de vida.
La conducta de búsqueda de alimento (foraging) ha sido definida por algunos autores como “parte de la conducta de alimentación que se lleva a cabo en ausencia de estímulos que informan a los predadores el lugar exacto de la presa” (Thomas, 1974, p. 941). De este modo, los animales en búsqueda de alimento, cuando no tienen contacto visual u olfativo directo con la presa, deben ser especialmente sensibles y recordar aquellos lugares en donde ha sido más probable encontrarlo (Dow & Lea, 1987; Stephen & Krebs, 1986). Al respecto, la evidencia experimental muestra que los organismos, tanto en espacios abiertos como en procedimientos de laboratorio, tienden a ser eficientes en la obtención del alimento, en los que se incluyen múltiples fuentes (Cabrera & Aparicio, 2006; Mellgren, 1982; Mellgren & Roper, 1986; Menzel, 1978).
El creciente interés en estudiar la búsqueda de alimento en espacios amplios ha llevado a la creación y adaptación de diversos aparatos que permiten estudiar tal o cual aspecto del fenómeno (ver Papini, Hermitte, Mustaca & Hout, 1989). El utilizar ambientes de mayor amplitud, por su parte, ha propiciado la pertinencia de registrar las rutas y trayectorias que los sujetos siguen para llegar a los lugares que posiblemente contengan alimento. Aunque estas observaciones concernientes a la conducta espacial también se han realizado en escenarios naturales, con respecto a la migración de aves (Able, 1995; Helms & Drury, 1960), peces y otras especies (Gallistel, 1990), en el ámbito de la psicología experimental han predominado algunos procedimientos que facilitan la observación en espacios considerablemente más reducidos, como es el caso de los laberintos y encierros, utilizando principalmente roedores como sujetos.
En cualquier laberinto, las trayectorias que sigue el organismo están restringidas a la forma y dirección de los brazos. Por otro lado, los procedimientos con espacios abiertos o encierros se caracterizan por no tener estas restricciones, por lo que permiten evaluar las trayectorias del organismo de manera más fina. Precisamente por tener espacios más amplios, uno de los principales usos de los espacios abiertos y encierros es el estudio y la identificación de patrones de exploración en roedores (Poucet, Chapuis, Durup & Thinus-Blanc, 1986; Renner & Seltzer, 1991). Además, por su capacidad, permite una mayor variación de procedimientos; por ejemplo, con una sola meta (Biegler & Morris, 1996; Tommasi & Vallortigara, 2000), o un gran número de ellas (Brown, DiGello, Milewski, Wilson & Kozak, 2000). Así mismo, con variaciones en la posición de los hitos y objetos ambientales (Biegler & Morris, 1996). Con estos procedimientos también se ha evaluado el aprendizaje espacial en diversas especies, incluyendo hámsteres (Poucet et al, 1986; Thinus-Blanc et al., 1987).
Para evaluar la eficiencia y el patrón de desplazamiento de hámsteres en una tarea de búsqueda de alimento sobre una superficie con múltiples metas, se construyó un encierro con una plataforma dentro del cual se colocaron 25 tubos que contenían alimento. Los sujetos tenían que recorrer la plataforma y trepar a la parte superior de los tubos para obtener la comida. Procedimientos similares (Brown & Terrinoni, 1996; Greene & Cook, 1997) han utilizado ratas como sujetos.
Método
Sujetos
Cuatro hámsteres dorados (Mesocricetus auratus) machos, de cinco meses de edad1, sirvieron como sujetos experimentales (H1, H2, H3 y H4). Los hámsteres fueron alojados en cajas habitación de acrílico (21 x 22 x 25 cm), con libre acceso a alimento y agua. Se mantuvo un ciclo de 12:12 horas de luz-oscuridad. Al inicio del experimento, todos los sujetos eran ingenuos.
Aparato
El encierro consistió en cuatro paredes de madera forradas de aluminio y piso de acero galvanizado, cubierto con pintura gris mate. El área total de la plataforma fue de 110 x 110 cm, con paredes de 42 cm de altura. En la plataforma se trazó una cuadrícula de 81 cuadrantes (nueve cuadrantes por lado), con marcador indeleble. Cada cuadrante se numeró del 1 al 9 por cada eje (“X” y “Y”), para poder registrar la posición de los sujetos en cada momento (ver Figura 1).
