










Serviços Personalizados
artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista Latinoamericana de Psicología
versão impressa ISSN 0120-0534
Rev. Latinoam. Psicol. v.38 n.1 Bogotá 2006
ARTÍCULOS
Análisis conceptual del aprendizaje observacional y la imitación
A conceptual analysis of observational learning and imitation
J. C. Pedro Arriaga-Ramírez*,**; María Guadalupe Ortega-Saavedra; Gustavo Meza Reyboso; Francisco Huichán Olivares; Edith Juárez Maldonado; Adriana Rodríguez Cuadros; Sara E. Cruz-Morales
Universidad Nacional Autónoma de México
RESUMEN
En este trabajo se analizaron los conceptos que se han presentado para explicar lo que ocurre en situaciones de aprendizaje social, aprendizaje observacional e imitación. Con el uso del modelo de desarrollo del comportamiento presentado por Hogan (1994b) para organizarlos se señaló que los conceptos analizados se refieren a diferentes fuentes de control del estímulo. Se revisaron, asimismo, los factores asociativos que determinan tanto el aprendizaje social como el cambio de valencia y la potenciación; se indicó, también, la relación del aprendizaje social con la disposición de la ocasión. Se sugiere que estos tres términos equivalen a un paradigma de discriminación condicional.
Palabras clave: Aprendizaje social, Aprendizaje observacional, Imitación, Factores asociativos, Discriminación condicional.
ABSTRACT
Concepts presented to explain social learning, observational learning and imitation were analyzed. Hogan’s (1994) model for behavioral development was used to organize them and to show that those concepts refer to different sources of stimulus control, rather than to different processes. Valence change and potentiation were named as associative factors that determine observational learning. The relation between observational learning and occasion setting was underlined. It is suggested that these three terms are variants of the conditional discrimination paradigm.
Keywords: Social learning, Observational learning, Imitation, Associative factors, Conditional Discrimination.
INTRODUCCIÓN
En los últimos 25 años se han presentado recopilaciones y análisis teóricos de conceptos respecto del aprendizaje social, el aprendizaje por observación y la imitación en organismos infrahumanos; diversos autores han analizado de manera crítica el aspecto social involucrado en el aprendizaje de nuevos comportamientos en los animales (Byrne, 1994; Coussi-Korbel & Fragaszy, 1995; Galef, 1988; Heyes, 1993, 1994, 1996; Heyes & Galef, 1996; Miklósi, 1999a; Moore, 1996; Whiten & Ham, 1992; Zentall, 1996), sin embargo, estos análisis sólo han producido una serie de conceptos que tratan de dar cuenta de algunos aspectos relacionados con el aprendizaje social, el aprendizaje observacional y la imitación y parecen sugerir que no hay relación entre ellos. En el ámbito latinoamericano se han publicado diferentes trabajos en los que se han analizado algunos factores que intervienen en el aprendizaje social y la imitación. Ejemplos de estos casos son los trabajos de Cabrera y Nieto (1993), Nieto y Cabrera (1994), Nieto, Cabrera, Guerra y Posadas-Andrews (1987) y Rodas (1974).
Debido a lo anterior, se planeó realizar un análisis de esos conceptos con el fin de proponer una manera de organizarlos y mostrar que, más que referirse a diferentes procesos, todos forman parte de uno global, y que cada uno se refiere a aspectos diferentes de una situación en la que se estudian tanto el aprendizaje social, como el aprendizaje observacional y la imitación. Los resultados de la revisión se presentarán en cinco secciones: a) descripción de los procedimientos para estudiar el aprendizaje social; b) conceptos utilizados en aprendizaje social; c) propuesta del uso de un modelo para analizar el aprendizaje social; d) factores que determinan el aprendizaje social; y e) conclusiones.
Es importante analizar de manera precisa los diferentes aspectos del aprendizaje social, del aprendizaje observacional y de la imitación porque, aunque los dos últimos son casos de aprendizaje social, no resultan sinónimos.
Estos conceptos se analizarán partiendo de lo más general a lo más específico. El aprendizaje observacional y la imitación son elementos del aprendizaje social, ya que en los tres casos se involucra la participación de un coespecífico. En el aprendizaje social un organismo puede influir de diversas maneras sobre otro u otros. Puede transmitir cierta información desde la olfativa o la auditiva hasta la visual, que puede involucrar, asimismo, la forma del comportamiento del coespecífico. El que “observa” puede “aprender” por medio del olfato, la audición o la visión; por ello, el aprendizaje social incluye tanto al aprendizaje observacional como a la imitación, así como a otros comportamientos que son guiados por señales olfativas, auditivas o visuales. Como se verá en el desarrollo de este trabajo, para la mayoría de los autores la imitación involucra aspectos de mayor complejidad que los del aprendizaje social y del aprendizaje observacional, pues requiere que un animal reproduzca de manera motriz un comportamiento que percibió de manera visual. También involucra la observación precisa de un comportamiento o respuestas del modelo o demostrador, a diferencia de algunos tipos de aprendizaje social y aprendizaje observacional que involucran la observación de la situación física o de los estímulos presentes en el episodio social en el que participó un coespecífico. De acuerdo con Heyes (1993), la diferencia podría resumirse en que en el aprendizaje social y el aprendizaje observacional los animales aprenden algo relativo al ambiente observando a otros; en cambio, en la imitación aprenden algo acerca del comportamiento observando a otros.
Procedimientos utilizados para el estudio del aprendizaje social
El aprendizaje social se produce en el medio natural en algunas especies animales. Al estudiarlo en situaciones controladas es necesario tomar en cuenta algunos aspectos del episodio de interacción social, como el hecho de que involucra de manera general la presentación de información a un observador (O) mediante un coespecífico demostrador (D). El D puede, por un lado, hacer contacto con los estímulos físicos presentes en el episodio de interacción y dejar un rastro olfatorio; por otro lado, puede presentar un comportamiento factible de ser conocido o novedoso para el O. Así, en un episodio de estos tipos de aprendizaje, el D puede actuar sobre el ambiente físico o presentar una respuesta ya existente en el conjunto de respuestas del O, aunque también puede presentar una respuesta novedosa inexistente en el repertorio del O. En cuanto al periodo temporal en que ocurren los comportamientos del D y del O, la presentación de comportamientos por medio del O puede ocurrir casi simultáneamente con la del D o incluso puede ocurrir tiempo después. Así, la respuesta del O se puede presentar, en un momento dado, en forma simultánea con la del D en un primer tiempo (t1), o tiempo después, esto es, en un segundo tiempo (t2).
Desde un punto de vista del procedimiento, cuando se estructura una situación para el estudio del aprendizaje social se presentan dos fases: la de demostración y la de prueba. En algunas situaciones, durante la fase de demostración están presentes tanto el D como el O e interactúan en el t1 y sólo se analiza este periodo en el que el comportamiento del D provee de información al O. En otro tipo de situaciones, en una fase posterior, durante la fase de prueba, en el t2, se evalúa el aprendizaje adquirido por el O en t1. Este segundo tipo de procedimiento involucra la evaluación del aprendizaje de un aspecto del comportamiento del D y permite excluir &como generadores del mismo comportamiento& al aprendizaje acerca de algún aspecto del ambiente, puesto que ni el D ni los residuos olfatorios se encuentran ya presentes.
Conceptos utilizados en aprendizaje social
En esta sección se analizan algunos de los conceptos utilizados en una situación de aprendizaje social. Ninguna de las aproximaciones revisadas, como se mencionó antes, relaciona todos los aspectos de la situación. Se analizarán con base en dos aspectos: primero, con relación a los estímulos de la situación y, segundo, de las respuestas.
