










Serviços Personalizados
artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista Latinoamericana de Psicología
versão impressa ISSN 0120-0534
Rev. Latinoam. Psicol. v.39 n.1 Bogotá abr. 2007
ARTÍCULOS
Bases fisiológicas del aprendizaje asociativo en mamíferos1
Physiological bases of associative learning in mammals
Agnès Gruart; José M. Delgado-García2
Universidad Pablo de Olavide, Sevilla, España
ABSTRACT
Classical conditioning of the corneal reflex is an useful model for the study of learning and memory processes. Our research group has studied the kinematics of reflex, voluntary, emotional and learned eyelid responses, as well as the physiology of facial motoneurons innervating the orbicularis oculi muscle; i.e., the muscle responsible for those eyelid movements. Facial motoneurons encode eyelid velocity during reflex lid responses, but eyelid position during learned lid responses. This functional difference reveals a selective processing (and/or origin) of both motor commands. Both the paravermal cerebelar cortex and the posterior interpositus nucleus are related to reflex and acquired eyelid responses, contributing to its proper performance, but not to its acquisition. At the same time, the hippocampus participates in the determination of the predictive value of the conditioned stimulus, when using classical conditioning procedures.
Keywords: Cerebellum, Hippocampus, Classic Conditioning, Eyelid, Mammals.
RESUMEN
El condicionamiento clásico del reflejo corneal es un modelo muy utilizado en el estudio de los procesos de aprendizaje y memoria. Nuestro grupo de investigación ha estudiado las características cinéticas de las respuestas palpebrales reflejas, voluntarias, emocionales y aprendidas, así como la fisiología de las motoneuronas que inervan el músculo orbicularis oculi, encargado de realizar dichos movimientos. Las motoneuronas faciales codifican la velocidad de cierre de los párpados durante respuestas reflejas y su posición durante respuestas aprendidas. Esta diferencia indica un diferente procesamiento (y/u origen) de ambas órdenes motoras. Tanto la corteza paravermal como el núcleo interpósito posterior del cerebelo están relacionados con los movimientos palpebrales reflejos y aprendidos, contribuyendo a su correcta realización, pero no a su aprendizaje. A su vez, el hipocampo interviene en la determinación del valor predictivo del estímulo condicionado, durante condicionamientos de tipo pavloviano.
Palabras clave: Cerebelo, Hipocampo, Condicionamiento clásico, Párpado, Mamíferos.
INTRODUCCIÓN
El presente texto contiene los resultados obtenidos en nuestro laboratorio a lo largo de 20 años sobre las bases fisiológicas del aprendizaje y la memoria, a los que se ha añadido los últimos hallazgos experimentales obtenidos por nuestro grupo, sobre todo en relación con la contribución del cerebelo y del hipocampo a la adquisición de nuevas habilidades motoras y cognitivas (Jiménez-Díaz et al., 2004; Leal-Campanario et al., 2005) y al desarrollo de modelos de condicionamiento clásico en ratones silvestres y transgénicos (Gruart et al., 2005). Hemos tratado de hacer aquí una presentación asequible y atractiva de un tema de investigación bastante árido a menudo. En su caso, una descripción más detallada desde el punto de vista cuantitativo, y contrastada con la bibliografía disponible, se puede encontrar en varios artículos publicados por nuestro grupo de investigación, a los cuales remitimos al lector interesado (Delgado-García et al., 1990; Delgado-García y Gruart, 2002; Gruart et al., 1994, 1995, 1997, 2000a, 2000b, 2000c, Gruart y Delgado-García, 1994; Magariños-Ascone et al., 1999, Múnera et al., 2001, Trigo et al., 1999a, 1999b). Existen también numerosas revisiones globales sobre el mismo tema realizadas por otros autores (Evinger et al., 1991; Gormezano et al., 1962, 1983; Thompson, 2005; Thompson y Krupa, 1994; Woody, 1986).
En primer lugar, nos ocuparemos de la descripción del sistema motor del párpado como un modelo experimental usado con frecuencia en el estudio de los mecanismos neuronales que subyacen al aprendizaje motor y cognitivo en los mamíferos. De hecho, el condicionamiento clásico de la respuesta palpebral (o de la membrana nictitante, en aquellas especies en que esto es posible desde el punto de vista técnico) se viene utilizando desde hace más de 50 años como ejemplo de generación de nuevas habilidades motoras mediante aprendizajes de tipo asociativo (Gormezano et al., 1962, 1983; Thompson, 2005; Thompson y Krupa, 1994). Seguidamente, se presentará la metodología que se sigue en este tipo de estudios, una descripción de la mecánica palpebral y de la fisiología de las motoneuronas que inervan el músculo orbicularis oculi y, finalmente, la contribución del cerebelo y del hipocampo a este tipo de aprendizaje motor. En tiempos recientes, ha crecido el apoyo a argumentos que consideran los procesos de aprendizaje motor desde el exclusivo punto de vista de la plasticidad neuronal, con especial referencia a los "sitios" (lugares del sistema nervioso central) donde aquéllos tienen lugar (Thompson y Krupa, 1994), así como de sus substratos moleculares (Bliss y Collindrige, 1993; Bracha y Bloedel, 1996; Edwards, 1995; Hawkins et al., 1983; Inda et al., 2005; Ito, 1989; Malenka, 1995). Por el contrario, se ha prestado una menor atención a los aspectos funcionales que tienen lugar in vivo, durante el momento mismo del aprendizaje (Delgado-García y Gruart, 2002). Además, hemos podido mostrar en un trabajo reciente que conviene separar de forma experimental aquellos fenómenos que corresponden al proceso de aprendizaje per se y aquellos otros que corresponden a la expresión motora o verbal de lo aprendido, evitando así errores importantes en la ubicación de las estructuras cerebrales que elaboran las respuestas aprendidas (Jiménez-Díaz et al., 2004). A estos aspectos funcionales se dedicará una atención especial en esta breve revisión sobre el condicionamiento clásico de la respuesta palpebral en mamíferos.
