










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkInteramerican Journal of Psychology
versão impressa ISSN 0034-9690
Interam. j. psychol. v.41 n.3 Porto Alegre dez. 2007
Estimação temporal de ratos com lesão hipocampal em discriminação condicional operante1
Temporal estimation of rats with hippocampal lesion in operant conditional discrimination
José Lino Oliveira Bueno2; Lézio Soares Bueno Júnior
Universidade de São Paulo, Ribeirão Preto, Brasil
RESUMO
O desempenho de ratos com lesão no giro denteado do hipocampo foi avaliado durante tarefa operante de discriminação condicional com manipulação da duração do estímulo-característica. Foram usados 16 ratos privados de água: oito lesados no giro denteado com microinjeções de colchicina e oito controle-operados. Os ratos foram submetidos a treino de discriminação condicional positiva (práticas L15 T5+ e T5-) e negativa (L5R15- e R15+), sendo L estímulo-característica luz branca, T (tom puro) e R (ruído branco) estímulos-alvo, 5 e 15 durações em segundos, seta () intervalo vazio de 5 s e + e - presença e ausência de reforçamento, respectivamente. O teste empregou práticas de extinção, com durações de L de 15, 13, 11, 9, 7 e 5 s. Os dados mostraram aquisição da discriminação condicional positiva e negativa, sem diferenças entre o grupo experimental e o controle. Diferentes durações de um estímulo podem adquirir propriedades condicionais. Contudo, o processamento da duração do estímulo-característica parece não depender somente do giro denteado do hipocampo.
T5+ e T5-) e negativa (L5R15- e R15+), sendo L estímulo-característica luz branca, T (tom puro) e R (ruído branco) estímulos-alvo, 5 e 15 durações em segundos, seta () intervalo vazio de 5 s e + e - presença e ausência de reforçamento, respectivamente. O teste empregou práticas de extinção, com durações de L de 15, 13, 11, 9, 7 e 5 s. Os dados mostraram aquisição da discriminação condicional positiva e negativa, sem diferenças entre o grupo experimental e o controle. Diferentes durações de um estímulo podem adquirir propriedades condicionais. Contudo, o processamento da duração do estímulo-característica parece não depender somente do giro denteado do hipocampo.
Palavras-chave: Estimação temporal, Discriminação condicional, Hipocampo, Lesão, Ratos.
ABSTRACT
The performance of rats with hippocampus´ dentate gyrus lesion was evaluated during an operant conditional discrimination task with manipulation of the duration of the feature-stimulus. Sixteen rats under water deprivation were used: eight lesioned in the dentate gyrus by colchicine microinjections and eight sham-operated. The rats were submitted to a conditional discrimination training with feature-positive practices (L15T5+ and T5-) and feature-negative practices (L5R15- and R15+), where L is feature-stimulus white light, T (pure tone) and R (white noise) are target-stimuli, 5 and 15 are durations in seconds, arrow () is a 5 s empty interval and + and - are presence and absence of reinforcement, respectively. In the test, practices in extinction were used, with L durations of 15, 13, 11, 9, 7 and 5 s. The data indicated acquisition of positive and negative conditional discrimination, without differences between the experimental group and the control group. Different durations of a stimulus can acquire conditional properties. However, the feature-stimulus duration processing apparently do not depends on the hippocampus´ dentate gyrus alone.
Keywords: Temporal estimation, Conditional discrimination, Hippocampus, Lesion, Rats.
Jenkins e Sainsbury (1969) realizaram procedimentos pavlovianos de discriminação condicional positiva que consistiram de: 1. apresentação de dois estímulos (X e A) formando um composto simultâneo (XA) seguido de estímulo incondicionado, como água ou comida (XA+); e 2. apresentação do estímulo A sozinho e não seguido de estímulo incondicionado (A-). Os resultados mostraram que, dependendo da condição em que A é apresentado, se acompanhado de X ou não, o estímulo A pode adquirir, respectivamente, propriedade excitatória ou inibitória. De forma semelhante, na discriminação condicional negativa, XA não é seguido de liberação de água ou comida (XA-), enquanto A sozinho é seguido da liberação de água ou comida (A+). Porém, na discriminação condicional negativa, A adquire propriedade inibitória se acompanhado de X e propriedade excitatória se apresentado sozinho. Tanto na discriminação condicional positiva quanto na negativa, X é denominado estímulo condicional ou estímulo-característica e A é denominado estímulo condicionado ou estímulo-alvo.
Os arranjos envolvendo os estímulos X e A podem ser simultâneos ou seriados. Ross e Holland (1981), utilizando arranjos simultâneos (XA+/A-) em um procedimento pavloviano com registro de alterações em diferentes categorias de comportamentos, fizeram um teste cujo resultado mostrou que X não adquire a propriedade de estímulo-característica, ou estímulo condicional, em arranjos simultâneos. Em um segundo experimento no mesmo trabalho, utilizou-se um procedimento pavloviano de discriminação condicional positiva em que X e A foram apresentados em série e separados por um intervalo vazio (XA). Neste procedimento, a série XA foi seguida de estímulo incondicionado (XA+) enquanto A apresentado sozinho não foi seguido de estímulo incondicionado (A-). Os dados da fase de trei no mostraram que a freqüência de respostas de orientação ao comedouro durante A precedido por X foi maior do que durante A apresentado sozinho. Num teste de transferência, a substituição de A por um estímulo B prejudicou a discriminação durante o estímulo-alvo, o que indicou que em procedimentos seriados ocorre associação básica do estímulo-alvo com o estímulo incondicionado, enquanto o estímulo X define a ocasião (occasion setting) durante a qual A será seguido de estímulo incondicionado.
A propriedade inibitória que um estímulo-característica pode adquirir durante treino de discriminação condicional negativa seriada foi examinada por diversos autores (Holland, 1984; Holland, 1985; Pearce & Hall, 1980; Rescorla & Holland, 1977; e Wagner & Rescorla, 1972). Holland e Lamarre (1984) submeteram animais ao procedimento simultâneo (XA- e A+) e seriado (XA- e A+) e mostraram que a substituição de A por um estímulo B durante o teste prejudicou a discriminação somente de animais treinados com procedimento seriado. Deste modo, em discriminação condicional negativa seriada, X sinaliza a ausência de estímulo incondicionado após a apresentação do estímulo-alvo, já que a freqüência de respostas após A apresentado sozinho é maior do que a freqüência de respostas após A precedido por X.
Na relação do animal com o ambiente, os estímulos não são definidos somente pelas características físicas detectadas pelo aparato sensorial, mas também pelo processamento subseqüente no sistema nervoso central (Gibson, 1960). Apesar de os receptores sensoriais serem sensíveis a muitos aspectos de um estímulo, somente alguns destes aspectos podem ser relevantes para o processamento da resposta comportamental. Este processo mediador do sistema nervoso central gera as representações dos estímulos externos (Lawrence, 1963).