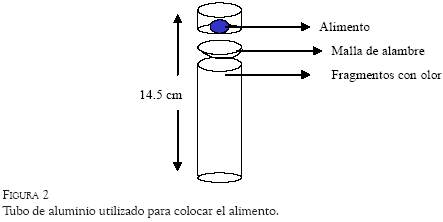
Sobre la plataforma, ubicados en el centro de cada cuadrante, se colocaron 25 tubos de aluminio de 1 pulgada de diámetro y 11 cm de altura. En la parte superior de cada tubo se dejó una pequeña concavidad de aproximadamente 1 cm de profundidad. A cada tubo se le añadió una extensión de 3.5 cm de Cloruro de Polivinilo PVC en el extremo superior, hasta alcanzar una altura total de 14.5 cm. Entre el tubo de aluminio y la extensión de PVC fue colocada una malla delgada de alambre que cubría la concavidad de la parte superior del tubo de aluminio. El alimento (una pieza de arroz inflado) se colocó dentro de la extensión de PVC sobre la malla, por lo que la profundidad para alcanzarlo fue de 3.5 cm (ver Figura 2). Pequeños fragmentos de comida se introdujeron debajo de la malla de alambre en el interior de la concavidad del tubo, con el propósito de que el olor fuera homogéneo en todos los tubos, aun después de haber sido extraída la pieza de alimento. Los 25 tubos fueron colocados en la plataforma, cada una con cinco filas de cinco tubos.
Se instalaron cuatro cortinas negras alrededor del aparato, las cuales se sostenían desde una estructura metálica tubular de 167 cm de altura, construida en torno a la caja experimental. La parte superior de esta estructura metálica fue cubierta por tablas de madera pintadas de negro, dejando descubierto un espacio para colocar la cámara de video. Ésta se montó sobre un trípode a una altura de 207 cm del piso, altura suficiente para abarcar toda el área de la plataforma. También en la parte superior de la estructura se colocó una luz blanca de 10 watts, y una bocina que producía ruido blanco. La cámara de video se conectó a un aparato de televisión a modo de circuito cerrado, para poder observar al animal desde el exterior, y a una videograbadora en la que se registraron las sesiones experimentales.
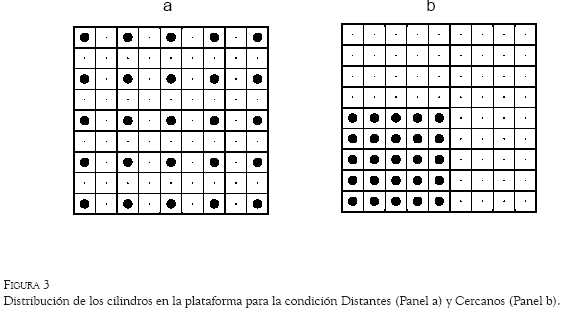
En este experimento se diseñaron dos condiciones. En la condición Distantes se colocaron los 25 tubos en cinco filas de 5 tubos cada una, guardando una distancia entre los tubos adyacentes de 21.5 cm (Figura 3a). En la condición Cercanos, el área utilizada de la plataforma se redujo a 60 x 60 cm, y la distancia entre los tubos adyacentes fue de 10 cm. Para reducir el tamaño de la superficie, se colocaron dos paredes en ángulo recto (con 60 cm de largo y 42 cm de alto cada una), en la región suroeste de la plataforma (ver Figura 3b). De esta manera, se guardaba la proporción de la distancia entre los tubos y el tamaño efectivo del área experimental.
Procedimiento
El primer día se entrenó a los sujetos para que obtuvieran el alimento dentro de los tubos de aluminio. Durante el entrenamiento, se colocó una tabla de madera en un extremo de la superficie, de manera que formaba un triángulo rectángulo junto con las paredes de la caja, mismos que formaban el ángulo recto, y la tabla de madera la hipotenusa. Dentro de esta superficie triangular, se encontraba disponible solamente un tubo que contenía una pieza de alimento. Cada sujeto fue colocado en la esquina correspondiente al ángulo recto, y permaneció el tiempo suficiente para que explorara el lugar y obtuviera la pieza de alimento. Todos los sujetos lo lograron durante la primera sesión. Al segundo día, una vez que todos los hámsteres aprendieron a subir al tubo y obtener el alimento, inició el experimento.