EL SISTEMA MOTOR PALPEBRAL COMO MODELO PARA EL ESTUDIO DE LOS MECANISMOS NEURONALES QUE SUBYACEN AL APRENDIZAJE MOTOR Y COGNITIVO
En la Figura 1 se ilustra el modelo experimental utilizado en este tipo de estudios. Al animal se le implantan de forma crónica (hasta 3-4 meses) electrodos bipolares para el registro de la actividad electromiográfica del músculo orbicularis oculi (orbicular de los párpados) y una pequeña bobina metálica en el párpado superior, para el registro de la posición palpebral por el método del seguidor magnético de la posición. El seguidor magnético de la posición es un instrumento que genera un campo magnético, de frecuencia y energía controladas, en dos planos del espacio: horizontal y vertical. Este instrumento es inocuo para el animal experimental y, de hecho, el seguidor magnético de la posición se utiliza en la clínica humana para estudios de la motórica ocular y palpebral. El animal experimental se coloca en el centro del campo magnético de modo que, cuando el párpado se mueve, la bobina implantada en el mismo se mueve también e induce un voltaje proporcional al ángulo y plano de desplazamiento (Becker y Fuchs, 1988; Gruart et al., 1995).
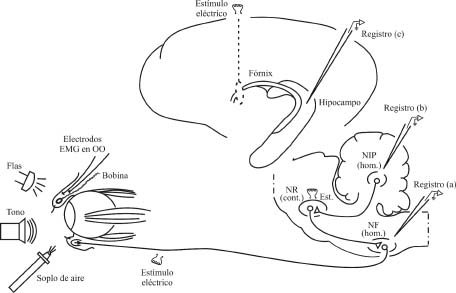
Figura 1. Modelo experimental para el estudio de la actividad cerebral durante el condicionamiento clásico del reflejo corneal. Al animal se le implanta un electrodo bipolar para el registro de la actividad electromiográfica (EMG) del músculo orbicularis oculi (OO). Se le implanta también una pequeña bobina metálica en el párpado superior, para medir los desplazamientos del mismo con la técnica del seguidor magnético de la posición. Para ello, el animal se coloca dentro de un campo magnético y el desplazamiento de la bobina metálica dentro de dicho campo induce un voltaje proporcional al ángulo de desplazamiento (Gruart et al., 1995, 2000a). De forma adicional, se pueden implantar otras dos bobinas alrededor del globo ocular para medir los desplazamientos rotacionales y translacionales del mismo (Delgado-García et al., 1990). Como estímulos se pueden utilizar soplos de aire (20-100 ms, 13 Kg/cm2) aplicados a la córnea, sonidos (20-350 ms, 600-2.400 Hercios, 90 dB), o flases de luz (1 ms). La actividad eléctrica neuronal se registra con micropipetas de vidrio rellenas de una solución conductora (NaCl). Las neuronas registradas se identifican in vivo mediante su activación antidrómica desde sus sitios de proyección. Se indican algunos puntos de registro, como el núcleo facial (a), el núcleo interpósito posterior homolateral (b), o el hipocampo (c). Abreviaturas: Est., estimulación; NF, núcleo motor facial; NIP, núcleo interpósito posterior del cerebelo; NR, núcleo rojo; cont., hom., contra- y homolateral. Figura modificada con permiso de Delgado-García (2003).
El animal experimental se prepara también para el registro de la actividad eléctrica unitaria neuronal en estructuras seleccionadas, como el núcleo motor facial (a, en Figura 1A), el núcleo interpósito del cerebelo (b, en Figura 1A), o las células piramidales del hipocampo (c, en Figura 1A). El registro se realiza con pipetas de vidrio de alta resistencia (35 Mohmios). Los registros de potenciales extracelulares de campo se suelen llevar a cabo con electrodos metálicos (acero inoxidable o tungsteno) de pequeño diámetro (50 mm), implantados de forma crónica, sobre todo si los experimentos se realizan en mamíferos de pequeño tamaño (rata, ratón). Las neuronas registradas se identifican in vivo mediante su activación antidrómica desde sus sitios de proyección. Por ejemplo, las motoneuronas faciales se activan antidrómicamente desde el nervio facial, las neuronas del núcleo interpósito del cerebelo desde el núcleo rojo y las células piramidales de CA3 del hipocampo desde el fórnix (Gruart et al., 2000a; Múnera et al., 2001; Trigo et al., 1999a).