Um animal possui uma representação se ele pode usar uma informação adquirida do ambiente, mesmo que aquela informação esteja ausente naquele momento (Gallistel, 1990). Esta capacidade cognitiva possibilita uma flexibilidade comportamental importante diante das mudanças imprevistas do meio (Vauclair, 1996). Uma representação corresponde a uma espécie de cópia de um estímulo, armazenada e topograficamente organizada em áreas sensoriais do cérebro. É o caso das representações modais, originadas de estímulos luminosos, sonoros, etc.
Por outro lado, animais com cérebros que possuem áreas associativas, além das áreas sensoriais, também são capazes de produzir representações amodais dos estímulos. As representações amodais resultariam da abstração a partir de atributos físicos dos estímulos, independentemente das modalidades dos estímulos que as originaram (Bueno, 1997). Meck e Church (1982) tornaram a idéia consistente ao demonstrarem que ratos são capazes de transferir a representação de uma duração adquirida com um estímulo lumi noso para um estímulo sonoro. Neste caso, a representação amodal da duração mostrou-se independente da modalidade luminosa, que a originou. Isto indica, portanto, que ratos podem processar representações amodais, como as durações.
Componentes temporais em procedimentos de discriminação condicional pavloviana foram examinados com a manipulação da duração do intervalo vazio por Holland (1998). Os efeitos de durações de estímulos em um procedimento pavloviano de discriminação condicional também foram examinados por Bueno, Wagatsuma e Martins (2007). Diferentemente de Holland (1998), o objetivo desse estudo foi investigar se diferentes durações de um estímulo-característica de mesma modalidade podem adquirir propriedades excitatórias ou inibitórias. No trabalho de Bueno et al. (2007) o procedimento pavloviano de discriminação condicional positiva envolveu dois tipos de práticas: 1. a apresentação de luz (L) como estímulo-característica com duração de 15 s (L15) seguida de um intervalo vazio de 5 s () precedendo um tom como estímulo-alvo (T) de 5 s (T5) e seguido de estímulo incondicionado (L15T5+); e 2. a apresentação de um tom de 5 s sozinho e sem estímulo incondicionado (T5-). Durante as mesmas sessões, os mesmos ratos também foram treinados em práticas de discriminação condicional negativa, com L5R15- (onde R é ruído branco) e R15+. Os resultados do treino mostraram que a freqüência de respostas durante T5 precedido por L15 foi maior do que a freqüência de respostas durante T5 sozinho. Além disto, a taxa de respostas de orientação ao bebedouro durante R15 sozinho foi maior do que durante R15 precedido por L5. Estes resultados indicam a aquisição de propriedade excitatória pela luz de 15 s e inibitória pela luz de 5 s. Portanto, uma mesma modalidade de estímulo, como a luz, pode adquirir propriedade excitatória e inibitória dependendo da duração com que é apresentada em treino de discriminação condicional positiva e negativa, respectivamente.
Os efeitos de durações de estímulos em discriminação condicional também foram examinados com procedimentos de condicionamento operante. Martins (2001) empregou o mesmo treino usado por Bueno et al. (2007), porém utilizando um procedimento em que as respostas medidas foram pressões à barra. Deste modo, era liberado um estímulo reforçador contingente a pressões à barra emitidas durante os estímulos-alvo. Após o treino, os animais foram testados em práticas em que os estímulos-alvo foram precedidos de diferentes durações de estímulos-característica, intermediárias entre as empregadas no treino (L15T5-, L13T5-, L11T5-, L9T5-, L7T5-, L5T5-, L15R15-, L13R15-, L11R15-, L9R15-, L7R15- e L5R15-). Os resultados do teste mostraram que a freqüência de respostas durante a apresentação do estímulo-alvo variou conforme a duração do estímulo-característica precedente. A freqüência de respostas emitidas durante T5 foi significativa mente maior quando este estímulo-alvo era precedido por L15, L13 e L11, em comparação com L9, L7 e L5. Estes resultados indicam a aquisição de propriedade excitatória por L com duração próxima ou igual a 15 s, pois esta duração do estímulo-característica foi empregada no treino de discriminação condicional positiva. Além disto, foi demonstrada a aquisição da propriedade inibitória por L com duração próxima ou igual a 5 s, já que esta duração foi empregada no treino de discriminação condicional negativa.
Os resultados de Bueno et al. (2007) e Martins (2001) indicam a aquisição de propriedade excitatória pela luz de 15 s e inibitória pela luz de 5 s, tanto com o emprego de um procedimento pavloviano (Bueno et al., 2007) quanto de um procedimento operante (Martins, 2001). Portanto, um estímulo de mesma modalidade, como a luz, pode adquirir para um mesmo animal e em condições experimentais semelhantes, propriedade excitatória e inibitória dependendo da duração com que é apresentada em treinamento de discriminação condicional positiva e negativa, respectivamente. De modo geral, os resultados dos dois trabalhos mostram que a manipulação de durações de estímulos afeta o desempenho de ratos em procedimentos de aprendizagem complexa.
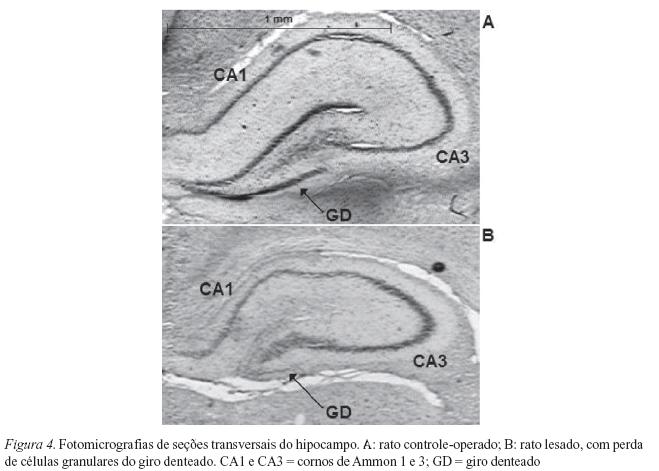
A formação hipocampal é composta por uma série de estruturas interconectadas: córtex entorrinal, giro denteado, cornos de Ammon (CA) e subiculum. As células granulares do giro denteado exercem importante papel no fluxo de informações que ocorre dentro do hipocampo, pois recebem impulsos excitatórios do córtex entorrinal e ativam, direta e indiretamente, os cornos de Ammon (CA). Goldschmidt e Steward (1980, 1982) demonstraram que injeções de colchicina no giro denteado do hipocampo produzem eliminação seletiva de suas células granulares, causando danos insignificantes em outras regiões hipocampais. A colchicina se liga às tubulinas no interior dessas células bloqueando o processo mitótico e o transporte de moléculas através dos axônios (Hanson & Estrom, 1978).