Los sujetos fueron sometidos a dos condiciones experimentales: la condición Distantes y la condición Cercanos. Las condiciones Distantes y Cercanos se distribuyeron aleatoriamente en 22 sesiones cada una. Las sesiones tuvieron lugar diariamente (siete sesiones por semana), y cada una constó de tres ensayos consecutivos de la misma condición (tres distantes o tres cercanos). Los ensayos terminaron cuando el hámster hubo recolectado las 25 piezas de alimento, o transcurridos 15 minutos desde el inicio del ensayo. El tiempo entre ensayos para cada sujeto fue de 10 minutos aproximadamente, durante los cuales se limpió la plataforma con una franela húmeda2, y se colocó nuevamente el alimento en los tubos vacíos. Todas las sesiones fueron videograbadas.
Análisis de datos
Las variables dependientes que se registraron en el presente experimento fueron: la trayectoria que siguieron los sujetos para obtener el alimento, la cantidad de alimento obtenido según el número de estaciones visitadas, y el tipo de transición entre una estación y la otra a lo largo del ensayo.
Resultados
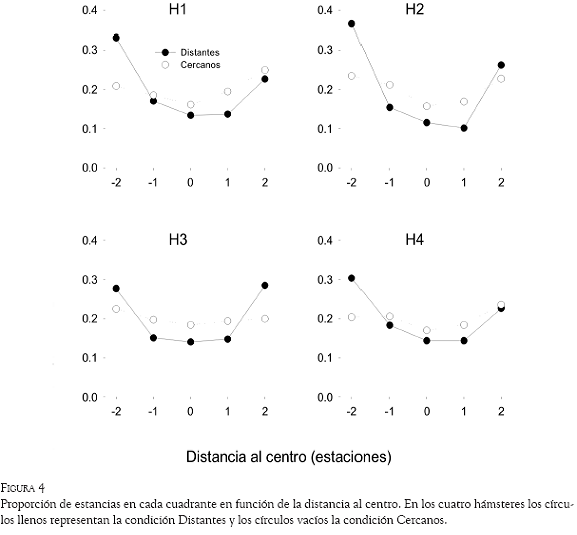
Para saber si los hámsteres al desplazarse ocupaban con más frecuencia los cuadrantes cercanos a las paredes de la plataforma que aquellos cercanos al centro, se calculó para cada sujeto la proporción de estancias en los cuadrantes en función de su distancia con respecto al centro de la plataforma (Figura 4). En el gráfico, los valores 2 y -2 en la abscisa representan los cuadrantes más cercanos a las paredes del encierro, mientras que el valor 0 representa el punto más lejano a los bordes, mismo que corresponde al centro de la plataforma. En todos los sujetos se observa que en la condición Distantes (círculos negros), la forma de la curva describe una “U”, lo que indica que la proporción de estancias en los cuadrantes más cercanos a las paredes fue mayor que en los cuadrantes centrales. En la condición Cercanos (círculos vacíos), aunque se obtuvo también una mayor proporción de estancias en los cuadrantes más próximos a los bordes, se muestra una curva más plana, lo cual señala que la tendencia a desplazarse por las orillas fue más débil.
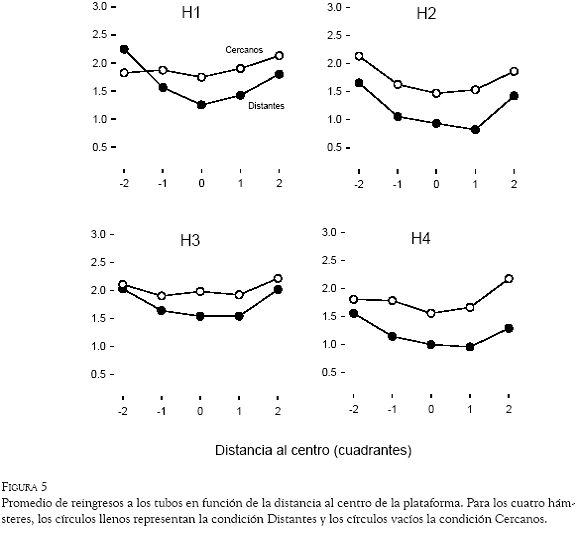
El promedio de subidas repetidas a un mismo tubo por ensayo se calculó en función de la distancia al centro de la plataforma. La Figura 5 representa, para los cuatro hámsteres, los datos obtenidos en la condición Distantes (círculos llenos) y en la condición Cercanos (círculos vacíos). Los cuatro hámsteres reincidieron en un mismo tubo en menos ocasiones durante la condición Distantes que en la condición Cercanos en todos los casos, excepto uno (H1 en la distancia al centro -2). En general, el número de subidas repetidas fue menor en las regiones centrales de la plataforma, y se incrementó en los lugares más lejanos del centro; en la condición Distantes, este resultado fue más general que en la condición de Cercanos, en la cual los sujetos H1 y H3 mostraron una curva más plana.