Como prueba de aprendizaje asociativo se suele utilizar el condicionamiento clásico del reflejo corneal, con paradigmas tanto de traza como de demora (Figura 2). En el paradigma de traza se presenta un breve (20 ms) soplo de aire en la córnea izquierda (1 Kg/cm2) o un tono (600-2400 Hercios, 90 dB) como estímulo condicionado. Unos 250-500 ms después de terminado el estímulo condicionado se presenta un intenso (100 ms, 3 Kg/cm2) soplo de aire en la misma córnea, como estímulo incondicionado. Por el contrario, en el paradigma de demora se presenta un sonido (600-2400 Hercios, 90 dB) como estímulo condicionado, el cual se mantiene por unos 350 ms. A los 250 ms de iniciado el sonido se presenta un soplo intenso en la córnea izquierda (100 ms, 3 Kg/cm2) como estímulo incondicionado, que termina simultáneamente con el estímulo condicionado. La frecuencia del sonido utilizado depende de la especie que se utiliza: a menor tamaño mayor frecuencia. También hay que indicar que, en algunos casos, como por ejemplo en especies difíciles de inmovilizar (rata, ratón) los soplos de aire se sustituyen por estímulos eléctricos (pulsos cuadrados bifásicos de 50-500 ms) de intensidad variable (< 1 mA, 13 × Umbral). En cada sesión de condicionamiento se presentan las parejas de estímulos condicionado e incondicionado entre 60 y 120 veces, según la especie animal a condicionar, separadas en bloques de 10 parejas. Se pueden encontrar más detalles de este modelo experimental en trabajos más especializados de nuestro grupo (Gruart et al., 1995; 2000c).
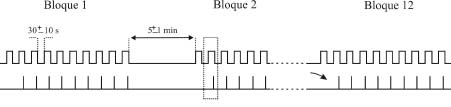
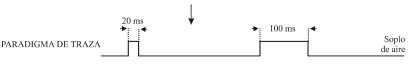
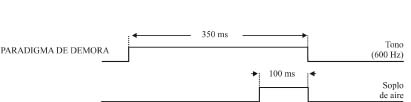
Figura 2. Paradigmas de condicionamiento clásico o pavloviano. Se ilustra el diseño experimental para condicionamientos de traza y de demora. Los estímulos condicionado e incondicionado se presentan emparejados en bloques de 10, con un intervalo variable entre parejas de unos 30 s. Se indica las duraciones más habituales de los estímulos, las cuales pueden variar, según el diseño experimental. Cada bloque (1, 2,..., 12) está separado del anterior unos 5 minutos. Normalmente, una sesión de condicionamiento dura unas dos horas para gatos, una hora para conejos y 30 min para ratas y ratones.
Ver texto para más detalles. Figura modificada con permiso de Delgado-García (2003).
CARACTERÍSTICAS CINÉTICAS Y FUNCIONALES DEL SISTEMA MOTOR PALPEBRAL
El sistema motor facial es peculiar por diversas razones. Desde el punto de vista evolutivo, este sistema motor es de aparición tardía, desarrollándose principalmente en felinos y primates. Un dato importante, olvidado con frecuencia, es que el sistema motor facial es un conjunto muscular de origen visceral modificado, lo que explica en cierto modo su fácil entrenamiento con el condicionamiento clásico o pavloviano, más fácil de aplicar en el sistema vegetativo autónomo (secreción salivar, por ejemplo) que en el sistema motor estriado (contracción de una extremidad). En el hombre, la musculatura facial está compuesta por más de 20 músculos diferentes. Una característica común a casi todos los músculos faciales es su pequeña masa, por lo que se afectan escasamente por la fuerza de la gravedad. Los músculos faciales, y entre ellos los que mueven el párpado, se denominan de carga constante, porque no están sometidos a cambios en el peso que deben movilizar cuando se contraen; en esto se diferencian de la musculatura espinal y de las extremidades, que son de carga variable; es decir, que están preparados para soportar pesos diferentes sin que ello impida su funcionamiento.
Gran parte de la musculatura facial (incluyendo la de los párpados) presenta una disposición plana, en mapa, ya que cada unidad motora ocupa un lugar preciso y exclusivo dentro del músculo. Esta disposición tiene una gran importancia funcional, pues cada motoneurona facial puede realizar una contribución específica y única a la mímica de la cara. De hecho, las motoneuronas faciales están distribuidas en subnúcleos dentro del núcleo o complejo motor facial. Incluso, las motoneuronas faciales mantienen un alto grado de somatotopía dentro de cada subnúcleo (Morcuende et al., 2003; Trigo et al., 1999a). En este sentido, se sabe que a cada subnúcleo llegan aferencias específicas para cada pequeña agrupación de motoneuronas. Por ejemplo, las motoneuronas que inervan la porción palpebral (tarsal) del músculo orbicularis oculi se localizan en la región más dorsal del núcleo intermedio facial y se activan principalmente durante respuestas reflejas originadas por estímulos mecánicos de la córnea y de la piel de los párpados. Por el contrario, las motoneuronas que inervan la porción orbitaria (septal) del músculo orbicularis oculi se activan preferentemente durante expresiones faciales de relacionadas con el estado emocional interno del sujeto, por lo que sus aferencias son preferentemente de origen límbico (Gordon, 1951; Trigo et al., 1999a). Básicamente, el músculo orbicularis oculi descansa sobre los bordes del hueso de la órbita, llamada área orbicular y sobre una parte móvil, denominada área palpebral, que forma el párpado propiamente dicho. En la zona palpebral se distingue una división pretarsal, cercana a los bordes de los párpados superior e inferior, y circundando a ésta, una parte preseptal, que se continúa con la parte orbital (Lander et al., 1996). Las fibras que forman el borde de los párpados (pretarsales) son pequeñas y mayoritariamente fásicas, participando en contracciones rápidas del párpado. En las partes más alejadas de la porción pretarsal se encuentran las fibras tipo 1 (tónicas), más gruesas y lentas, y que alcanzan una proporción del 20 %. Este gradiente en tamaño y composición se conserva constante en las diferentes especies estudiadas (conejos, monos, seres humanos, etc.) (McLoon y Wirtschafter, 1991).