Referente a los aspectos de los estímulos están, en primer término, los propuestos por Galef (1988), quien analizó algunos conceptos usados para explicar el aprendizaje por observación en animales y mencionó los siguientes: señalamiento local, definido por Thorpe (1956, 1963) como: “imitación aparente que resulta de dirigir la atención del animal a un objeto particular o una parte particular del ambiente” (p. 134). Este concepto es el que se ha empleado más frecuentemente para explicar el aprendizaje observacional o la “aparente imitación” en animales. No obstante, Galef advirtió sobre la posibilidad de error al considerar que dicho concepto explica también a la imitación mediante una referencia a un escrito de Davis (1973), quien menciona: “... el uso del término [imitación] no debe ocultarnos que permanecemos ignorantes de muchos de los rasgos importantes de las interacciones sociales etiquetadas así: las condiciones necesarias y suficientes para que un organismo dirija la atención de otro a una porción del ambiente, y si, de hecho, cambios en el foco de “atención” verdaderamente tienen algo que ver con tales fenómenos [el aprendizaje social y la imitación]” (p. 15).
Galef (1988) indicó que el término resaltamiento del estímulo, utilizado por Spence (1937), es más apropiado, pues no hace referencia a aspectos no observables como la “atención” del O; además, tiene mayor alcance al incluir una clase de objetos que comparten características de estímulo con un objeto que el D manipula y con el que hace contacto o marca. Estos dos conceptos &señalamiento local y resaltamiento del estímulo&, hacen referencia a los aspectos estímulo de la situación de aprendizaje social.
En segundo término, mencionaremos los conceptos que hacen referencia al componente de las respuestas producidas en una situación social, en la cual se supone que la presencia del D &durante la fase de demostración en el t1-, facilita la emisión de respuestas que ya forman parte del repertorio del sujeto O.
Uno de estos conceptos, analizado por Galef (1988) es el de la facilitación social, introducido por Zajonc (1965, 1969) quien indica que la simple presencia de otros energiza todas las respuestas educidas por la situación estímulo confrontada por el individuo O en un momento dado, y que las respuestas más dominantes (aquellas que tienen una mayor probabilidad de emisión) son las que más se facilitan por la presencia de otros. No obstante, Galef mencionó que la evidencia que justifique el uso del concepto de facilitación social es escasa.
Otro término mencionado por Galef (1988) que se ha utilizado para explicar el aprendizaje observacional, es el de comportamiento contagioso. Thorpe lo define como “la ejecución de un patrón más o menos instintivo de comportamiento por uno [demostrador], tenderá a actuar como un liberador del mismo comportamiento en otros [observadores] y así iniciar la misma línea de acción en el mismo grupo” (1963, p. 133). Galef agrega: “Obviamente, el comportamiento contagioso, aunque produce resaltamiento [mediado] social [mente], es inadecuado para producir aprendizaje o transmisión social” (p. 18). Sin embargo, otros autores mencionan que aunque los observadores no estén controlados por la forma del comportamiento de los demostradores, el comportamiento contagioso puede contribuir para aumentar la probabilidad de algunas respuestas que podrían influir en la selección de alimentos (Suboski & Bartashunas, 1984). En este punto se puede señalar que una diferencia entre el resaltamiento social, el aprendizaje de tipo social y la transmisión social de información radica en que el aprendizaje y la transmisión social tienen mayor permanencia mientras que el resaltamiento social resulta efímero.
El nombre condicionamiento observacional, se ha utilizado con el fin de atribuir una explicación en términos de condicionamiento pavloviano al desarrollo de evitación a las víboras en monos rhesus (Cook, Mineka, Wolkenstein & Laitsch, 1985). Algunos autores también consideran que en ciertas situaciones el aprendizaje por observación puede considerarse como una forma de condicionamiento clásico (Heyes, 1994; Heyes & Dawson, 1990; Whiten & Ham, 1992). En el caso del desarrollo de evitación a las víboras, un O ingenuo asocia a la serpiente &estímulo condicionado&, con la reacción de miedo de los demostradores &estímulo incondicionado& y dichos autores suponen que esto puede ser equivalente a un ensayo de condicionamiento pavloviano, es decir, la víbora producirá en el O la misma respuesta condicionada que en el D.
Comportamiento dependiente de la igualación es un término utilizado tanto por Miller y Dollard (1941) como por Skinner (1953). Estos autores lo utilizan al interpretar el aprendizaje observacional como una respuesta reforzada ante estímulos discriminativos provistos por las acciones del D. Whiten y Ham (1992) subdividieron los procesos sociales en aprendizaje social e influencia social. Ellos explican en forma separada los dos tipos. En el aprendizaje social, señalan, el sujeto O “... aprende algún aspecto de la semejanza comportamental de A [el demostrador], mientras que en la influencia social B [el observador] no aprende la semejanza [comportamental] de A...” (p. 248). Whiten y Ham incluyen al aprendizaje dependiente de igualación en la subdivisión influencia social. En este caso, los observadores se comportan en una situación semejante a la del modelo y responden a los estímulos discriminativos; como resultado de ello, se produce un comportamiento semejante al del D como producto de aprendizaje no-social, por ejemplo, aprendizaje del tipo ensayo y error. En esta situación los O sólo “se comportan respecto de” los estímulos discriminativos o partes del ambiente que son tocados o marcados con residuos orgánicos por los D; no reproducen la forma del comportamiento de éstos. La semejanza comportamental se produce por el seguimiento de los estímulos discriminativos provistos por la situación y no por la reproducción del comportamiento del D.
Copia fue otro término utilizado por Miller y Dollard (1941) para subrayar una diferencia respecto del comportamiento dependiente de la igualación. La diferencia está en que en la copia, supuestamente, el animal es sensible a la relación igual/diferente entre su comportamiento y el del modelo o D. Esta situación podría representar un refuerzo diferencial del comportamiento; sin embargo, el implicar la existencia de un procedimiento de refuerzo diferencial por parte de los animales no se ha demostrado (Galef, 1988). En la copia, los animales igualan el comportamiento de otros (por ejemplo, el canto en los pájaros), y no se puede identificar una consecuencia para la igualación ni explicar el por qué se mantiene este comportamiento, o si son sensibles o no a la relación igual/diferente.
Por otro lado, está el aprendizaje de relaciones respuesta-reforzador o emulación de meta, extraídas de los comportamientos presentados por el D y emitidos después por el O (Whiten & Ham, 1992).
Otro término utilizado es imitación. Para diferenciarlo de los anteriores, referidos a las respuestas, se menciona la opinión de diversos autores. Suboski y Bartashunas (1984) puntualizaron, respecto de los comportamientos que se adquieren observando a coespecíficos, que Klopfer (1961) ya había señalado que algunas de las respuestas producidas en lo social son transitorias, mientras que las imitativas permanecen como un cambio relativamente permanente. Esto es, en el caso de los comportamientos facilitados en el ámbito social, ambas se presentan en un tiempo t1, mientras que en los comportamientos imitativos, la ejecución del comportamiento aprendido se da en el tiempo t2. Este aspecto podría ser un factor que diferencie la imitación de otros tipos de comportamiento en el cual interviene un coespecífico. Una condición más para que un comportamiento se considere aprendido por medio de la imitación, es que sea nuevo en el repertorio del O. Estos aspectos &separar los momentos de la demostración y el de la prueba, y considerar lo novedoso del comportamiento& pueden diferenciar los comportamientos aprendidos en el terreno social como la imitación, de otros tipos de comportamientos que sólo son mediados socialmente (Heyes, 1993, 1994, 1996; Whiten & Ham, 1992).