Ratos com lesões hipocampais mostram deficiência no desempenho de diversas tarefas. Estudos com lesões seletivas do hipocampo têm indicado que suas diferentes regiões podem estar envolvidas em processos comportamentais distintos (Xavier, Oliveira-Filho & Santos, 1999). Costa, Bueno e Xavier (2005) e Bueno, Costa, Xavier e Dima (2006) realizaram experimentos envolvendo lesão seletiva do giro denteado do hipocampo por aplicações de colchicina. Esses estudos mostraram que há participação das células granulares do giro denteado do hipocampo no processamento necessário ao desempenho de ratos em tarefas temporais. Nesses trabalhos, ratos foram treinados em um procedimento de DRL (differential reinforcement of low rate), uma tarefa em que uma pressão à barra só é reforçada se separada da resposta anterior por determinado intervalo de tempo. Costa et al. (2005) realizaram o experimento com cirurgia de lesão no giro denteado entre a fase de treino e a de teste de DRL, enquanto Bueno et al. (2006) realizaram as cirurgias antes do treino de DRL. Os dados do teste pós-lesão de Costa et al. (2005) indicaram que os animais com lesão no giro denteado mantiveram a capacidade de espaçar suas respostas por um determinado intervalo. Porém, em relação aos controle-operados, os animais lesados encurtaram para cerca de 16 s a duração do intervalo treinado antes das cirurgias, que foi de 20 s. Os dados da aquisição de DRL de Bueno et al. (2006) também indicaram prejuízo no desempenho de ratos lesados no giro denteado do hipocampo.
Considerando-se que a lesão do giro denteado do hipocampo prejudicou o desempenho de ratos submetidos a esquemas temporais de reforçamento durante tarefa de DRL, podem ser úteis procedimentos que também acessem a estimação de durações de estímulos em aprendizagem complexa. Moreira e Bueno (2003) mostraram que a lesão neonatal das células granulares do giro denteado hipocampal não afeta o desempenho de ratos em tarefas de discriminação condicional, em procedimentos que não envolvem manipulação da duração dos estímulos-característica ou estímulos-alvo. Entretanto, a lesão destas células pode afetar o desempenho do animal em procedimentos de discriminação condicional em que se dá a manipulação das durações de estímulos envolvidos, uma vez que estas células parecem ser importantes para o desempenho em tarefas temporais, como o DRL (Costa et al., 2005 e Bueno et al., 2006).
O presente estudo visou à avaliação do desempenho de ratos com lesão do giro denteado do hipocampo durante uma tarefa de discriminação condicional com manipulação da duração do estímulo-característica. Ratos lesados e não lesados foram submetidos a sessões de treino com o emprego de estímulos-característica de durações diferentes, associadas respectivamente a propriedades positiva ou negativa, a fim de se investigar a relação do giro dentado do hipocampo com a representação de durações.
Material e Método
Sujeitos
Foram utilizados 16 ratos Wistar adultos machos provenientes do biotério central do campus de Ribeirão Preto da Universidade de São Paulo, todos experimentalmente ingênuos, com 90 dias de idade no início do experimento e pesando, inicialmente, de 200 a 250 g. Durante todo o experimento, os animais foram mantidos em gaiolas individuais dentro de um biotério com ciclo diário de iluminação das 6:00 às 18:00. O peso dos animais foi mantido em 85% do peso estável durante todo o experimento pela privação de água. O lote foi dividido, por sorteio, em dois grupos: o de animais a serem lesados e o de animais a sofrerem cirurgia falsa (controle-operados).
Equipamento
Foram empregadas quatro caixas operantes idênticas (modelo Lafayette 80201) medindo 20 cm x 20 cm x 23 cm com teto, lado frontal e lado traseiro de acrílico, laterais de alumínio e piso de grade metálica. Cada caixa operante continha um bebedouro no centro da parede de alumínio esquerda e uma barra localizada à direita do bebedouro. As caixas operantes possuíam iluminação individual ininterrupta com uma lâmpada vermelha de 5 W sobre o teto de acrílico e continham uma lâmpada branca de 5 W, também sobre o teto de acrílico, cuja luz funcionou como o estímulo-característica L. Os estímulos-alvo, emitidos por caixas de som de 20 W individuais para cada caixa operante, foram um tom puro de 1.000 Hz e 30 dB (T), produzido por um gerador de áudio, e um ruído branco de 68 dB (R), previamente gravado em fita K7. Cada caixa operante, junto com suas lâmpadas e sua caixa de som, foi acusticamente isolada dentro de uma caixa de madeira medindo 55 cm x 55 cm x 55 cm. As quatro caixas de madeira foram, por sua vez, mantidas dentro de cubículos experimentais também isolados acusticamente. Controlou-se todo o experimento em uma sala adjacente através de uma interface conectada a um microcomputador, acionada por programas computacionais preparados com Microsoft QuickBASIC 4.0 (ambiente MS-DOS). As respostas de pressão à barra de cada rato foram registradas automaticamente pelos mesmos programas computacionais.
Lesão Seletiva
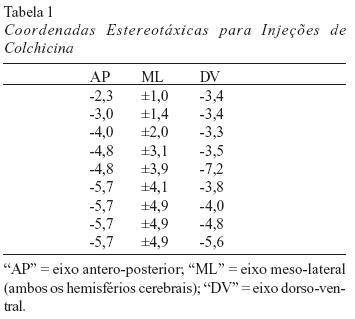
Os ratos foram submetidos à cirurgia antes da fase experimental. Os animais foram anestesiados com injeção intraperitoneal de uma solução anestésica que contém pentobarbital sódico (Hypnol 3%) diluído a 32,7%. A diluição do pentobarbital sódico foi feita no próprio laboratório e cada rato recebeu 0,3 mL da solução anestésica para cada 100 g de massa corpórea. Após a anestesia, a cabeça dos animais foi fixada em aparelho estereotáxico, o couro cabeludo sofreu uma incisão longitudinal com bisturi e a face dorsal do crânio foi removida por desgaste utilizando-se uma broca. Desta forma, o encéfalo ficou exposto para a aplicação de microinjeções da neurotoxina colchicina dissolvida em tampão fosfato-salina na concentração de 7 mg/mL e pH 7,4. As microinjeções obedeceram às coordenadas de Paxinos e Watson (1986) e foram aplicadas com microsseringas Hamilton de 5 mL possuindo micropipetas de vidro adaptadas à extremidade da agulha de metal.