Con alguna excepción, los músculos faciales y, en concreto, los del párpado, carecen de propioceptores típicos. Desde el punto de vista funcional, esto se traduce en que no hay reflejo miotático directo, o de estiramiento, en la musculatura palpebral (ver referencias en Trigo et al., 1999b). Esta es probablemente la causa de que las motoneuronas que inervan el músculo orbicularis oculi carezcan de información relativa a la posición que ocupa el párpado sobre el globo ocular. La posición palpebral durante la vigilia la determina el músculo elevador de los párpados, ya que las motoneuronas que inervan este músculo tienen información de la posición del ojo en la órbita (mirada hacia arriba o abajo) como el resto de la musculatura extraocular que mueve el ojo en el plano vertical (Becker y Fuchs, 1998; Delgado-García, 2000). Durante los movimientos palpebrales rápidos que acompañan a las sacadas oculares, el músculo orbicularis oculi está completamente relajado. Este músculo se activa exclusivamente cuando los párpados se cierran de forma refleja (por estímulos de la piel, sonoros o visuales) y durante movimientos voluntarios, aprendidos, o de origen emocional. Llama la atención que la respuesta palpebral condicionada (aprendida mediante condicionamiento clásico) se parece más en su cinética a la respuesta palpebral de tipo emocional que a la respuesta de tipo reflejo o al movimiento palpebral voluntario (Gruart et al., 1995). Esto sugiere que los mecanismos neuronales que producen las respuestas palpebrales aprendidas pueden compartir determinados circuitos efectores con los que generan las respuestas palpebrales de origen emocional.
Los párpados se pueden cerrar de forma pasiva cuando se relaja el músculo elevador que los mantiene abiertos, incluso si nos colocamos cabeza abajo. Por tanto, el cierre pasivo de los párpados (por ejemplo, al dormir) no ocurre por acción de la gravedad, sino por la liberación de las tensiones acumuladas en los tendones y aponeurosis palpebrales, tras la relajación del músculo elevador de los párpados (Evinger et al., 1991; Trigo et al., 2003). Hay que tener en cuenta que el cierre activo de los párpados en los primates es el resultado exclusivo de la contracción del músculo orbicularis oculi, mientras su apertura depende de la acción del músculo elevador. Pero, en animales con membrana nictitante (la mayoría de vertebrados terrestres) existe un músculo que se inserta en el polo posterior del globo ocular, el músculo retractor bulbi, que retrae el ojo hacia el fondo de la órbita, para permitir el deslizamiento de dicha membrana por la superficie corneal. Esta retracción del globo ocular contribuye también al cierre de los párpados (Evinger et al., 1991; Gruart et al., 1995). El músculo retractor bulbi está formado por fibras grandes y de contracción rápida; cada unidad motora cuenta con unas 70 fibras y no tiene inervación múltiple (Lennerstand, 1975). Debido a estas características, el músculo retractor bulbi participa en movimientos rápidos como el parpadeo reflejo; así, su función básica es la de protección del ojo. Por el contrario, el músculo orbicularis oculi participa también en la expresión de las emociones (Bach-y-Rita, 1971) y en la generación de respuestas aprendidas (Trigo et al., 1999). Por último, algunas especies como el conejo y el gato son capaces de cocontraer los cuatro músculos rectos que se insertan en el globo ocular, sobre todo en respuesta a estímulos corneales muy intensos, lo que hace que el ojo se retraiga hacia el fondo de la órbita, contribuyendo al cierre de los párpados (Gruart et al., 2000c).
Por lo tanto, un movimiento tan simple en apariencia con el de cerrar los párpados es el resultado motor de la acción de muy diversos músculos, gobernados a su vez por diferentes centros motores troncoencefálicos. Asimismo, con los párpados se pueden realizar movimientos palpebrales muy diferentes (pasivos, reflejos, automáticos, voluntarios, aprendidos y de origen emocional), todos ellos de una extraordinaria velocidad, precisión y expresividad.