Como ya se mencionó, esta abundancia de conceptos no parece indicar que haya una relación entre ellos.
Modelo para analizar el aprendizaje social
En esta sección se propone cómo organizar lo que ocurre en una situación de aprendizaje social y la forma en que los conceptos se podrían analizar de una manera integrada.
Una manera de aproximarse a esta integración puede ser aplicando la idea de sistema de comportamiento propuesto en el modelo de J. A. Hogan (1994b). Basado en su modelo para analizar problemas de desarrollo y la organización del comportamiento, Hogan (1988) propuso un modelo para analizar el sistema motivacional de hambre de los pollos domésticos. Hogan (1994a), al hablar del concepto de causa en el estudio del comportamiento, propuso un sistema estructural de comportamientos en el que se incluyen mecanismos perceptuales, centrales y motores. En este sistema también podrían incluirse las estructuras cognoscitivas como ideas, pensamientos o memorias. Aunque este sistema puede considerarse como enmarcado en el estudio cognoscitivo del comportamiento, su importancia radica en que permite organizar los hallazgos relacionados con un aspecto del comportamiento, en este caso el aprendizaje social, el aprendizaje observacional y la imitación.
De manera semejante, van Kampen (1996) propone un modelo para analizar la impronta filial basado en el modelo de Hogan (1988). La Figura 1 &reproducida del artículo de Hogan (1994b) & muestra una representación gráfica de dos sistemas de comportamiento y sus componentes. Cada uno de éstos se integra por varios niveles cuyas conexiones pueden volverlos realmente complejos. Las conexiones propuestas podrían tener una relación isomórfica con las conexiones del sistema nervioso, aunque esto no implica centros localizados en el mismo. El modelo está formado por mecanismos perceptuales en la parte de entrada, los cuales resuelven el problema de reconocimiento de estímulos. En la parte de salida hay mecanismos motrices, responsables de coordinar los movimientos musculares que forman el comportamiento. Así, los patrones de movimiento que observamos son manifestación de los mecanismos motrices del sistema. Los mecanismos centrales del sistema de comportamiento integran la entrada de varios mecanismos perceptuales y coordinan la activación de los mecanismos motores. La actividad de un mecanismo central de un sistema de comportamiento se llama a menudo el estado interno o la motivación del animal. Además de que este modelo permite organizar los hallazgos de algún aspecto comportamental, el relacionarlo con una organización del ambiente podría dar una visión más clara de un acontecimiento comportamental como el aprendizaje social.
De acuerdo con Hogan (1994a), un sistema de comportamiento puede ser considerado como una descripción de la estructura del comportamiento y puede ser definido como cualquier organización de mecanismos perceptuales, centrales y motrices que funcionan como unidad en algunas situaciones. A su vez, el autor define a los mecanismos de comportamiento como unidades básicas que pueden ser motrices, centrales y perceptuales. La calidad de mecanismo está dada porque su activación tiene como resultado un evento de interés comportamental, esto es, un patrón motriz específico, un estado interno identificable o como una percepción en particular. Características adicionales de los mecanismos son que pueden ser estructuras cognoscitivas y pueden incluir entidades tales como ideas, pensamientos y memorias. Aun cuando el autor mencionado define a los mecanismos comportamentales como estructuras en el sistema nervioso, señala que éstas no involucran un estudio neurofisiológico. Como una característica adicional, la definición de Hogan de un sistema de comportamiento está dada en términos estructurales, en contraste con lo que sería una definición enunciada en términos funcionales. El autor diferencia estas definiciones poniendo como ejemplo al comportamiento sexual. Una definición estructural del comportamiento sexual incluiría la descripción de los procesos perceptuales que analizan los estímulos, activan el mecanismo central de la coordinación sexual y también añadiría la descripción de los patrones motrices que ocurren cuando se activa el mecanismo central. En contraste, una descripción funcional del comportamiento sexual enfatizaría, sobre todo, la reproducción y los comportamientos que condujeran a la propagación exitosa de la especie. De acuerdo con estas aclaraciones, el aprendizaje por observación, el aprendizaje social y la imitación pueden ser considerados como sistemas de comportamiento, ya que funcionan como una unidad y a su vez, estas unidades pueden estar formadas por una percepción, un estado interno y un patrón motriz específico. Hogan señala además, que los mecanismos comportamentales son estructuras cognoscitivas. Este aspecto está íntimamente relacionado con las características del aprendizaje por observación que menciona Bandura (1969). Bandura señala que es necesario que un mecanismo cognoscitivo medie el tiempo entre una presentación comportamental de un modelo y la subsiguiente ejecución del comportamiento observado.
ANÁLISIS CONCEPTUAL DEL APRENDIZAJE OBSERVACIONAL Y LA IMITACIÓN
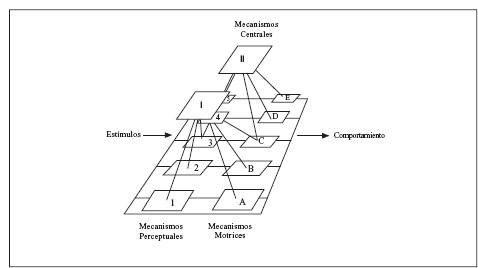
Figura 1. Esquematización de los sistemas de comportamiento. Los estímulos del mundo externo, incluyendo los producidos por el comportamiento, se analizan por los mecanismos perceptuales. La salida de los mecanismos perceptuales puede integrarse por los mecanismos centrales y/o ser canalizada directamente por los mecanismos motrices. La salida de los mecanismos motrices resulta como comportamiento. En este diagrama el Mecanismo Central I, los Mecanismos Perceptuales 1, 2 y 3 y los Mecanismos Motrices A, B y C, forman un sistema de comportamiento; el Mecanismo Central II, los Mecanismos Perceptuales 3, 4 y 5 y los Mecanismos Motrices C, D y E forman un segundo sistema de comportamiento. Las unidades 1-A, 2-B y los siguientes pueden ser considerados sistemas de comportamiento menos complejos. Adaptado de Hogan (1988)
Una forma de aplicar la noción de sistema de Hogan al aprendizaje social involucra el análisis de los insumos de estímulo y los productos motrices. Entre estos dos extremos, los mecanismos centrales coordinan los elementos perceptuales y producen respuestas motrices. Cada uno de estos sectores del modelo, perceptual y motriz tienen a su vez diferentes niveles que organizan las acciones de sus elementos. Por un lado, los elementos perceptuales permiten, primero, efectuar el reconocimiento de los objetos y, después, la función de los objetos estímulo. Los mecanismos motrices integran las respuestas y al final producen un patrón motriz. En medio de estos mecanismos se encuentran los mecanismos centrales.