O bregma do crânio foi tomado como ponto zero para o eixo ântero-posterior. Os pontos zero do eixo meso-lateral fizeram parte da linha mediana, evidenciada pela sutura sagital do crânio. E os pontos zero do eixo dorso-ventral foram orientados com base na superfície da dura-máter. Os pontos de injeção foram os mesmos usados por Xavier et al. (1999) e Costa et al. (2005), e estão expostos na Tabela 1. Em cada ponto estereotáxico foi injetado 0,06 mL da neurotoxina. Ao final de cada cirurgia o corte do couro cabeludo foi suturado. Os ratos controle-operados foram submetidos aos mesmos procedimentos, exceto pela injeção da neurotoxina colchicina.
Treino
Depois de uma recuperação pós-cirúrgica de duas semanas, todos os ratos foram modelados para localizar o bebedouro e pressionar a barra. Em seguida, fez-se uma sessão de CRF (continuous reinforcement) com 100 reforços para cada um dos animais. Após o CRF, fez-se um pré-treino de discriminação simples e, em seguida, um treino de discriminação condicional.
No pré-treino de discriminação simples, todos os ratos foram submetidos a 12 sessões diárias de, em média, 20 minutos. Cada sessão conteve cinco apresentações de tom puro de 5 s e cinco de ruído branco de 15 s, todas em ordem aleatória. As pressões à barra só foram reforçadas durante a apresentação destes estímulos sonoros, que foram separados por intervalos entre práticas de 2 min, em média. O tom puro e o ruído branco foram contrabalanceados para o grupo experimental e o controle. Assim, para metade do grupo experimental, as práticas de pré-treino foram T5+ (tom puro de 5 s sinalizando oportunidade de reforço) e R15+ (ruído branco de 15 s sinalizando oportunidade de reforço); para a outra metade do grupo experimental, as práticas de pré-treino foram T15+ (tom puro de 15 s sinalizando oportunidade de reforço) e R5+ (ruído branco de 5 s sinalizando oportunidade de reforço). Utilizou-se a mesma divisão para o grupo controle. A descrição do pré-treino, inclusive as práticas de contrabalanceamento, está na Tabela 2.

No treino de discriminação condicional, todos os ratos foram submetidos a 60 sessões diárias de, em média, 60 minutos. Cada sessão conteve quatro tipos de práticas: duas de discriminação condicional positiva e duas de discriminação condicional negativa. As duas práticas de discriminação condicional positiva foram L15T5+, que representa luz de 15 s seguida de intervalo vazio de 5s precedendo tom puro de 5 s (sendo que pressões à barra durante o tom puro de 5 s eram reforçadas) e T5-, que representa tom puro de 5 s apresentado sozinho e sem oportunidade de reforço. As duas práticas de discriminação condicional negativa foram L5R15-, que representa luz de 5 s seguida de intervalo vazio de 5s precedendo ruído branco de 15 s (sendo que pressões à barra durante o ruído branco de 15s não eram reforçadas) e R15+, que representa ruído branco de 15 s apresentado sozinho e com oportunidade de reforço. Cada um dos quatro tipos de práticas foi apresentado três vezes por sessão em ordem aleatória, e os intervalos entre práticas duraram, em média, 5 min. A fim de evitar o controle do desempenho do animal pelo tempo total de cada prática composta, estas tiveram uma duração total de 25 s. Para tanto, o estímulo-alvo precedido por L15 teve a duração de 5 s e o estímulo-alvo precedido por L5 teve a duração de 15 s. Os estímulos-alvo tom puro e ruído branco foram contrabalanceados para o grupo experimental e o controle. Assim, para metade do grupo experimental, os estímulos-alvo de discriminação condicional positiva e negativa foram, respectivamente, T e R; para a outra metade do grupo experimental, os estímulos-alvo de discriminação condicional positiva e negativa foram, respectivamente, R e T. Utilizou-se a mesma divisão para o grupo controle. Todas as práticas, inclusive as de contrabalanceamento, estão descritas na Tabela 2.
O critério de aquisição para o término do treino foi a observação da estabilização do desempenho em discriminação condicional positiva e negativa ao longo de três blocos de cinco sessões sucessivos.
Teste
Após o treino foi iniciada a fase de teste. A primeira sessão diária desta fase foi uma sessão de replicação do treino; a segunda sessão foi de teste de discriminação condicional positiva; a terceira sessão foi outra replicação do treino e a quarta foi uma sessão de teste de discriminação condicional negativa. Este esquema de alternância foi, então, repetido, o que resultou na realização de quatro sessões diárias de teste: duas de teste de discriminação condicional positiva e duas de teste de discriminação condicional negativa. As práticas do teste diferiram das de treino por apresentarem diferentes durações do estímulo-característica L: 15 s, 13 s, 11 s, 9 s, 7 s e 5 s. Além disto, as práticas do teste não incluíram oportunidades de reforçamento, ou seja, foram feitas em extinção. Deste modo, nas duas sessões de teste de discriminação condicional positiva foram apresentadas as práticas: L15T5-, L13T5-, L11T5-, L9T5-, L7T5-, L5T5- e T5-. Nas duas sessões de teste de discriminação condicional negativa, as práticas foram L15R15-, L13R15-, L11R15-, L9R15-, L7R15-, L5R15- e R15-. Para os ratos de contrabalanceamento, o teste de discriminação condicional positiva teve as práticas L15R5-, L13R5-, L11R5-, L9R5-, L7R5-, L5R5- e T5-. Ainda para o grupo de contrabalanceamento, o teste de discriminação condicional negativa teve as práticas L15T15-, L13T15-, L11T15-, L9T15-, L7T15-, L5T15- e T15. Cada prática de teste foi apresentada somente uma vez por sessão.
Histologia
Ao término do procedimento experimental, os ratos foram perfundidos com solução tampão de sulfeto de sódio. Os encéfalos foram retirados dos crânios, fixados em carnoy e incluídos em parafina. Os cortes histológicos, de 20 mm, foram feitos ao micrótomo. A cada 10 fatias, uma era colocada em lâmina histológica e as demais eram descartadas. Utilizou-se a coloração com violeta de cresil para visualização de corpúsculos de Nissl e, portanto, dos corpos celulares das células granulares pertencentes ao giro denteado do hipocampo.
Depois de preparadas, as lâminas foram observadas por meio de um microscópio óptico Olympus acoplado a uma câmara do mesmo fabricante. As imagens capturadas pela câmara foram diretamente transmitidas a um microcomputador, por meio do qual se obtiveram fotomicrografias digitais dos cortes histológicos de todos os encéfalos extraídos, evidenciando-se o hipocampo em cada um dos hemisférios cerebrais. De posse das fotomicrografias, foi medida a área do giro denteado de cada rato, utilizando-se o programa computacional Image Tool, desenvolvido na UTHSCSA (The University of Texas Health Science Center in San Antonio).
Análise estatística
Foi empregado o teste t para amostras independentes a fim de comparar as áreas médias dos giros denteados de ratos lesados e controle-operados com base nas fotomicrografias do hipocampo. Deste modo, foi avaliada a efetividade das lesões.