FISIOLOGÍA DE LAS MOTONEURONAS FACIALES QUE INERVAN EL MÚSCULO ORBICULARIS OCULI
Hemos registrado la actividad de las motoneuronas faciales (Figura 3A) que inervan al músculo orbicularis oculi en el gato despierto y alerta durante la realización de respuestas palpebrales reflejas y condicionadas (Trigo et al., 1999a), así como la de las motoneuronas que inervan el músculo retractor bulbi, localizadas en el núcleo accesorio del motor ocular externo. La identificación de las motoneuronas faciales se lleva a cabo mediante la activación antidrómica de las neuronas registradas al aplicar pulsos de corriente en los electrodos implantados en la rama palpebral del nervio facial, o VII par craneal (Figura 3A). Las motoneuronas de los músculos orbicularis oculi y retractor bulbi generan dos brotes de potenciales de acción (a los 46 y 1016 ms) en respuesta a la presentación de un soplo de aire intenso a la córnea homolateral (Figura 3B). En el gato, las motoneuronas faciales, pero no las del orbicularis oculi, responden a la presentación de tonos o de flases de luz. Mediante análisis lineales de la regresión se ha comprobado que las motoneuronas faciales codifican la velocidad con que el párpado se cierra durante el reflejo corneal, pero no la posición del párpado sobre el globo ocular (Trigo et al., 1999a). De hecho, y como ya se ha indicado más arriba, la posición del párpado sobre el ojo se determina por la actividad de las motoneuronas que inervan el músculo elevador de los párpados (Becker y Fuchs, 1988).
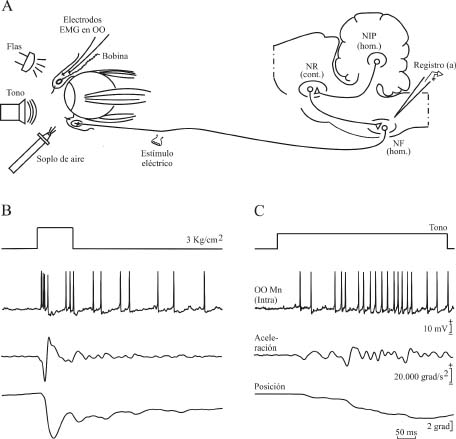
Figura 3. Diseño experimental (A) y actividad de las motoneuronas faciales que inervan el músculo orbicularis oculi durante respuestas palpebrales reflejas (B) y condicionadas (C). En A, se muestra el diseño experimental para el registro en el núcleo facial de motoneuronas del músculo orbicularis oculi. En B y C, y de arriba abajo, se muestran el estímulo aplicado, el registro intracelular de una motoneurona facial y la aceleración (en grados/s2) y posición (en grados) del párpado. El desplazamiento hacia abajo del trazado de la posición del párpado indica su cierre. En B, se aplica un soplo de aire durante 100 ms a la córnea homolateral al registro, para inducir el reflejo corneal. En C, se muestra un ejemplo de respuesta condicionada evocada por la sola presentación del estímulo condicionado (un tono de 600 Hercios y 90 dB). La calibración en C es también para B. Ver texto para más detalles. Abreviaturas: cont., contralateral; hom., homolateral; EMG, electromiografía; OO, músculo orbicularis oculi; NF, núcleo facial; NIP, núcleo interpósito posterior; NR, núcleo rojo. Datos tomados y modificados de Trigo et al. (1999a). Figura modificada con permiso de Delgado-García (2003).
Las motoneuronas faciales funcionan de un modo muy distinto al descrito durante la realización de respuestas palpebrales aprendidas (Figura 3C). En el gato, las motoneuronas que inervan el músculo retractor bulbi no se activan durante las respuestas palpebrales condicionadas (Trigo et al., 1999a), aunque se asume que sí lo hacen en el conejo. Esto puede deberse a que la respuesta aprendida tiene una velocidad pico que no llega al 10% de la que se alcanza durante las respuestas palpebrales reflejas, lo que sugiere que la señal de activación que llega a las motoneuronas del músculo retractor bulbi durante el condicionamiento no es suficientemente intensa para despolarizarlas. Por el contrario, las motoneuronas faciales que inervan el músculo orbicularis oculi sí se despolarizan lo suficiente para producir potenciales de acción ante la presentación de los estímulos condicionado e incondicionado. De todas formas, la activación de estas motoneuronas durante el condicionamiento clásico del reflejo corneal es más débil que la que presentan durante la realización de respuestas palpebrales reflejas. Es importante señalar que las motoneuronas faciales no codifican la velocidad del desplazamiento del párpado durante las respuestas condicionadas, sino su posición. De hecho, en la Figura 3 se puede observar el disparo preferentemente fásico (o en brotes) que tienen estas motoneuronas durante las respuestas palpebrales reflejas (Figura 3B) y preferentemente tónico durante las respuestas palpebrales aprendidas (Figura 3C). Estos datos sugieren que las respuestas reflejas se elaboran en el soma de las motoneuronas, por lo que producen una gran despolarización de la misma, mientras que las respuestas aprendidas se elaboran en las dendritas, causando una despolarización sostenida, pero menos intensa (Trigo et al., 1999a). Recientemente, se ha descrito que determinados aferentes colinérgicos a las motoneuronas faciales pueden intervenir en la generación del disparo tónico que caracteriza la producción de respuestas palpebrales aprendidas (Magariños-Ascone et al., 1999), de un modo probablemente similar al descrito por nuestro grupo para la generación de la señal de posición ocular tras la realización de un movimiento sacádico del ojo (ver Navarro-López et al., 2004).
¿DÓNDE Y CÓMO SE GENERAN LAS RESPUESTAS PALPEBRALES APRENDIDAS?