En la Figura 2 puede verse una propuesta para analizar, con base en el modelo de Hogan (1994b), el insumo perceptual y la producción de mecanismos motrices que se producen en un episodio de aprendizaje social. En la sección de mecanismos perceptuales se incluyen los cambios en el ambiente que pueden incidir en el O durante este episodio. Los estímulos que percibe pueden ser de tipo olfatorio (Ol), visual &color o contraste & (V), gustativo (S) y motriz (M). El O tiene la posibilidad de reconocer los rasgos de estos diferentes estímulos en el primer nivel del modelo, en la parte correspondiente a los mismos. A partir de ellos puede reconocer los objetos del conjunto y determinar si el estímulo es alimento (A), un coespecífico o un depredador (Pr). En el tercer nivel del modelo se reconoce la función del estímulo percibido y el O puede distinguir si el estímulo es un indicador de alimento al que puede aproximarse y puede cumplir la función de comer (Ca) o si es un peligro del que debe alejarse (P). Estos aspectos pueden estar sujetos a la intervención de mecanismos centrales que conducen a la emisión de una respuesta. En el caso de la imitación, que implica aprendizaje acerca de la forma del comportamiento de un coespecífico percibido de manera visual, debe participar un transductor (T) que permita transformar los movimientos observados en acción o comportamiento. Los resultados de un experimento de Pellegrino, Fadiga, Fogassi, Gallese y Rizzolatti (1992), sugieren que en la corteza premotora en macacos (Macaca nemestrina) existen neuronas sensibles al “significado” de las acciones. Pellegrino y cols. encontraron que algunas neuronas, que se activaban cuando los sujetos hacían un movimiento de la mano, también se activaban cuando observaban al experimentador realizar movimientos semejantes. Este tipo de respuestas de la corteza premotora parece ser un elemento que permite suponer la existencia de tal transductor en algunos de los sujetos en los que se produce la imitación. Por su parte, Iacoboni y cols.. (2000), indicaron que también en humanos se produce esta activación de neuronas en el surco temporal superior, y que es máxima durante la imitación, esto es, en la condición en la que hay congruencia entre la acción observada y la acción que va a ser ejecutada. Por otro lado, Miklósi (1999b) mencionó que el sistema de igualación observación- ejecución propuesto por Rizzolatti y Arbib (1998) provee una teoría general que explica los fenómenos imitativos y los coloca en un contexto evolutivo. Estos autores concuerdan con Miklósi (Rizzolatti & Arbib, 1999).

Figura 2. El modelo de desarrollo presentado por Hogan (1994b) presenta los mecanismos perceptuales, centrales y motrices. T representa al transductor; las funciones: Ca, comida; P, peligro; la integración motriz: Co, alimentación; Hu, huir; los objetos: A, alimento; Pr, depredador; los patrones motrices Cr, comer; H, huir; Al, agredirlo; I, imitar; los tipos de estímulos: Ol, olfativo; V, visual S, gustativo y M, movimiento. En la parte inferior los estímulos y la conducta. Los elementos con línea continua muestran los aspectos ya desarrollados y los de línea punteada, aquellos que pueden desarrollarse.
La integración motriz puede presentarse a un nivel en el que no intervengan los mecanismos centrales &como lo indica la línea continua de la Figura 2& y dependiendo de los estímulos, los rasgos, los objetos y la función de los mismos, puede producirse alimentación (Co) o huída (Hu). No obstante, puede intervenir el transductor cuando se involucra el reconocimiento del movimiento y la reproducción del mismo observado en un coespecífico. El patrón motriz puede ser de diversos tipos respecto del ambiente y el animal, O puede aproximarse al alimento y comer (Cr) como patrón motriz, alejarse del depredador o peligro y huir (H) o agredirlo (Al) en respuesta. Si lo que percibe y reconoce son los movimientos o el comportamiento de un coespecífico, con la intervención del transductor, a nivel central, puede imitar (I) los movimientos observados en el coespecífico. Este análisis indica que los diferentes conceptos relativos al aprendizaje observacional se refieren a diferentes aspectos de situaciones en las que el O recibe información de diferentes aspectos del ambiente o del movimiento de un coespecífico. Por ejemplo, si reconoce aspectos físicos de la situación y del alimento y la función de éste, puede alimentarse pero si esto ocurre en t1, se produce lo que se clasificó arriba como resaltamiento de estímulo. Por otro lado, si todo ocurre en t1, es decir si percibe y reconoce los movimientos del coespecífico, y reproduce respuestas que ya existen en su repertorio, se produce lo que se identificó como facilitación de respuesta. Si reconoce el comportamiento y emite en t1 una respuesta que ya existe en su repertorio, el resultado es contagio o la emisión de la misma respuesta. Sin embargo, si el transductor participa y transforma lo observado en movimientos, puede imitar a un coespecífico en t2 con una respuesta nueva en su repertorio, lo que implica, como mencionó Klopfer (1961) que un aspecto que determina que se produzca imitación es que el comportamiento se reproduzca en el t2 y como mencionan Zentall (1996) y Byrne (1994) que la respuesta reproducida sea nueva en el repertorio del O. Al parecer, la pertinencia de las explicaciones señaladas depende de lo que la situación de aprendizaje social le permite o le exige al O. Para que realmente se produzca imitación en una situación controlada, se debe asegurar que el procedimiento de demostración permite que el O reconozca movimientos del D y no sólo los aspectos físicos de los estímulos presentes y los del coespecífico.
Unos ejemplos de estos casos son los estudios de Galef y Buckley (1996) y los estudios de Laland y Plotkin (1991, 1993). En el primero de ellos se hicieron seis experimentos en los que se evaluó el efecto de colocar ratas que dejaran senderos marcados con residuos orgánicos en un laberinto en forma de T. Los autores encontraron que las ratas seguían los caminos marcados por las ratas líderes. Este tipo de comportamiento social identificado como resaltamiento del estímulo puede parecer semejante a la imitación. Sin embargo, las ratas seguidoras sólo se guiaban por las marcas dejadas por las líderes. A primera vista podría parecer que están presentando el mismo comportamiento que las líderes, pero sólo están siguiendo señales olfatorias en el laberinto. En el segundo par de estudios, Laland y Plotkin colocaron animales que dejaron residuos orgánicos, heces y orina, en la mitad de una caja en forma de U en ángulo recto, esto es una U cuadrada. Veinticuatro horas más tarde colocaron animales ingenuos y alimento en los dos extremos de la U. Encontraron que los sujetos ingenuos prefirieron alimentarse en el lado que estaba marcado con residuos orgánicos. Este comportamiento puede parecer, si no se conoce el diseño del experimento, que los sujetos ingenuos están comportándose como los líderes. Sin embargo, sólo están alimentándose en la sección marcada de la caja experimental.
Ejemplos de este tipo de situación, facilitación social, ocurren cuando los animales se alimentan en grupos y una situación de amenaza se presenta. Los animales presentan la misma respuesta, una respuesta ya existente en su comportamiento y puede parecer a primera vista que de alguna manera están copiando a los otros animales. En este caso pueden observar el comportamiento de sus vecinos y realizar las mismas acciones, acciones que no son nuevas en el grupo de respuestas individuales.
Ejemplos de imitación los encontramos en diversos estudios. En este caso se utiliza el método de dos acciones. En este método se utilizan dos demostradores que están entrenados a realizar una acción diferente cada uno. En la fase de demostración, se les coloca a demostrar individualmente la acción que han aprendido a un observador ingenuo. El aparato utilizado está construido de tal forma que puede producir alimento con dos manipulaciones diferentes, que corresponden a las acciones que cada uno de los demostradores domina. En la fase de prueba, el observador debe realizar la acción que su respectivo demostrador presentó, no sólo “abrir” el alimentador. Si ejecuta la acción que le fue demostrada, se puede concluir que la repitió como resultado de haber observado cuidadosamente los movimientos del demostrador. Sólo en este caso se puede decir que se produjo imitación. Como el observador es un sujeto ingenuo, la respuesta o acción evidentemente es nueva en su conjunto de comportamientos.