Las figuras 6 y 7 muestran, para las condiciones de Cercanos y Distantes respectivamente, la distancia media de desplazamiento entre dos subidas consecutivas en función del momento del ensayo (el momento del ensayo fue cuantificado por la sucesión acumulada de cuadrantes recorridos en el transcurso del ensayo, y la distancia fue calculada en cuadrantes). En la Figura 6 (condición Cercanos) se observa que, en general, al inicio del ensayo las distancias entre las subidas fueron pequeñas en los cuatro sujetos; pero conforme transcurre el ensayo, hay mayor oscilación entre distancias largas y pequeñas con una tendencia a incrementar las distancias largas. El sujeto H3 fue quien mostró mayor distancia (un máximo de 12 cuadrantes entre subidas), mientras que el sujeto H2 fue quien menor distancia recorrió (un máximo de cuatro cuadrantes).

La tendencia a incrementar la distancia recorrida entre elecciones en momentos más tardíos del ensayo se observó también en la condición Distantes (Figura 7). Sin embargo, hubo mayor estabilidad al inicio del ensayo en las distancias mínimas recorridas con respecto a la condición Cercanos. En esta condición el sujeto H2 mostró la mayor distancia recorrida entre subidas, con un máximo de 16 cuadrantes recorridos, mientras que el sujeto H4, quien menor distancia recorrió, tuvo un máximo de cuatro cuadrantes entre subidas. Nótese que la distancia mínima posible entre la elección de dos estaciones era de dos cuadrantes para la condición Cercanos (véase Figura 3a), mientras que para la condición Distantes, la distancia mínima posible era de un cuadrante (Figura 3b).

Los desplazamientos que realizaron los sujetos para elegir las distintas estaciones fueron clasificados en cuatro tipos: transición a una estación adyacente (vertical u horizontalmente), transición a una estación en diagonal, transición a alguna estación separada por al menos otra estación intermedia (salto) y, finalmente, transición en la que el sujeto regresa inmediatamente a la estación previamente elegida.
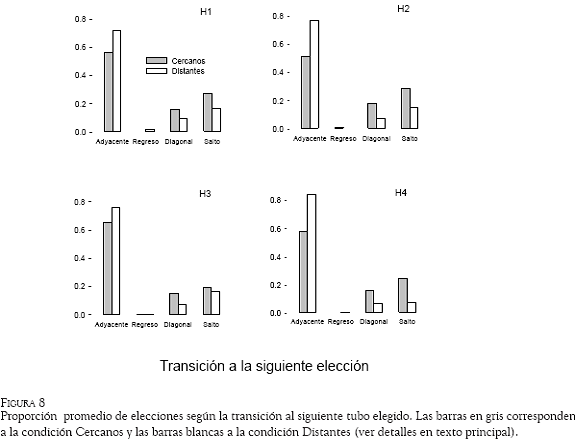
En la Figura 8 se muestra la proporción promedio de cada tipo de desplazamiento efectuado entre dos subidas consecutivas a las estaciones, para los cuatro sujetos. Las barras en gris representan los datos para la condición en la cual las estaciones se mantuvieron cercanas, mientras que las barras blancas representan la condición en la que las estaciones estuvieron distantes. En todos los sujetos, la proporción más alta de transiciones fue del tipo adyacente para ambas condiciones; sin embargo, ésta fue superior en la condición de las estaciones distantes (promedio = 7.8), con relación a la condición de estaciones cercanas (promedio = 5.9). Todos los sujetos obtuvieron mayor proporción de transiciones en diagonal y en salto con las estaciones cercanas que con las estaciones lejanas. La transición con menor proporción fue la de regreso, en la que se obtuvieron valores muy cercanos a cero en todos los sujetos.