Como consecuencia de lo expuesto anteriormente, surge la cuestión acerca del origen de las señales neuronales que generan las respuestas motoras condicionadas, o aprendidas. En un trabajo realizado en colaboración con la Dr. Ugolini del CNRS, GyfsurYvette, Francia (Morcuende et al., 2002), hemos identificado el complejo sistema premotor que proyecta sobre las motoneuronas que inervan el músculo orbicularis oculi. Este estudio se realizó usando el virus atenuado de la rabia como marcador transináptico retrógrado. Para ello, se inyectó una pequeñísima cantidad de este virus en el párpado superior de ratas y, con un anticuerpo específico, se revelaron las proyecciones desde el músculo facial. El estudio se realizó en distintos periodos tras las inyección del virus, para conocer cuáles eran las conexiones directas o motoras (es decir de las motoneuronas faciales) y las premotoras (es decir, las correspondientes a las neuronas que proyectan sobre las motoneuronas). Así, se fueron localizando las estructuras cerebrales a las que llega el virus mediante transporte transináptico retrógrado al cabo de un día, de dos, de tres, etc. De dicho estudio se concluye que hay numerosos centros troncoencefálicos, cerebelosos, y de la corteza cerebral, relacionados más o menos directamente con la elaboración de las diferentes órdenes motoras que determinan la producción de movimientos palpebrales reflejos, voluntarios, emocionales y aprendidos. A continuación se presentan los estudios electrofisiológicos realizados en algunas de las estructuras cerebrales identificadas por su carácter pre o prepremotor; es decir, que proyectan mono, bi o polisinápticamente sobre las motoneuronas del músculo orbicular de los párpados.
Existen dos estructuras que se han propuesto con gran énfasis como el (posible) sitio o sitios dónde ocurren los procesos moleculares que subyacen al aprendizaje motor; éstas son el hipocampo y el cerebelo (Bracha y Bloedel, 1996; Clack y Squire, 1998; Gruart et al., 2000a, Jiménez-Díaz et al., 2004; Múnera et al., 2001; Thompson, 2005; Thompson y Krupa, 1994; Woody, 1986). Concretamente, se ha sugerido que en el hipocampo ocurren procesos funcionales y estructurales relacionados con la potenciación a largo plazo (o LTP, siglas en inglés de longterm potentiation), mientras que en el cerebelo se tienen lugar mecanismos de inhibición a largo plazo (o LTD, siglas en inglés para long-term depression). Hasta el momento, la mayoría de estudios realizados sobre ambos fenómenos lo han sido en preparaciones in vitro (Bliss y Collingridge, 1993; Edwards, 1995; Hawkins et al., 1983; Ito, 1989; Malenka, 1995; Woody, 1986). A la vista de esta situación, nuestro grupo ha prestado un interés especial al estudio de la actividad neuronal que se registra en dichas estructuras en el animal en condiciones fisiológicas y durante el proceso mismo de la adquisición de las respuestas palpebrales condicionadas.
PARTICIPACIÓN DEL CEREBELO EN EL CONDICIONAMIENTO CLÁSICO DEL REFLEJO CORNEAL
En un estudio inicial (Gruart y Delgado-García, 1994) describimos que en los núcleos profundos del cerebelo se encuentran neuronas relacionadas de un modo u otro con los movimientos reflejos del párpado, producidos por estímulos trigeminales, acústicos y visuales. En efecto, el marcaje de los núcleos cerebelosos tras la inyección del virus atenuado de la rabia en el párpado superior indica una localización difusa de neuronas nucleares relacionadas con la motórica palpebral, principalmente en la región lateral del núcleo interpósito posterior y en la cara medial del núcleo dentado (Morcuende et al., 2002). Así pues, existe una zona nuclear del cerebelo donde se concentra un importante cúmulo de neuronas relacionadas con los movimientos palpebrales. Esta zona se ubica, tanto en la rata como en el gato, en el polo rostral del núcleo interpósito posterior, en inmediata vecindad con el núcleo dentado (Gruart et al., 1994, 2000a; Gruart y Delgado-García, 1994; Morcuende et al., 2002) y se pueden identificar durante el registro electrofisiológico mediante la activación antidrómica desde el núcleo rojo (Figura 4A).
Las neuronas del polo rostral del interpósito posterior presentan una actividad eléctrica claramente relacionada con respuestas palpebrales reflejas y condicionadas. De acuerdo con los resultados obtenidos en diversos estudios (Delgado-García y Gruart, 2002; Gruart et al., 2000a; Gruart y Delgado-García, 1994; Jiménez-Díaz et al., 2004), dichas neuronas se pueden clasificar en dos tipos (A y B) en función de la actividad eléctrica que presentan durante el movimiento palpebral. Las neuronas tipo A se activan unos 20 ms después del inicio del movimiento palpebral (ver Figura 4B) y el perfil de su frecuencia instantánea de disparo semeja a veces el perfil del movimiento palpebral. Sin embargo, no se ha podido demostrar que la frecuencia instantánea de disparo de estas neuronas tipo A esté linealmente relacionado con la posición, velocidad o aceleración del párpado durante movimientos reflejos, voluntarios o aprendidos. Las neuronas tipo A tienen una frecuencia media de disparo de 1060 potenciales de acción/s en el gato despierto y alerta. Esta frecuencia se incrementa considerablemente (hasta los 300 potenciales de acción/s) durante los movimientos palpebrales reflejos y aprendidos (Figura 4B). Por otra parte, las neuronas tipo B, que se localizan entremezcladas con las neuronas tipo A en el mismo polo rostral del interpósito posterior se inhiben durante el movimiento palpebral, ya sea de origen reflejo o condicionado (ver Figura 4C). Las neuronas tipo B tienen una frecuencia espontánea de disparo de 3080 potenciales de acción/s, frecuencia que disminuye hasta prácticamente desaparecer durante el movimiento palpebral. En este caso también, la inhibición en el disparo neuronal se inicia unos milisegundos después que el movimiento palpebral.