En el primer ejemplo de este comportamiento, Zentall, Sutton y Sherburne (1996) evaluaron la imitación en palomas (Columba livia). Utilizaron el método de dos acciones en el que las palomas podían obtener alimento de dos formas, pisando la palanca o picoteándola. Encontraron que las palomas realizaron la misma acción que su respectivo demostrador, esto es, la imitaron. En un segundo estudio, Akins y Zentall (1996) repitieron el mismo procedimiento, pero en este caso, utilizando codornices japonesas (Coturnix japonica). Los autores encontraron el mismo resultado, las codornices imitaron al demostrador ejecutando la acción correspondiente. En el tercer estudio, Akins, Klein y Zentall (2002) reportaron imitación en la codorniz japonesa (Coturnix japonica). En este estudio utilizaron las acciones de mover una malla de alambre para tener acceso al alimento. Los dos demostradores aprendieron una sola forma de la acción, abrir el alimentador empujando la malla hacia la derecha o hacia la izquierda.
Otra técnica que se ha utilizado para determinar si se produce imitación en chimpancés (Pan troglodytes) es la de entrenar deliberadamente algunas acciones y después evaluar el efecto de presentar acciones novedosas adicionales. En el estudio de Custance, Whiten y Bard (1995), dos chimpancés imitaron 13 y 17 acciones, respectivamente. Se les entrenó en 15 acciones y después se les presentaron 48 acciones novedosas. Los autores señalaron que los chimpancés imitaron las acciones novedosas sin recibir entrenamiento deliberado. La chimpancé llamada Katrina imitó 13 acciones y el chimpancé llamado Scott 17. En este estudio se registraron las respuestas mediante dos observadores independientes. Encontraron que el acuerdo entre ellos fue estadísticamente significativo.
El modelo propuesto para analizar el aprendizaje social parece indicar que los diferentes conceptos que se han utilizado para explicar estos aspectos se refieren a fuentes diversas de control del estímulo. El uso de dicho análisis evita dar nombres diferentes para cada fuente de control del estímulo que participa en el desarrollo del aprendizaje social y señala que hay una relación entre estos aspectos. Adicionalmente, se requiere describir la manera en que el comportamiento social podría mantenerse tras ser adquirido. Después de este momento, el comportamiento social está sujeto a los cambios que le produzcan las consecuencias que dichos comportamientos reciban en forma posterior a su emisión (Galef, 1995). Al mencionar cómo es que pueden perdurar las respuestas aprendidas en lo social, Galef utiliza dos conceptos propuestos por Staddon y Simmelhag (1971), quienes indicaron que el comportamiento, haciendo una analogía con la teoría neo-darwinista de la evolución, está sujeto a los principios de variación y selección. El aprendizaje social es un factor de variación, pues genera comportamiento nuevo, como la variación genética. Una vez producida dicha variación comportamental ésta puede estar sujeta a los principios selectivos del refuerzo, el cual evita que un comportamiento seleccionado desaparezca, como la selección del más apto. De acuerdo con Staddon y Simmelhag, el refuerzo selecciona los comportamientos producidos por la variación y evita que desaparezcan del repertorio del organismo. Esta selección funciona como un proceso de tipo no-social que asegura la fidelidad y la longevidad de los comportamientos aprendidos socialmente. En esta forma se pueden organizar los diversos conceptos utilizados en el estudio del aprendizaje social y describir la producción y el mantenimiento del comportamiento social.
Factores que determinan el aprendizaje social
En esta sección se analizarán algunos de los factores que determinan el aprendizaje social. Algunos autores han mencionado factores de manera aislada, por ejemplo, el cambio de valencia de los estímulos (Hogan, 1988) o la potenciación (Cabrera & Nieto, 1993). Conceptos que se revisarán de manera individual y se indicará cómo pueden llegar a intervenir en el aprendizaje social.
La presencia de un coespecífico en una situación puede producir un cambio en la importancia que ella tiene para un O, y este cambio de relevancia puede ser producto de la presencia del D en esa situación, a lo cual se le ha llamado en algunos escritos relacionados con el aprendizaje social, cambio de la valencia del estímulo acompañado por el D. La presencia del D hace que la evaluación de ese estímulo cambie. D. E. Hogan (1988) menciona lo que caracteriza al cambio de valencia de la siguiente manera:
“... la hipótesis de la transformación de valencia sostiene que el comportamiento de un demostrador puede funcionar como estímulo “instruccional” que enfoca la atención del observador hacia una relación predictiva entre un estímulo hedónicamente neutral y un evento biológicamente significativo (i. e., una contingencia pavloviana estímulo-refuerzo). [Además explica.] El proceso instruccional no implica una relación encubierta de parte del demostrador para enseñar al observador hacia dónde mirar o qué hacer con los estímulos con los que entra en contacto. En su lugar, la acción abierta del demostrador es funcional en el sentido de que causa que el observador aprenda acerca de una contingencia estímulo-reforzador que de otra manera podría ser descubierta independientemente por el observador” (véase Zentall & Hogan, 1976, p. 226).
Este término no explica a qué se debe el cambio, sólo le da un nombre. La función del comportamiento del D en el cambio de valencia podría considerarse como un estímulo condicional (Cumming & Berryman, 1965; Lashley, 1938) para la relación predictiva mencionada por Zentall y Hogan. Esto es, el comportamiento del D funciona como un estímulo que determina cierta función de la relación predictiva mencionada. Este efecto podría presentarse también para una relación entre respuestas además de en la relación entre estímulos mencionada por esos autores.
Los cambios en la función de los elementos estímulo y respuesta observados en los experimentos de aprendizaje social y que no están involucrados directamente con el sistema alimentario, se han explicado en términos de potenciación (Cabrera & Nieto, 1993). La potenciación fue descrita por Rusiniak, Hankins, García y Brett (1979) en un estudio en el que las ratas no asociaron el olor de esencia de almendra con el efecto tóxico del cloruro de litio; sin embargo, cuando el olor se les presentó en compañía del sabor dulce de la sacarina, el olor se hizo fuertemente aversivo (LoLordo & Droungas, 1989). En un análisis de la potenciación, LoLordo y Droungas (1989) presentaron dos teorías alternas para explicarla: la primera, la teoría entre compuestos, sugiere que cada uno de los elementos de un compuesto se asocia con un estímulo incondicionado que forma asociaciones individuales. El compuesto también forma asociaciones con el estímulo incondicionado. Como consecuencia, al presentar uno de los componentes del compuesto se establece una activación doble de la representación del estímulo incondicionado. Acorde con esta teoría, la potenciación del condicionamiento del olor, primer estímulo condicionado, depende de manera importante de la formación y mantenimiento de la asociación sabor-enfermedad, la asociación del segundo estímulo condicionado con el estímulo incondicionado. La fuerza del efecto asociativo del olor debería no sólo reflejar la fuerza de las asociaciones entre el olor y el sabor, sino también la fuerza de la asociación entre el sabor y la enfermedad. Como consecuencia de esto, cualquier disminución de la asociación entre los estímulos debería reducir la potenciación.