Las transiciones de tipo adyacente (Figura 8, barras a la izquierda) pueden ocurrir de manera sucesiva desde una sola transición hasta un máximo de cuatro en cadena (el número de estaciones disponibles por línea es de cinco). En la Figura 9 se muestra, para la condición Cercanos (panel izquierdo) y la condición Distantes (panel derecho), la proporción de cadenas de transiciones adyacentes de longitudes iguales a 1, 2, 3, y 4 por ensayo, en función del momento en el que ocurren. Las cadenas de una sola transición están representadas por líneas con círculos vacíos; las de dos transiciones, por círculos negros, las de tres, por los triángulos vacíos; y las de cuatro transiciones sucesivas, con los triángulos negros.

Para la condición Cercanos (panel izquierdo), las secuencias de solamente una transición son las de mayor proporción de ocurrencia en todos los sujetos desde los primeros momentos del ensayo, mostrando una función bitónica a lo largo del ensayo. La proporción para el resto de las cadenas es notablemente inferior; la secuencia de dos transiciones es la de mayor proporción (ver principalmente sujeto H4), y disminuye a lo largo del ensayo. Por otro lado, en la condición Distantes (panel derecho), a excepción de un sujeto (H1), que tuvo la mayor proporción de una sola transición adyacente, tres sujetos muestran que las cadenas de cuatro transiciones adyacentes sucesivas ocurrieron en mayor proporción (sujetos H2 y H3), o igual (sujeto H4), a las cadenas de una sola transición en los primeros momentos del ensayo. En momentos posteriores del ensayo, las cadenas de longitud igual a 1 ocurrieron en mayor proporción mostrando, al igual que durante la condición Cercanos, una función bitónica en “U” invertida.
Discusión
Los resultados mostraron cierta similitud en la estrategia de búsqueda y obtención de alimento en las dos condiciones experimentales empleadas. Sin embargo, algunas diferencias fueron notables según el tamaño del espacio experimental y la distancia entre las metas. En la condición Distantes, los hámsteres mostraron una fuerte preferencia por el alimento cercano a las paredes en los momentos iniciales del ensayo, y posteriormente obtuvieron el alimento ubicado en los lugares más centrales de la plataforma. Por el contrario, en la condición Cercanos, las estancias en el centro de la plataforma fueron más abundantes que en la condición Distantes. El sesgo espacial hacia los bordes en lugares amplios ha sido observado habitualmente en roedores (Barnett, 1963), y se le ha asociado frecuentemente como un mecanismo de protección de predadores. Esta tendencia a desplazarse junto a los bordes del encierro en la condición Distantes facilitó que las elecciones de tubos adyacentes ocurrieran en secuencias a lo largo de un eje (columnas o filas) en mayor medida que aquellas ocurridas en la condición Cercanos. En esta última condición, las transiciones en diagonal y en salto tuvieron mayor proporción que en la condición Distantes.
El predominio de las elecciones adyacentes en este experimento es consistente con resultados de estudios obtenidos con ratas como sujetos (Brown et al, 2000; Brown & Terrinoni, 1996; Brown, Zeiler & John, 2001). Esta secuencia de elecciones adyacentes ha podido ser parcialmente alterada por Brown et al. (2001), colocando el alimento de manera alternada en los tubos o en disposición diagonal, arguyendo la formación de una representación del espacio.
Una explicación al hallazgo de las elecciones adyacentes es que, en momentos tempranos del ensayo, cuando aún todos los tubos contienen alimento, conforme el hámster se desplaza se va encontrando con los tubos en secuencia, siguiendo el contorno del encierro en la condición Distantes, y frecuentando regiones centrales en la condición Cercanos. De este modo, los sujetos trepan a los tubos que guardan mayor cercanía entre sí (elecciones adyacentes). En términos de las teorías de optimación en la búsqueda de alimento (ver Stephens & Krebs, 1986), se trata de una situación de elección de presas, en la cual la tarea consiste en una repetición de la secuencia: búsqueda, encuentro, y toma de decisión. La decisión que se toma en este caso es entre interrumpir o seguir buscando el alimento3 (Getty, Kamil & Real, 1987; Stephens & Krebs, 1986). Cuando las presas están dispersas en un espacio, las más cercanas tendrán mayor valor que las lejanas (Stephens & Krebs, 1986). En este experimento, los tubos adyacentes proveen mayor beneficio por su cercanía, y tienen ventaja sobre tubos más alejados; por esta razón la distancia entre subidas es muy cercana a la mínima posible en todos los hámsteres al inicio de los ensayos. En el caso de la condición Distantes, este factor se conjuga con la tendencia a seguir los bordes del encierro, de tal manera que las secuencias de elecciones adyacentes son más largas que en la condición Cercanos.