Posteriormente, com base no registro de pressões à barra do pré-treino de discriminação simples, foi calculada, para cada sessão, a porcentagem de respostas emitidas durante os estímulos sonoros T5 e R15 (R5 e T15 para o grupo de contrabalanceamento) em relação ao total de respostas emitidas. Com as porcentagens de cada sessão, foram construídas duas curvas de aquisição representativas do desempenho dos grupos experimental e controle ao longo do pré-treino. As duas curvas foram analisadas com ANOVA (análise de variância) para avaliar se houve: (1) efeito de grupos, ou seja, se houve diferença de desempenho entre os grupos desconsiderando-se as formas das curvas de aquisição; (2) efeito de sessões, ou seja, se houve aquisição ao longo das sessões; e (3) efeito de interação grupos/sessões, ou seja, se houve diferença significativa entre as curvas de aquisição referentes aos dois grupos de ratos ao longo do pré-treino.
Por sua vez, os dados do treino forneceram porcentagens de estímulos durante os quais foi emitida pelo menos uma resposta em relação ao total de vezes em que os estímulos foram apresentados, incluindo as apresentações em que não foi emitida nenhuma resposta. As curvas resultantes desta análise também foram submetidas à ANOVA para que se avaliasse efeito de grupos, efeito de sessões e efeito de interação grupos/sessões. Para a análise dos dados do treino, as 60 sessões foram divididas em blocos de cinco sessões.
A ANOVA também foi utilizada para a análise dos dados obtidos com o teste de discriminação condicional. Com base em números médios de respostas durante os estímulos-alvo, apresentados nas duas sessões de teste de discriminação condicional positiva e nas duas sessões de teste de discriminação condicional negativa, avaliou-se o desempenho dos ratos após a apresentação dos estímulos-característica de diferentes durações. Tal comparação foi feita entre as diferentes durações do estímulo-característica através do método post hoc de Newman-Keuls. O nível a adotado durante toda a análise estatística foi p£0,05.
Resultados
Pré-treino
Houve aquisição da discriminação simples, já que a porcentagem de respostas emitidas durante os estímulos aumentou ao longo das 12 sessões, tanto para o grupo experimental quanto para o controle. A ANOVA indicou efeito de sessões, confirmando a aquisição da discriminação simples nos dois grupos (F11,154=21,9552; p<0,05). Entretanto, não foi encontrado efeito de grupos nem de interação de grupos e sessões.
Também houve diminuição do número de respostas ao longo do pré-treino, independentemente da presença ou ausência dos estímulos sonoros, o que também indica aquisição da discriminação simples para ambos os grupos de ratos (F11,154=14,0914; p<0,05). Não houve efeito de interação de grupos e sessões, mas foi detectado efeito de grupos (F1,14=5,9366; p<0,05), o que indica que os ratos lesados emitiram respostas a taxas regularmente mais altas do que os controle-operados do início ao fim do pré-treino.
Treino
Ambos os grupos de ratos iniciaram o treino com altas taxas de respostas durante qualquer estímulo-alvo, independentemente se precedidos ou não por estímulos-característica. Isto mostra que, nas primeiras sessões de treino, os ratos ainda se comportavam conforme as contingências estabelecidas no pré-treino de discriminação simples (Figura 2).
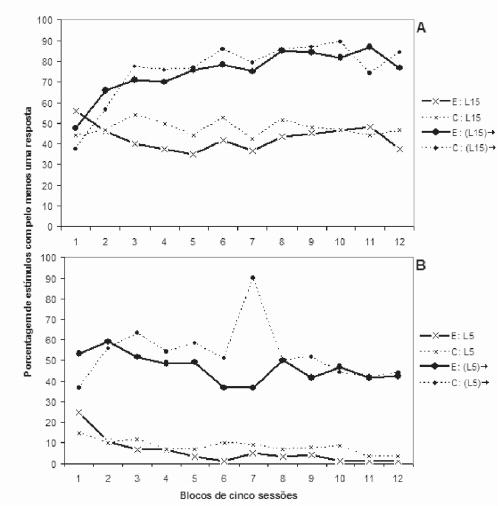
No treino de discriminação condicional positiva, para os dois grupos de ratos não houve efeito de grupos (F1,14=0,2459; p>0,05), nem de sessões (F11,154=0,5390; p>0,05) e nem de interação grupos/sessões (F11,154=0,7341; p>0,05) quanto à porcentagem de apresentações de L15 durante as quais foi emitida pelo menos uma resposta (Figura 1A). Quanto ao intervalo vazio de cinco segundos após L15, também não houve efeito de grupos (F1,14=0,0302; p>0,05) nem de interação grupos/sessões (F11,154=0,9494; p>0,05). No entanto, ao longo das sessões houve aumento significativo da porcentagem de intervalos vazios após L15 com pelo menos uma resposta (F11,154=10,6748; p<0,05), o que indica que L15 adquiriu propriedade excitatória para ambos os grupos de ratos (Figura 1A).
No treino de discriminação condicional negativa, para os dois grupos de ratos não houve efeito de grupos (F1,14=0,7545; p>0,05) nem de interação grupos/sessões (F11,154=1,7767; p>0,05) quanto à porcentagem de apresentações de L5 durante as quais foi emitida pelo menos uma resposta (Figura 1B). No entanto, ao longo das sessões houve diminuição significativa da porcentagem de L5 com pelo menos uma resposta (F11,154=7,0324; p<0,05), o que indica que L5 adquiriu propri edade inibitória para ambos os grupos de ratos (Figura 1B). Quanto ao intervalo vazio de cinco segundos após L5, não houve efeito de grupos (F1,14=1,0863; p>0,05). Porém, ao longo das sessões houve diminuição significativa da porcentagem de intervalos vazios após L5 com pelo menos uma resposta, confirmada pelo efeito de sessões (F11,154=2,5092; p<0,05) e pelo efeito de interação grupos/sessões (F11,154=3,7918; p<0,05). Isto indica, novamente, que L5 adquiriu propriedade inibitória para ambos os grupos de ratos (Figura 1B).