La segunda teoría de la potenciación que presentaron LoLordo y Droungas (1989) fue llamada canalización sensorial y de compuerta. Dicha teoría supone que los estímulos gustativos “pertenecen” a un sistema interno o visceral, de allí que sea fácil establecer asociaciones entre las señales gustativas y el malestar gastrointestinal. Las señales exteroceptivas &como luces y sonidos& “pertenecen” a otro sistema externo de defensa. Algunos estímulos, como el olor, son relevantes para ambos sistemas, tanto el interno como el externo, porque el olor puede ser una propiedad de un alimento tóxico o de un depredador que se aproxima. En ausencia de una señal de sabor, el olor se dispara (is gated) al sistema externo y exhibe propiedades semejantes a las de los estímulos exteroceptivos. Bajo tales condiciones, el olor se asocia con estímulos aversivos y/o dolorosos externos. Contrariamente, en presencia de señales relacionadas con la alimentación, por ejemplo, el sabor de los alimentos, el olor se dispara al sistema de alimentación y exhibe propiedades semejantes a aquellas naturales del sabor.
En términos semejantes podría considerarse al efecto de un coespecífico con propiedades diferentes a las del alimento, y que puede disparar a un aspecto del ambiente o del comportamiento del D hacia un sistema interno de alimentación o a uno de afiliación (Bolhuis & Honey, 1994). En esta situación, el comportamiento social del sujeto observador se dirige hacia un estímulo, ya sea parte del ambiente o la forma de un comportamiento y cambia su relevancia para el observador. Un ejemplo de esto es lo que ocurre con los pollos cuando se les presenta un estímulo arbitrario, verbigracia, un objeto geométrico acompañado con componentes auditivos propios del sistema de afiliación, como una grabación con el llamado maternal de una gallina.
Este aspecto de la potenciación requiere de más estudios que permitan determinar si el coespecífico dirige las características de un aspecto del ambiente o del comportamiento de un D hacia un sistema de alimentación o a un sistema de afiliación en una situación de aprendizaje social. En el caso de la potenciación, así como en el cambio de valencia, el coespecífico puede funcionar como un estímulo condicional que cambia la función de las relaciones y la relevancia de los estímulos físicos, así como las respuestas que lo acompañan.
El aprendizaje social puede analizarse, también, como una discriminación condicional del tipo disposición de la ocasión. En los experimentos de aprendizaje social, las relaciones entre estímulos y respuestas pueden ser más complejas que en las de aprendizaje individual. Fragaszy (1998) señala que en este tipo de experimentos &que involucran un elemento de aprendizaje social& se produce una situación parecida a aquella en la que se observa disposición de la ocasión (Holland, 1983, 1985; Holland & Ross, 1981; Schmajuk, Lamoureux & Holland, 1998; Ross & Holland, 1981; Skinner, 1938). Sin embargo, el paradigma de disposición de la ocasión, estudiado ampliamente por Holland y sus colaboradores, involucra un manejo preciso de las relaciones entre los estímulos rasgo (feature) y blanco (target). Cualquier alteración de la simultaneidad, configuración o en la secuencia de presentación produce cambios concomitantes en el condicionamiento. Este aspecto de la presentación de los estímulos de manera ya sea secuencial o simultánea, no se ha controlado de manera precisa en los experimentos de aprendizaje social. La interacción entre los estímulos y los demostradores son un tanto aleatorias, si se les compara con los procedimientos en los que se analiza la disposición de la ocasión. Las situaciones en las que el aprendizaje social produce relaciones estímulo-estímulo podrían analizarse con base en el paradigma de disposición de la ocasión, siempre que se hubiera hecho un manejo preciso de la presentación de los estímulos físicos involucrados y su relación con los demostradores. En el análisis de la disposición de la ocasión que hizo Holland (1985), consideró a ésta como una discriminación condicional tomando en cuenta que tanto la disposición de la ocasión como la discriminación condicional resultan sensibles a las alteraciones producidas en el hipocampo. Esta sensibilidad no se presenta en los procedimientos de condicionamiento clásico simple, no condicionales a la presentación de otro estímulo. Como se señaló arriba, en el aprendizaje social con animales no-humanos como sujetos, los demostradores pueden funcionar como estímulos condicionales, esto es, indicar al observador una relación entre estímulos que de otra forma no percibiría o la percibiría mediante el aprendizaje individual, sin la participación del coespecífico. Al introducir un demostrador, la situación presentada al observador cambia de una situación en la que es expuesto a relaciones entre estímulos o relaciones entre estímulos y respuestas, a una más compleja en la que la presencia del demostrador, por el solo hecho de presentarse, afecta la relación entre estímulos o entre estímulos y respuestas. Al aplicar la noción de disposición de la ocasión al aprendizaje social podemos considerar que el demostrador o modelo puede funcionar de manera análoga a un estímulo rasgo y realizar un efecto modulador (Swartzentruber, 1995). Este efecto modulador se manifiesta afectando la relación entre un estímulo de la situación, como podría ser un sabor característico del alimento consumido por el demostrador, una acción o comportamiento y la relación de alguno de ellos con un estímulo incondicionado o con un refuerzo. Este efecto modulador afectaría la relación entre estímulos y la relación entre estímulos y respuestas reduciendo el umbral para que se produzcan asociaciones entre los estímulos o las respuestas con un estímulo incondicionado o con un refuerzo, en una situación de aprendizaje social.
Hallazgos recientes (Kogan, Frankland & Silva, 2000; Kogan, Stern, Coblentz & Silva, 1998) indican también que el reconocimiento social y la transmisión social de preferencia alimenticia &ambas formas de aprendizaje social&, son sensibles a las alteraciones de la síntesis de proteínas en el hipocampo, igual que sucede con la ejecución bajo procedimientos del tipo disposición de la ocasión que es sensible a las lesiones en esta estructura cerebral. Los paradigmas de aprendizaje social que involucran el aprendizaje de relaciones respuesta-estímulo pueden considerarse, asimismo, como discriminaciones condicionales en las que el coespecífico, por sí mismo, agrega un aspecto adicional a la forma o topografía de su comportamiento y cambia la relevancia de la relación para el observador. En aquellos casos en los cuales se presentan relaciones estímulo-estímulo y respuesta-estímulo, el coespecífico funciona como un estímulo condicional a la relación entre estímulos del primer caso y a la relación respuesta-estímulo del segundo caso. En ambos, el coespecífico puede funcionar como un productor de una valencia positiva en las relaciones percibidas por el observador. Tomando en cuenta que un estímulo se potencia positivamente cuando se presenta con un coespecífico (Cabrera & Nieto, 1993), o cuando un estímulo olfativo o visual se presenta con uno gustativo (Droungas & LoLordo, 1991; Galef & Osborne, 1978; LoLordo & Droungas, 1989; Thiele & Frieman, 1994), la simple presencia del coespecífico hace que, en la transmisión de preferencias alimenticias (Galef, 1985), la aversión por rechazo directo a un estímulo presentado con un coespecífico sea difícil de lograr. En algunos experimentos de transmisión social de preferencia alimenticia, tal vez se ha producido un cambio de valencia en los casos en los que se incrementa la preferencia por un alimento característico (D. E. Hogan, 1988). Este resultado puede considerarse como un aprendizaje de relaciones estímulo-estímulo en las que el coespecífico funciona como el estímulo condicional. De manera específica, en las situaciones en las que se produce un aumento en la preferencia por un alimento con características particulares, el coespecífico funcionó como un estímulo “instruccional” que condicionalmente indicó a los observadores que el estímulo alimento con diversas características es seguro para consumirse. En los tres casos &cambio de valencia, potenciación y disposición de la ocasión& el coespecífico funciona como un estímulo condicional que cambia la relevancia de la relación entre estímulos o entre respuestas que lo acompaña.