Conforme transcurre el ensayo y el sujeto captura el alimento, se altera la distribución y cantidad de alimento disponible en el encierro, de modo que es cada vez más escaso. En estos momentos tardíos del ensayo, los hámsteres tienden a recorrer mayores distancias antes de subir a otro tubo. En este momento los tubos cercanos dejan de ser los más provechosos (Stephens & Krebs, 1986), y los hámsteres dejan de trepar a aquellos cuyo alimento ha sido agotado. Este resultado corresponde a las descripciones sobre memoria espacial a corto plazo en estudios sobre optimación de la búsqueda del alimento (Armstrong, Gass & Sutherland, 1987), en los cuales el animal evita los lugares en los que ya consumió el alimento (Olton & Samuelson, 1976), y acude a aquellos que aún no ha visitado y, por tanto, pueden proveer mayor beneficio. Tal explicación es compatible con el modelo de difusión espacial de Reid y Staddon (1998), en el que se asume que los organismos elegirán aquellos lugares que localmente proveen mayor ganancia (ver Staddon & Reid, 1990), mostrando un gradiente de desplazamiento en el que los sujetos se dirigirán a los lugares en donde aún existe probabilidad de encontrar comida.
Sin embargo, a pesar de que los hámsteres subían cada vez menos a los tubos en donde ya habían consumido el alimento, no cesó el número de reingresos, principalmente en las regiones cercanas a los bordes. Ello indica, por un lado, que los sujetos, al desplazarse por las orillas, tendían a trepar a algunos de los tubos que encontraban a su paso, a pesar de que ya no contenían alimento; por otro lado, muestra que los sujetos no pudieron discriminar óptimamente entre aquellos tubos ya visitados y los aún sin visitar. Un factor responsable de este resultado pudo ser la cantidad de tubos presentes en la situación; según Olton (1982), por ejemplo, la cantidad de información que puede ser recordada en ratas es de aproximadamente 12 lugares o hasta 15, utilizando laberintos radiales con 17 brazos (Olton, Collison & Werz, 1977; Wilkie & Slobin, 1983). En este experimento, en cambio, el número de lugares ascendió a 25, lo que representa una mayor carga en memoria (Olton, 1982).
Otro factor que pudo haber influido en este resultado fue la eliminación de los hitos extralaberinto, al circundarse la caja con cortinas negras. En muchos estudios sobre memoria espacial, los sujetos tienen acceso a hitos extralaberinto (Suzuky, Augerinos & Black, 1980), así como a ciertas señales dentro del laberinto (Olton & Samuelson, 1976), que facilitan el recuerdo de aquellos lugares a donde ya ingresaron anteriormente. Se ha observado en hámsteres una tendencia a guiarse por estímulos visuales, cuando éstos son contextuales o extralaberinto (Etienne, Joris, Maurer & Teroni, 1990). Estudios en los que se eliminan los hitos extralaberinto (Brown & Moore, 1997) han mostrado que algunos sujetos desarrollan estrategias más rígidas y estereotipadas de desplazamiento, como el ingresar a los brazos adyacentes en cada elección, cuando se utilizan laberintos radiales (Yoerg & Kamil, 1982).