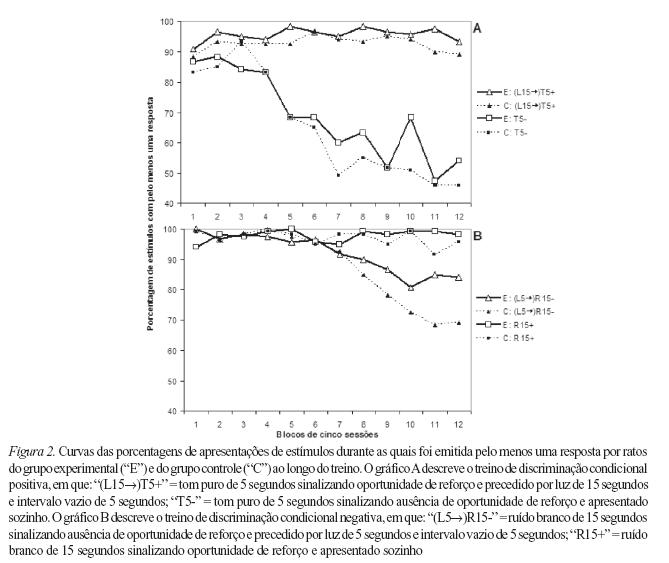
No treino de discriminação condicional positiva, para os dois grupos de ratos não houve efeito de grupos (F1,14=1,2557; p>0,05), nem de sessões (F11,154=1,2642; p>0,05) e nem de interação grupos/sessões (F11,154=0,3699; p>0,05) quanto à porcentagem de apresentações de T5+ precedido por L15 durante as quais foi emitida pelo menos uma resposta (Figura 2A). Quanto ao T5- apresentado sozinho, também não houve efeito de grupos (F1,14=0,1652; p>0,05) nem de interação grupos/sessões (F11,154=0,6111; p>0,05). No entanto, ao longo das sessões houve diminuição significativa da porcentagem de T5- apresentado sozi nho com pelo menos uma resposta para ambos os grupos de ratos (F11,154=13,1438; p<0,05) (Figura 2A).
Tanto o aumento do responder durante o intervalo vazio após L15 quanto a comparação entre as curvas correspondentes a T5+ precedido por L15 e T5- apresentado sozinho indicam que L15 adquiriu propriedade excitatória para os dois grupos de ratos. Portanto, os ratos do grupo experimental e do grupo controle adquiriram, sem diferenças significativas de desempenho entre os dois grupos, a discriminação condicional positiva.
No treino de discriminação condicional negativa, para os dois grupos de ratos não houve efeito de grupos (F1,14=2,3936; p>0,05) nem de interação grupos/sessões (F11,154=1,3395; p>0,05) quanto à porcentagem de apresentações de R15- precedido por L5 durante as quais foi emitida pelo menos uma resposta (Figura 2B). Porém, ao longo das sessões houve diminuição significativa da porcentagem de R15- precedido por L5 com pelo menos uma resposta para ambos os grupos de ratos (F11,154=11,0225; p<0,05). Quanto ao R15+ apresentado sozinho, para os dois grupos de ratos não houve efeito de grupos (F1,14=0,4368; p>0,05), nem de sessões (F11,154=1,2758; p>0,05) e nem de interação grupos/sessões (F11,154=1,6768; p>0,05).
Tanto a diminuição do responder durante L5 e o intervalo vazio após L5 quanto a comparação entre as curvas correspondentes a R15- precedido por L5 e R15+ apresentado sozinho indicam que L5 adquiriu propriedade inibitória. Portanto, os ratos do grupo experimental e do grupo controle adquiriram, sem diferenças significativas de desempenho entre os dois grupos, a discriminação condicional negativa.
Teste
Os resultados do teste de discriminação condicional positiva não indicaram diferenças significativas entre o grupo experimental e o controle, já que não houve efeito de grupos ao longo de todas as durações de L (F1,14 = 2,7467; p > 0,05) (Figura 3A). Para os dois grupos de ratos, o número de respostas durante os estímulos-alvo precedidos por L15 e L13 foi significativamente maior do que o número de respostas durante estímulos-alvo precedidos por L9, L7 e L5 (Newman-Keuls; p < 0,05). O número de respostas após L11 só é significativamente maior em relação ao número de respostas após L5. E o número de respostas durante o estímulo-alvo apresentado sozinho (T5-) é significativamente menor em relação a todos os demais estímulos-alvo, não importando a duração da luz precedente.
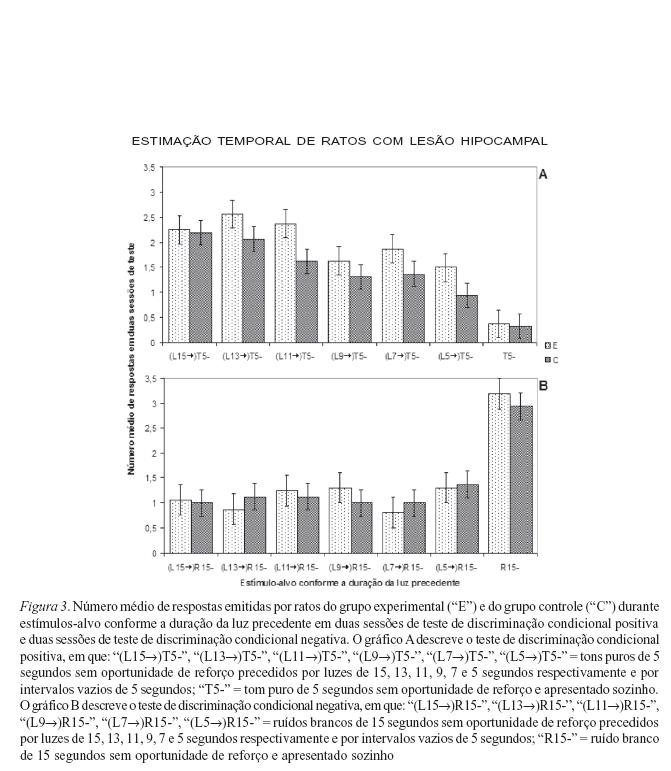
Quanto ao teste de discriminação condicional negativa (Figura 3B), também não houve diferença entre os desempenhos dos grupos experimental e controle ao longo de todas as durações de L (F1,14 = 0,0253; p > 0,05). Para ambos os grupos a taxa de respostas foi significativamente mais alta durante o estímulo-alvo sozinho em relação aos demais estímulos-alvo, não importando a duração da luz precedente (Newman-Keuls; p= 0,05). Dentre os números de respostas durante estímulos-alvo precedidos por quaisquer durações de L, o teste de Newman-Keuls não indicou nenhuma diferença significativa.
Histologia
A área do giro denteado de animais lesados mostrou-se significativamente diminuída em comparação com animais controle-operados: t = -5,8463; p < 0,05 (hemisfério direito) e t = -3,3990; p < 0,05 (hemisfério esquerdo) (Figura 4).
Discussão
A análise do treino de discriminação condicional permite concluir que um estímulo-característica de mesma modalidade pode adquirir propriedade excitatória ou inibitória, dependendo da duração com que é apresentado em treino de discriminação condicional positiva e negativa, respectivamente. Estes resultados foram obtidos tanto para animais do grupo experimental quanto para animais do grupo controle. Assim, as durações, como representações amodais, podem ser discriminadas por ratos, o que confirma Meck e Church (1982).