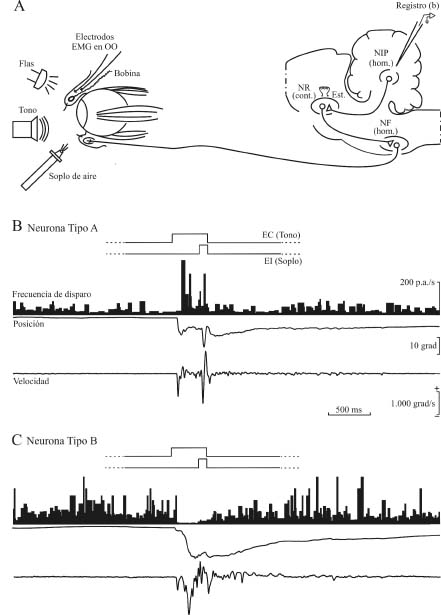
Figura 4. Diseño experimental y actividad de dos neuronas de tipos diferentes localizadas en el núcleo interpósito posterior del cerebelo durante la realización de movimientos palpebrales condicionados. En A se muestra el diseño experimental para el registro de neuronas en el núcleo interpósito posterior (NIP) homolateral (hom.) al párpado al que se aplica el estímulo incondicionado. En B se muestra la actividad eléctrica de una neurona tipo A, mientras que en C se muestra la actividad de una neurona tipo B. En ambos casos, y de arriba abajo, se ilustran el paradigma de condicionamiento (demora), la frecuencia de disparo (en potenciales de acción/s), así como la posición (en grados) y la velocidad (en grados/s) del párpado. El desplazamiento hacia abajo del trazado de la posición del párpado indica su cierre. Las calibraciones incluidas en B son también para C. Abreviaturas: cont., contralateral; EMG, electromiografía; OO, músculo orbicularis oculi; NF, núcleo facial; NR, núcleo rojo. Datos tomados y modificados de Gruart et al. (2000a). Figura modificada con permiso de Delgado-García (2003)
Los cambios en frecuencia (activación o inhibición) que se observan durante el registro electrofisiológico de las neuronas tipo A y B del núcleo interpósito posterior del cerebelo no tienen lugar al inicio de las sesiones de condicionamiento, sino que ocurren con una pendiente muy poco pronunciada (del orden de 0,05 potenciales de acción/s/ensayo). Esta pendiente indica que dichas neuronas aumentan (las tipo A) o disminuyen (las tipo B) sus frecuencias medias de disparo de potenciales de acción alrededor de 5 potenciales de acción/s cada 100 ensayos; es decir, cada sesión de entrenamiento. Además, es importante resaltar que este incremento en actividad sólo es detectable en el intervalo entre la presentación del estímulo condicionado y el inicio del estímulo incondicionado; es decir, durante la realización de la respuesta palpebral aprendida. En cualquier caso, la actividad de estas neuronas está ligada a la aparición de la respuesta condicionada y no a la mera presentación de la pareja de estímulos condicionado e incondicionado. Este último punto indica que el núcleo interpósito posterior del cerebelo está claramente relacionado con la realización del movimiento palpebral aprendido, pero no con su iniciación o génesis, ya que la activación (tipo A) o inhibición (tipo B) no precede, sino que sigue al inicio de la respuesta condicionada (Delgado-García y Gruart, 2002; Jiménez-Díaz et al., 2004).
Recientemente, hemos publicado un experimento que pone en evidencia el importante papel del núcleo interpósito posterior del cerebelo en la realización de la respuesta palpebral aprendida o condicionada (Jiménez-Díaz et al., 2004). El experimento consiste en la microestimulación de la zona habitual de registro, ubicada en el polo rostral del núcleo interpósito posterior. Esta micro-estimulación produce una respuesta palpebral de perfil cinético similar al de la respuesta condicionada, sobre todo si se acompaña de la presentación de un estímulo condicionado (ver Figura 5 en Jiménez-Díaz et al., 2004). Por el contrario, la microinyección de muscimol (un conocido agonista del receptor GABAA) en la misma zona produce una disminución en la amplitud de la respuesta palpebral condicionada (ver Figura 3 en Jiménez-Díaz et al., 2004). En este último caso, la inhibición de las neuronas del núcleo interpósito por la aplicación local de muscimol disminuye su efecto facilitador (a través del núcleo rojo) sobre la activación de las motoneuronas que inervan el músculo orbicularis oculi. El resultado motor es la elaboración de una respuesta palpebral aprendida de menor amplitud de lo normal.