Referencias
Able, K. P. (1995). Orientation and Navigation: A Perspective on Fifty Years of Research. The Condor, 97, 592-604. [ Links ]
Armstrong, D. P., Gass, C. L. & Sutherland, G. D. (1987). Should Foragers Remember where Theyve Been? Explorations of a Simulation Model Based on the Behavior and Energetics of Territorial Hummingbirds. En A. C. Kamil, J. R. Krebs & H. R. Pulliam (Eds.), Foraging behavior (pp. 563-586). New York: Plenum Press. [ Links ]
Barnett, S.A. (1963). The Rat: A Study in Behaviour. London: Methuen. [ Links ]
Bartke, A. (1985). Male Hamster Reproductive Endocrinology. En H. I. Siegel (Ed.), The Hamster: Reproduction and Behavior (pp. 73-98). New York: Plenum Press. [ Links ]
Biegler, R. & Morris, G. M. (1996). Landmark Stability: Studies Exploring whether the Perceived Stability of the Environment Influences Spatial Representation. The Journal of Experimental Biology, 199, 187-193. [ Links ]
Brown, M. F., DiGello, E., Milewski, M., Wilson, M. & Kozak, M. (2000). Spatial Pattern Learning in Rats: Conditional Control by Two Patterns. Animal Learning and Behavior, 28, 278-287. [ Links ]
Brown, M. F. & Moore, J. A. (1997). In the Dark II: Spatial Choice when Access to Extrinsic Spatial Cues is Eliminated. Animal Learning and Behavior, 25, 335-346. [ Links ]
Brown, M. F. & Terrinoni, M. (1996). Control of Choice by the Spatial Configuration of Goals. Journal of Experimental Psychology: Animal Behavior Processes, 22, 438-446. [ Links ]
Brown, M. F., Zeiler, C. & John, A. (2001). Spatial Pattern Learning in Rats: Control by an Iterative Pattern. Journal of Experimental Psychology: Animal Behavior Processes, 27, 407-416. [ Links ]
Cabrera, F. & Aparicio, C. F. (2006). Travel, Sensitivity to Reinforcement, and Multiple Alternatives. Brazilian Journal of Behavior Analysis, 2, 1-14. [ Links ]
Davis, S. F., Kring, J. P. & Richardson, T. R. (1999). Endogenous Odor Control of Animal Runway Performance: Generality, Questions, and Utility. The Psychological Record, 49, 397-408. [ Links ]
Dow, S. M., & Lea, S. E. G. (1987). Foraging in a Changing Environment: Simulations in the Operant Laboratory. En M. L. Commons., A. Kacelnik., & S. J. Shettleworth (Eds.), Quantitative Analyses of Behavior: Vol. VI Foraging (pp. 89-113). New Jersey: Lawrence Erlbaum Associates, Publishers. [ Links ]
Etienne, A. S., Joris, S., Maurer, R. & Teroni, E. (1990). Enhancing the Impact of Visual Extra-maze Cues in a Spatial Orientation Task. Behavioural Brain Research, 38, 199-210. [ Links ]
Gallistel, C. R. (1990). The Organization of Learning. Cambridge: MIT Press. [ Links ]
Getty, T., Kamil, A. C. & Real, P. G. (1987). Signal Detection Theory and Foraging for Cryptic or Mimetic Prey. En A.C. Kamil, J. R. Krebs & H. R. Pulliam (Eds.), Foraging behavior (pp. 525-548). New York: Plenum Press. [ Links ]
Greene, C. M. & Cook, R. G. (1997). Landmark Geometry and Identity Controls Spatial Navigation in Rats. Animal Learning and Behavior, 25, 312-323. [ Links ]
Helms, C. W. & Drury, W. H. (1960). Winter and Migratory Weight and Fat Field Studies on some North American Buntings. Journal of Ornithological Investigation, 31, 1-40. [ Links ]
Johnston, R. E. (1985). Communication. En H. I. Siegel (Ed.), The Hamster: Reproduction and Behavior (pp. 121-154). New York: Plenum Press. [ Links ]
Mellgren, R. L. (1982). Foraging in a Simulated Natural Environment: Theres a Rat Loose in the Lab. Journal of the Experimental Analysis of Behavior, 38, 93-100. [ Links ]
Mellgren, R. L. & Roper, T. J. (1986). Spatial Learning and Discrimination of Food Patches in the European Badger (Meles meles L.). Animal Behavior, 34, 1129-1134. [ Links ]