CONCLUSIONES
La contribución principal del análisis presentado consiste en proponer el uso del modelo de J. A. Hogan (1988, 1994b) para organizar los diferentes aspectos que se han estudiado en el aprendizaje social. Dicha propuesta no pretende sugerir que alguna aproximación al estudio del comportamiento social, ya sea cognoscitivo o comportamental, sea más completa ni se propone tampoco un planteamiento ecléctico. El propósito de este análisis fue tratar de señalar cómo los aspectos de una situación en la que se aprende el comportamiento social están relacionados, amén de señalar que pueden considerarse de manera conjunta. El estudio provee un marco de referencia para formular y analizar los problemas de manera organizada (van Kampen, 1996). De la organización que permite ese modelo se puede colegir que los diferentes conceptos se refieren a diferentes fuentes de control del estímulo más que a diferentes procesos comportamentales. Este análisis permitiría proveer una visión integrada de estas situaciones; además, la hipótesis del transductor parece indicar que se están descubriendo las conexiones internas del modelo de entrada-salida aplicado al estudio del comportamiento.
Por otro lado, se señaló que un paradigma de discriminación condicional está tras las explicaciones de cómo se aprenden los comportamientos sociales, como son el cambio de valencia, la potenciación y la disposición de la ocasión. El análisis propuesto no involucra la participación de procesos diferentes al aprendizaje asociativo tradicional.
Referências
Akins, C. K. & Zentall, T. R. (1996). Imitative learning in male Japanese quail (Coturnix japonica) using the two-action method. Journal of Comparative Psychology, 110, 316-320. [ Links ]
Akins, C. K., Klein, E.D. & Zentall, T. R. (2002). Imitative learning in Japanese quail (Coturnix japonica) using the bidirectional control procedure. Animal Learning and Behavior, 30, 275-281. [ Links ]
Bandura, A. (1969). Principles of behavior modification. Nueva York: Holt, Rinehart & Winston, Inc. [ Links ] Bolhuis, J. J. & Honey, R. C. (1994). Within-event learning during filial imprinting. Journal of Experimental Psychology: Animal Behavior Processes, 20, 240-248. [ Links ]
Byrne, R. W. (1994). The evolution of intelligence. En P. J. B. Slater & T. R. Halliday (Eds.), Behaviour and evolution (pp. 223-264). Inglaterra: Cambridge University Press. [ Links ]
Cabrera, R. & Nieto, J. (1993). Potenciación de preferencias alimenticias en crías mediante su interacción con la rata madre. Psicológica, 14, 151-160. [ Links ]
Cook, M., Mineka, S., Wolkenstein, B. & Laitsch, K. (1985). Observational conditioning of snake fear in unrelated rhesus monkeys. Journal of Abnormal Psychology, 94, 591-610. [ Links ]
Coussi-Korbel, S. & Fragaszy, D. M. (1995). On the relation between social dynamics and social learning. Animal Behaviour, 50, 1441-1453. [ Links ]
Cumming, W. W. & Berryman, R. (1965). The complex discriminated operant: Studies of matching-to-sample and related problems. En D. I. Mostofsky (Ed.), Stimulus generalization (pp. 284-330). Stanford, CA: Stanford University Press. [ Links ]
Custance, D. M., Whiten, A. & Bard, K. A. (1995). Can young chimpanzees (Pan troglodytes) imitate arbitrary actions? Hayes / Hayes (1952) revisited. Behaviour, 132, 835-859. [ Links ]
Davis, J. M. (1973). Imitation: A review and critique. En P. P. G. Bateson & P. H. Klopfer (Eds.), Perspectives in ethology (pp. 43- 72). Nueva York: Plenum. [ Links ]
Droungas, A. & LoLordo, V. M. (1991). Taste-mediated potentiation of odor aversion induced by lithium chloride: Effects of preconditioning exposure to the conditioned stimulus and postconditioning extinction of the taste aversion. Learning and Motivation, 22, 291-310. [ Links ]
Fragaszy, D. M. (1998, junio). The ties that (might) bind. Presentado en Social Learning and Cultural Transmission: From Invertebrates to Great Apes and Humans Towards a Biological Synthesis, Nápoles. [ Links ]
Galef, B, G. Jr. (1985). Direct and indirect behavioral pathways to the social transmission of food avoidance. En P. Bronstein & N. S. Braveman (Eds.), Experimental assessments and clinical applications of conditioned food aversions (pp. 203-215). Nueva York: Annals of the New York Academy of Sciences,. [ Links ]
Galef, B. G. Jr. (1988). Imitation in animals: history, definition and interpretation of data from the Psychological Laboratory. En T. R. Zentall & B. G. Galef (Eds.), Social learning: Psychological and biological perspectives (pp. 3-28). Hillsdale, NJ: Earlbaum. [ Links ]
Galef, B. G., Jr. (1995). Why behaviour patterns that rats learn socially are locally adaptive. Animal Behaviour, 49, 1325-1334. [ Links ]
Galef, B. G., Jr. & Buckley, L. L. (1996). Use of foraging trails by Norway rats. Animal Behaviour, 51, 765-771. [ Links ]
Galef, B. G. Jr. & Osborne, B. (1978). Novel taste facilitation of the association of visual cues with toxicosis in rats. Journal of Comparative and Physiological Psychology, 92, 907-916. [ Links ]
Heyes, C. M. (1993). Imitation, culture and cognition. Animal Behaviour, 46, 999-1010. [ Links ]
Heyes, C. M. (1994). Social learning in animals: Categories and mechanisms. Biological Reviews, 69, 207-231. [ Links ]
Heyes, C. M. (1996). Introduction: Identifying and defining imitation. En C. M. Heyes & B. G. Galef, Jr. (Eds.), Social learning in animals: The roots of culture (pp. 211-220). San Diego, CA: Academic Press. [ Links ]
Heyes, C. M. & Dawson, G. R. (1990). A demonstration of observational learning in rats using a bidirectional control. Quarterly Journal of Experimental Psychology, 42B, 59-71. [ Links ]
Heyes, C. M. & Galef, Jr., B. G. (1996). Social learning in animals: The roots of culture. San Diego, CA: Academic Press. [ Links ]
Hogan, D. E. (1988). Learned imitation in pigeons. En T. R. Zentall & B. G. Galef (Eds.), Social learning: Psychological and biological perspectives (pp. 225-238). Hillsdale, NJ: Earlbaum. [ Links ]
Hogan, J. A. (1988). Cause and function in the development of behavior systems. En E. M. Blass (Ed.), Handbook of behavioral neurobiology (Vol. 9, 63-106). Nueva York: Plenum. [ Links ]
Hogan, J. A. (1994a). The concept of cause in the study of behavior. En J. A. Hogan & J. J. Bolhuis (Eds.), Causal mechanisms of behavioural development (pp. 3-15). Cambridge University Press. [ Links ]
Hogan, J. A. (1994b). Structure and development of behavior systems. Psychonomic Bulletin and Review, 1, 439-450. [ Links ]
Holland, P. C. (1983). Occasion-setting in Pavlovian feature positive discriminations. En M. L. Commons, R. J. Herrnstein & A. R. Wagner (Eds.), Quantitative analyses of behavior: Discrimination processes (Vol. 4, pp. 183-206). Nueva York: Ballinger. [ Links ]