O teste de discriminação condicional positiva realizado por Martins (2001) apontou para a possibilidade de os ratos estarem sob controle de uma bi-seção temporal, pois naquele traba lho o número de respostas após L15, L13 e L11 foi significativamente maior do que o número de respostas após L9, L7 e L5. No presente trabalho, para os dois grupos de ratos os números de respostas após L9, L7 e L5 só foram significativamente menores do que os números de respostas após L15 e L13, enquanto os dados representativos de L11 só diferiram significativamente dos de L5. Apesar destas distinções entre Martins (2001) e o presente trabalho quanto ao teste de discriminação condicional positiva, de modo geral os resultados do teste mostram que ratos, após aprenderem a discriminar entre propriedades condicionais de luz de longa duração e luz de curta duração, respondem conforme duas categorias de duração (longa e curta) quando submetidos a um teste que apresenta durações intermediárias. Este desempenho pode estar sob controle de um processo de bi-seção temporal (Church & Deluty, 1977).
Os ratos lesados no giro denteado responderam a taxas regularmente mais altas durante todo o pré-treino. Taxas maiores de respostas de ratos lesados no hipocampo em comparação com o grupo controle também foram observadas em outros trabalhos envolvendo discriminação, tanto com procedimentos operantes quanto com procedimentos pavlovianos (Alvarado & Rudy, 1995; Davidson, McKernan, & Jarrard, 1993; Holland, Lamourex, Han, & Gallagher 1999; Moreira & Bueno, 2003; Sutherland & Rudy, 1989). A lesão seletiva do giro denteado, especificamente, já foi descrita como envolvida no aumento da atividade motora (Barone, Bonner, Tandon, McGinty & Tilson, 1992).
Apesar de os ratos do grupo experimental terem respondido a taxas regularmente mais altas do que os ratos do grupo controle durante todo o pré-treino, os resultados do treino não apontam para a participação das células granulares do giro denteado hipocampal na estimação temporal, o que contrasta com os resultados de Costa et al. (2005) e Bueno et al. (2006) obtidos com DRL. Ao discutir o desempenho de ratos em DRL, O´Keefe e Nadel (1978) defendem que ratos baseiam a estimação temporal em seqüências comportamentais estereotipadas, o que é coerente com a teoria do relógio comportamental de Killeen e Fetterman (1988). A perda seletiva de células granulares do giro denteado provoca aumento da atividade motora de ratos (Barone et al., 1992). Assim, o aumento da atividade motora decorrente da lesão seletiva do giro denteado pode afetar as seqüências comportamentais nas quais os ratos se engajam durante a produção de intervalos, o que explicaria o prejuízo de desempenho no DRL.
Os resultados do pré-treino do presente estudo confirmam que a lesão seletiva do giro denteado aumenta a atividade motora. No entanto, os resultados do treino não mostram prejuízo de desempenho dos ratos lesados. Portanto, a ativação das células do giro denteado parece não exercer papel fundamental na estimação temporal, mas sua lesão prejudica o relógio comportamental através do aumento da atividade motora, o que explica o prejuízo de desempenho em DRL e a falta desse prejuízo em discriminação condicional com manipulação da duração de estímulos. Isso mostra que enquanto dados obtidos com DRL são mais coerentes com a teoria do relógio comportamental (Killeen & Fetterman, 1988), os dados obtidos com discriminação condicional temporal são mais coerentes com modelos cognitivos de processamento temporal, como o modelo do relógio interno defendido na teoria da expectativa escalar (Gibbon, 1977; Gibbon & Church, 1990).
Ainda que os animais lesados do presente estudo tenham adquirido normalmente a discriminação condicional temporal, é possível que seqüências comportamentais este reotipadas realizadas durante as práticas tenham sido afetadas pela lesão do giro denteado. No entanto, como este trabalho registrou somente o comportamento operante de pressão à barra, os dados não permitem acessar a eventual diferenciação das seqüências comportamentais de ratos do grupo experimental e do grupo controle. Bueno et al. (2007) realizaram um treino idêntico ao utilizado pelo presente estudo, porém com registro de diferentes categorias de comportamentos sob procedimento pavloviano de discriminação condicional. Assim, a retomada da abordagem pavloviana de Bueno et al. (2007) possibilitaria a análise das seqüências comportamentais de ratos lesados no giro denteado durante a discriminação condicional temporal.
Uma hipótese alternativa é a de que a lesão do giro denteado do hipocampo afeta a estimação temporal tanto em procedimentos de DRL quanto em procedimentos de discriminação condicional temporal. Neste caso, os ratos do presente estudo teriam encurtado de maneira equivalente todas as durações de L apresentadas durante o treino e o teste. Ou seja, se a duração de 15 s foi subestimada em determinada proporção, assim como aconteceu nos estudo de Costa et al. (2005) e Bueno et al. (2006), todas as durações intermediárias apresentadas no teste podem ter sido subestimadas na mesma proporção. Dessa forma, a diferença relativa entre as representações das durações intermediárias de L apresentadas no teste teria se mantido inalterada para os ratos lesados, devido às diminuições equivalentes das durações representadas. Procedimentos com manipulações específicas do efeito de bi-seção temporal poderiam esclarecer melhor o efeito de lesão do giro denteado do hipocampo sobre a estimação de durações de estímulos.
Portanto, para melhor compreensão sobre os processos neurofisiológicos subjacentes à representação de durações, são necessários novos estudos que evidenciem a distinção entre dois processos: a estimação temporal não sinalizada e a discriminação de durações de estímulos. Tais estudos poderão esclarecer o papel específico que o giro denteado hipocampal pode exercer no processamento do tempo subjetivo.