Menzel, E. W. (1978). Cognitive Mapping in Chimpanzees. En S. H. Hulse, H. Fowler & W. K. Honig (Eds.), Cognitive Processes in Animal Behavior (pp. 375-422). New Jersey: Lawrence Erlbaum Associates. [ Links ]
Olton, D. S. (1982). Staying and Shifting. Their Effects on Discrimination Learning. En M. L. Commons, R. J. Herrnstein y H. Rachlin (Eds.), Quantitative Analyses of Behavior: Vol. II. Matching and maximizing accounts (pp. 205-225). Cambridge: Ballinger Publishing Company. [ Links ]
Olton, D. S., Collison, C. & Werz, A. (1977). Spatial Memory and Radial Arm Maze Performance of Rats. Learning and Motivation, 8, 289-314. [ Links ]
Olton, D. S. & Samuelson, R. J. (1976). Remembrance of Places Passed: Spatial Memory in Rats. Journal of Experimental Psychology: Animal Behavior Processes, 2, 97-116. [ Links ]
Papini, M. R., Hermitte, G., Mustaca, A. E. & Hout, G. (1989). Aprendizaje espacial en mamíferos marsupiales y placentarios. Revista Latinoamericana de Psicología, 21, 57-66. [ Links ]
Poucet, B., Chapuis, N., Durup, M. & Thinus-Blanc, C. (1986). A Study of Exploratory Behavior as an Index of Spatial Knowledge in Hamsters. Animal Learning and Behavior, 14, 93-100. [ Links ]
Reid, A. K. & Staddon, J. E. R. (1998). A Dynamic Route Finder for the Cognitive Map. Psychological Review, 105, 585-601. [ Links ]
Renner, M. J. & Seltzer, C. P. (1991). Molar Characteristics of Exploratory and Investigatory Behavior in the Rat (Rattus novergicus). Journal of Comparative Psychology, 105, 326-339. [ Links ]
Staddon, J. E. R. & Reid, A. K. (1990). On Dynamics of Generalization. Psychological Review, 97, 576-578. [ Links ]
Stephens, D. W. & Krebs, J. R. (1986). Foraging Theory. Princeton: Princeton University Press. [ Links ]
Suzuki, S., Augerinos, G. & Black, A.H. (1980). Stimulus Control of Spatial Behavior on the Eight-arm Maze in Rats. Learning and Motivation, 11, 1-18. [ Links ]
Thinus-Blanc, C., Bouzouba, K., Chaix, N., Chapuis, M., Durup, M. & Poucet, B. (1987). A Study of Spatial Parameters Encoded During Exploration in Hamsters. Journal of Experimental Psychology: Animal Behavioral Processes, 13, 418-427. [ Links ]
Thomas, G. (1974). The Influences of Encountering a Food Object on Subsequent Searching Behaviour in gasterosteus aculeatus L. Animal Behaviour, 22, 941-952. [ Links ]
Tommasi, L. & Vallortigara, G. (2000). Searching for the Center: Spatial Cognition in the Domestic Chick (Gallus gallus). Journal of Experimental Psychology: Animal Behavior Processes, 26, 447-486. [ Links ]
Wilkie, D. M. & Slobin, P. (1983). Gerbils in Space: Performance on the 17-arm Radial Maze. Journal of the Experimental Analysis of Behavior, 40, 301-312. [ Links ]
Yoerg, S. I. & Kamil, A. C. (1982). Response Strategies in the Radial Arm Maze: Running around in Circles. Animal Learning and Behavior, 10, 530-534. [ Links ]
Recibido: Septiembre 12 de 2007
Revisado: Febrero 1 de 2008
Aceptado: Febrero 3 de 2008
*Universidad de Guadalajara, CEIC, Francisco de Quevedo 180, Col Arcos Vallarta CP 44130, Guadalajara, Jalisco, México. AP 5-374, Correo electrónico: fcabrera@cencar.udg.mx
1Los hámsteres alcanzan su madurez sexual a los 50-55 días de nacidos (Bartke, 1985).
2Limpiar la superficie con un trapo húmedo evita la permanencia de olores en el espacio que puedan sesgar la ubicación de los hámsteres (ver Davis, Kring & Richardson, 1999; Johnston, 1985).
3En la literatura sobre conducta de búsqueda de alimento, se hace la distinción entre el modelo de elección de presas y el de elección de zonas (patches). La elección de presas asume un cambio abrupto en el momento de tomar la presa y seguir buscando otra; la elección de zonas supone un agotamiento gradual de la fuente de energía, por lo que la toma de decisión en este caso consiste en cuándo abandonar una zona y trasladarse a otra (Stephens & Krebs, 1986).