Holland, P. C. (1985). The nature of conditioned inhibition in serial and simultaneous feature negative discriminations. En R. R. Miller & N. E. Spear (Eds.), Information processing in animals: Conditioned inhibition (pp. 267-298). Hillsdale, NJ: Lawrence Erlbaum. [ Links ]
Holland, P. C. & Ross, R. T. (1981). Within-compound associations in a serial compound conditioning. Journal of Experimental Psychology: Animal Behavior Processes, 7, 228-241. [ Links ]
Iacoboni, M., Koski, L. M., Brass, M., Bekkering, H., Woods, R. P., Dubeau, M. C., Mazziotta, J. C. & Rizzolatti, G. (2000). Reafferent copies of imitated actions in the right superior temporal cortex. Proceedings of the National Academy of Sciences, 98, 13995-13999. [ Links ]
Klopfer, P. H. (1961). Observational learning in birds: The establishment of behavioral modes. Behaviour, 17, 71-79. [ Links ]
Kogan, J. H., Frankland, P. W. & Silva, A. J. (2000). Long-term memory underlying hippocampus-dependent social recognition in mice. Hippocampus, 10, 47-56. [ Links ]
Kogan, J. H., Stern, J. H., Coblentz, J. & Silva, A. J. (1998, junio). Parallel properties of memory and long-term potentiation in CREBaD- mutant mice. Cartel presentado en Social Learning and Cultural Transmission: From Invertebrates to Great Apes and Humans Towards a Biological Synthesis, Nápoles. [ Links ]
Laland, K. N. & Plotkin, H. C. (1991). Excretory deposits surrounding food sites facilitate social learning of food references in Norway rats. Animal Behaviour, 41, 997-1005. [ Links ]
Laland, K. N. & Plotkin, H. C. (1993). Social transmission of food preferences among Norway rats by marking of food sites and by gustatory contact. Animal Learning and Behavior, 21, 35-42. [ Links ]
Lashley, K. S. (1938). Conditional reactions in the rat. Journal of Psychology, 6, 311-324. [ Links ]
LoLordo, V. M. & Droungas, A. (1989). Selective associations and adaptive specializations: Taste aversions and phobias. En S. B. Klein & R. R. Mowrer (Eds.), Contemporary learning theories: Instrumental conditioning theory and the impact of biological constraints on learning (pp. 145-179). Hillsdale, NJ: Lawrence Erlbaum. [ Links ]
Miklósi, A. (1999a). The ethological analysis of imitation. Biological Reviews, 74, 347-374. [ Links ]
Miklósi, A. (1999b). From grasping to speech: Imitation might provide the missing link. Trends in Neurosciences, 22, 151-152. [ Links ]
Miller, N. E. & Dollard, J. (1941). Social learning and imitation. New Haven: Yale University Press. [ Links ]
Moore, B. R. (1996). The evolution of imitative learning. En C. M. Heyes & B. G. Galef, Jr. (Eds.), Social learning in animals: The roots of culture (pp. 221-243). San Diego, CA: Academic Press. [ Links ]
Nieto, J. & Cabrera, R. (1994). La evolución cultural en animales. En J. L. Díaz (Ed.), La mente y el comportamiento animal: Ensayos en etología cognitiva (pp. 91-106). México: Fondo de Cultura Económica. [ Links ]
Nieto, J., Cabrera, R., Guerra, J. & Posadas-Andrews, A. (1987). Tradiciones alimenticias: difusión de estrategias alimenticias novedosas en grupos animales. Revista Mexicana de Análisis de la Conducta, 13, 105-125. [ Links ]
Pellegrino, G. di, Fadiga, L., Fogassi, L., Gallese, V. & Rizzolatti, G. (1992). Understanding motor events: A neurophysiological study. Experimental Brain Research, 91, 176-180. [ Links ]
Rizzolatti, G. & Arbib, M. A. (1998). Language within our grasp. Trends in Neurosciences, 21, 188-194. [ Links ]
Rizzolatti, G. & Arbib, M. A. (1999). Reply. Trends in Neurosciences, 22, 152. [ Links ]
Rodas, R. (1974). Variables que influyen en la imitación voluntaria y en el aprendizaje imitativo. Revista Latinoamericana de Psicología, 6, 363-379. [ Links ]
Ross, R. T. & Holland, P. C. (1981). Conditioning of simultaneous and serial feature-positive discriminations. Animal Learning and Behavior, 9, 293-303. [ Links ]
Rusiniak, K. W., Hankins, W. G., García, J. & Brett, L. (1979). Flavor-illness aversions: I. Potentiation of odor by taste in rats. Behavioral and Neural Biology, 33, 74-92. [ Links ]
Schmajuk, N. A., Lamoreux, J. A. & Holland, P. C. (1998). Occasion setting: A neural network approach. Psychological Review, 105, 3-32. [ Links ]
Skinner, B. F. (1938). The behavior of organisms: An experimental analysis. Englewood Cliffs, NJ: Prentice Hall. [ Links ]
Skinner, B. F. (1953). Science and human behavior. Nueva York: Macmillan. [ Links ]
Spence, K. W. (1937). Experimental studies of learning and higher mental processes in infra-human primates. Psychological Bulletin, 34, 806-850. [ Links ]
Staddon, J. E. R. & Simmelhag, V. L. (1971). The “superstition” experiment: A reexamination of its implications for the principles of adaptive behavior. Psychological Review, 78, 3-43.
Suboski, M. D. & Bartashunas, C. (1984). Mechanisms for social transmission of pecking preferences to neonatal chicks. Journal of Experimental Psychology: Animal Behavior Processes, 10, 182-194. [ Links ]
Swartzentruber, D. (1995). Modulatory mechanisms of Pavlovian conditioning. Animal Learning and Behavior, 23, 123-143. [ Links ]
Thiele, T. E. & Frieman, J. (1994). Taste-potentiated color aversions in pigeons: Examination of CS preexposure and subsequent effects on postconditioning extinction of the taste aversion. Animal Learning and Behavior, 22, 436-441. [ Links ]
Thorpe, W. H. (1956). Learning and instinct in animals. Londres: Methuen. [ Links ]
Thorpe, W. H. (1963). Learning and instinct in animals. (2ª Ed.). Londres: Methuen. [ Links ]
van Kampen, H. S. (1996). A framework for the study of filial imprinting and the development of attachment. Psychonomic Bulletin and Review, 3, 3-20. [ Links ]
Whiten, A. & Ham, R. (1992). On the nature and evolution of imitation in the animal kingdom: Reappraisal of a century of research. Advances in the Study of Behavior, 21, 239-283. [ Links ]
Zajonc, R. B. (1965). Social facilitation. Science, 149, 269-274. [ Links ]
Zajonc, R. B. (1969). Coaction. En R. B. Zajonc (Ed.), Animal social psychology (pp. 9-12). Nueva York: Wiley. [ Links ]
Zentall, T. R. (1996). An analysis of imitative learning in animals. En C. M. Heyes & B. G. Galef, Jr. (Eds.), Social learning in animals: The roots of culture (pp. 221-243). San Diego, CA: Academic Press. [ Links ]
Zentall, T. R. & Hogan, D. E. (1976). Imitation and social facilitation in the pigeon. Animal Learning and Behavior, 4, 427-430. [ Links ]
Zentall, T. R., Sutton, J. E. & Sherburne, L. M. (1996). True imitative learning in pigeons. Psychological Science, 7, 343-346. [ Links ]
Recepción: mayo de 2004
Aceptación final:junio de 2005
* El autor desea agradecer al CONACYT el apoyo con el proyecto Núm. 3665P-H9608, 1997, PADEP Núm. 019306, 1996, y al PAPCA 2000 y 2003 por apoyo parcial del trabajo.
** Correspondencia: J. C. PEDRO ARRIAGA R. E-mail: jcpedro@servidor.unam.mx