Referências
Alvarado, M. C., & Rudy, J. W. (1995). A comparison of kainic acid plus colchicine and ibotenic acid-induced hippocampal formation damage on four configural tasks in rats. Behavioral Neuroscience,109(6), 1052-1062. [ Links ]
Barone, S., Bonner, M., Tandon, P., McGinty, J. F., & Tilson, H. A. (1992). The neurobiological effects of colchicine: Modulation by nerve growth factor. Brain Research Bulletin, 28 (2), 265-274. [ Links ]
Bueno, J. L. O. (1997). O imaginário animal. Psicologia Universidade de São Paulo, 8(2), 165-180. [ Links ]
Bueno, J. L. O., Costa, V. C. I., Xavier, G. F., & Dima, L. L. (2006). Aquisição de uma tarefa temporal (DRL) por ratos submetidos a lesão seletiva do giro denteado. Psicologia: Reflexão & Crítica, 19(1), 159-165. [ Links ]
Bueno, J. L. O., Wagatsuma, A., & Martins, M. R. (2007). Estimação de tempo em ratos submetidos a um procedimento de discriminação condicional. Psicologia: Reflexão & Crítica, 20(2), 238-245. [ Links ]
Church, R. M., & Deluty, M. Z. (1977). Bisection of temporal intervals. Journal of ExperimentalPsychology: Animal Behavior Processes, 3(3), 216-228. [ Links ]
Costa, V. C. I., Bueno, J. L. O., & Xavier, G. F. (2005). Dentate gyrus-selective colchicine lesion and performance in temporal and spatial tasks. Behavioural Brain Research, 160(2), 286-303. [ Links ]
Davidson, T. L., McKernan, M. G., & Jarrard, L. E. (1993). Hippocampal lesions do not impair negative patterning: A challenge to configural association theory. Behavioral Neuroscience, 107(2), 227-234. [ Links ]
Gallistel, C. R. (1990). The organization of learning. Cambridge: MIT Press. [ Links ]
Gibbon, J. (1977). Scalar expectancy theory and Weber´s law in animal timing. Psychological Review, 84(3), 279-325. [ Links ]
Gibbon, J., & Church, R. M. (1990). Representation of time. Cognition, 37(1-2), 23-54. [ Links ]
Gibson, J. J. (1960). The concept of stimulus in psychology. American Psychologist, 15(11), 694-703. [ Links ]
Goldschmidt, R. B., & Steward, O. (1980). Preferential neurotoxicity of colchicine for granule cells of the dentate gyrus of the adult rat. Proceedings of The National Academy of Sciences The United States of America: Biological Sciences, 77(5), 3047-3051. [ Links ]
Goldschmidt, R. B., & Steward, O. (1982). Neurotoxic effects of colchicine: Differential susceptibility of CNS neuronal populations. Neuroscience, 7(3), 695-714. [ Links ]
Hanson, M. & Estrom, A. (1978). Mitosis inhibitors and axonal transport. International Reviews of Cytology, 7(Suppl.), 373-402. [ Links ]
Holland, P. C. (1984). Differential effects of reinforcement of an inhibitory feature after serial and simultaneous feature negative discrimination training. Journal of Experimental Psychology: Animal Behavior Processes, 10(4), 461-475. [ Links ]
Holland, P. C. (1985). The nature of conditioned inhibition in serial and simultaneous feature-negative discriminations. In R. R. Miller & N. E. Spear (Eds.), Information processing in animals: Conditioned inhibition (pp. 267-297). Hillsdale: Erlbaun. [ Links ]
Holland, P. C. (1998). Temporal control in Pavlovian occasion setting. Behavioural Processes, 44(2), 225-236. [ Links ]
Holland, P. C., & Lamarre, J. (1984). Transfer of inhibition after serial and simultaneous feature negative discrimination training. Learning and Motivation, 15(3), 219-243. [ Links ]
Holland, P. C. Lamoreux, J. A., Han, J-S., & Gallagher, M. (1999). Hippocampal lesions interfere with Pavlovian negative occasion setting. Hippocampus, 9(2), 143-157 [ Links ]
Jenkins, H. M., & Sainsbury, R. S. (1969). The development of stimulus control through differential reinforcement. In N. J. Mackintosh & W. K. Honig (Eds.), Fundamental issues in associative learning. Halifax: Dalhoise University Press. [ Links ]
Killeen, P. R., & Fetterman, J. G. (1988). A behavioral theory of timing. Psychological Review, 95(2), 274-295. [ Links ]
Lawrence, D. H. (1963). The nature of a stimulus: Some relationships between learning and perception. In S. Koch (Ed.), Psychology: A study of a science (pp. 179-212). New York: McGraw-Hill. [ Links ]
Martins, M. R. (2001). Estimação de tempo em ratos submetidos a uma tarefa de discriminação condicional em um procedimento operante. Dissertação de bacharelado não-publicada, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo. Ribeirão Preto, Brasil. [ Links ]
Meck, W. H., & Church, R. M. (1982). Abstraction of temporal attributes. Journal of Experimental Psychology: Animal Behavior Processes, 8(3), 226-243. [ Links ]
Moreira, R. C. M., & Bueno, J. L. O. (2003). Conditional discrimination learning and negative patterning in rats with neonatal hippocampal lesion induced by ionizing radiation. Behavioural Brain Research,138(1), 29-44. [ Links ]
O´Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford: Oxford University Press. [ Links ]
Paxinos, G., & Watson, C. (1986). The rat brain in stereotaxic coordinates. London: Academic Press. [ Links ]
Pearce, J. M., & Hall, G. (1980). A model for pavlovian learning: variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychological Review, 87(6), 532-552. [ Links ]
Rescorla, R. A., & Holland, P. C. (1977). Associations in pavlovian conditioned inhibition. Learning and Motivation, 8(4), 429-447. [ Links ]
Ross, R. T., & Holland, P. C. (1981). Conditioning of simultaneous and serial feature-positive discriminations. Animal Learning & Behavior, 9(3), 293-303. [ Links ]
Sutherland, R. J., & Rudy, J. W. (1989). Configural association theory: The role of the hippocampal formation in learning, memory and amnesia. Psychobiology, 17(2), 129-144. [ Links ]
Vauclair, J. (1996). Animal cognition: An introduction to modern comparative psychology. Cambridge: Harvard University Press. [ Links ]
Wagner, A. R., & Rescorla, R. A. (1972). Inhibition in pavlovian conditioning: Applications of a theory. In M. S. Halliday & R. A. Boakes (Eds.), Inhibition and learning (pp. 301-336). London: Academic Press. [ Links ]
Xavier, G. F., Oliveira-Filho, F. J. B., & Santos, A. M. G. (1999). Dentate gyrus-selective colchicine lesion and disruption of performance in spatial tasks: Difficulties in "place strategy" because of a lack of flexibility in the use of environmental cues? Hippocampus, 9(6), 668-681. [ Links ]
Received 16/11/2006
Accepted 09/05/2007
José Lino Oliveira Bueno. Graduado em Filosofia, obteve seu Doutorado em Psicologia Experimental no Instituto de Psicologia da Universidade de São Paulo (USP). É Professor Titular de Psicobiologia da USP, Campus de Ribeirão Preto, onde coordena o Laboratório de Processos Associativos, Controle Temporal e Memória e o Laboratório de Estética Experimental. Foi Fulbright Visiting Professor na Duke University (USA) e é membro do Editorial Board de "Philosophy and Behavior".
Lézio Bueno Jr. Graduado em Biologia pelo Departamento de Biologia da Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, é atualmente aluno do Programa de Mestrado em Psicobiologia da USP-Ribeirão Preto.
1 A pesquisa foi financiada com Bolsa de Pesquisador I do CNPq para J.L.O. Bueno e de Iniciação Científica do CNPq para L. Bueno Jr.
2 Endereço: Faculdade de Filosofia, Ciências e Letras, Departamento de Psicologia e Educação, Universidade de São Paulo. Avenida dos Bandeirantes 3900.CEP 14040-930 Ribeirão Preto, SP, Brasil. E-mail: jldobuen@ffclrp.usp.br