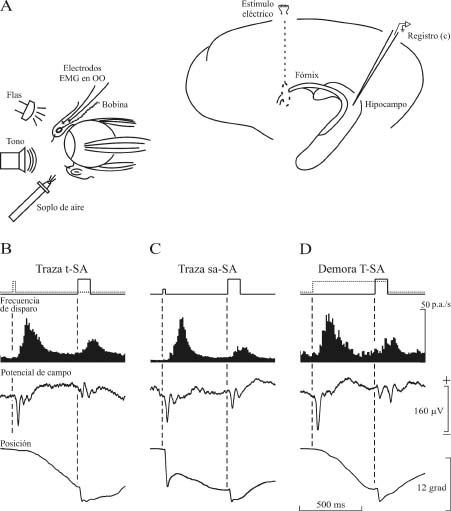
Figura 5. Actividad de las células piramidales del hipocampo durante distintos paradigmas de condicionamiento clásico. En A se muestra el diseño experimental para el registro de la actividad eléctrica de neuronas piramidales de CA3 y CA1 que proyectan sus axones a través del fórnix. En B se muestra la actividad eléctrica de una neurona piramidal de la capa CA3 durante un condicionamiento clásico del reflejo corneal con un paradigma de traza. Como estímulo condicionado (EC) se utiliza un breve tono (20 ms, 600 Hercios, 90 dB). En C se utiliza como EC un débil soplo (20 ms, 1 Kg / cm2), mientras que en D el EC es un tono duradero (350 ms, 600 Hercios, 90 dB). En los tres casos, el estímulo incondicionado consiste en un soplo de aire largo e intenso (100 ms, 3 Kg / cm2). De arriba a abajo se presentan el paradigma experimental, la frecuencia de disparo de la neurona (en potenciales de acción/s), el potencial de campo (en mV) evocado por los estímulos condicionado e incondicionado y la posición (en grados) del párpado. El desplazamiento hacia abajo del trazado de la posición del párpado indica su cierre. Nótese que los tres paradigmas de condicionamiento producen un cambio similar de la actividad neuronal, pero un movimiento palpebral claramente distinto. Las calibraciones en D son también para B-C. Abreviaturas: EMG, electromiografía; OO, músculo orbicularis oculi. Datos tomados y modificados de Múnera et al. (2001). Figura modificada con permiso de Delgado-García (2003).
Así pues, el núcleo interpósito posterior del cerebelo contribuye a la despolarización de las motoneuronas faciales durante el intervalo de tiempo entre el inicio del estímulo condicionado y del incondicionado, momento en que aparece la respuesta palpebral condicionada. Este efecto se ejerce preferentemente por las neuronas tipo A, las cuales proyectan a la región dorso-medial del núcleo rojo, mientras que las neuronas rubrales proyectan monosinápticamente sobre las motoneuronas del músculo orbicularis oculi (Morcuende et al., 2002). A su vez, es probable que las neuronas tipo B proyecten sobre interneuronas localizadas en las proximidades del núcleo del tercer par craneal y que contribuyan a inhibir (o desfacilitar) la actividad de las motoneuronas del músculo elevador de los párpados, favoreciendo así el cierre de los mismos durante la respuesta condicionada (Gruart y Delgado-García, 1994). De acuerdo con estos datos experimentales, el cerebelo no parece funcionar como un órgano coordinador, integrador y computador del movimiento aprendido (Thompson y Krupa, 1994), sino más bien como un órgano reforzador (Delgado-García y Gruart, 2002; Jiménez-Díaz et al., 2004) de la orden motora procedente de alguna otra estructura cerebral, probablemente de la corteza motora y premotora (Aou et al., 1992; Gruart et al., 2000b; Leal-Campanario et al., 2005).
PARTICIPACIÓN DEL HIPOCAMPO EN EL CONDICIONAMIENTO CLÁSICO DEL REFLEJO CORNEAL
Nuestro grupo de investigación ha abordado también el estudio de la actividad eléctrica de las neuronas piramidales de las zonas CA3 y CA1 del hipocampo durante la realización de respuestas palpebrales reflejas y aprendidas (Múnera et al., 2001). Las neuronas registradas se activaron antidrómicamente desde el fórnix (Figura 5A). Las células piramidales registradas en el gato despierto y en condiciones fisiológicas producen un brote de pocos potenciales de acción unos 60-80 ms después de presentado un estímulo novedoso, como un sonido, o un breve soplo de aire aplicado a la córnea del lado contralateral al de registro (ver Figura 5). Esta respuesta eléctrica neuronal tiende a desaparecer con la presentación repetida del mismo estímulo. Sin embargo, cuando un estímulo condicionado (el mismo tono o soplo débil de aire) se empareja repetidamente con un estímulo incondicionado (soplo de aire fuerte, de unos 3 Kg/cm2 de presión, aplicado a la córnea), la respuesta neuronal al estímulo condicionado aumenta considerablemente a lo largo de las sesiones de condicionamiento, mientras que la respuesta al estímulo incondicionado disminuye o se mantiene igual (ver Figura 1 en Múnera et al., 2001). Este incremento en la frecuencia instantánea de disparo de las células piramidales tiene una pendiente que se asemeja (aproximadamente 0,05 potenciales de acción/s/ensayo) a la ya descrita para las neuronas del cerebelo, lo que sugiere que los procesos neuronales relacionados con este aprendizaje asociativo pueden estar ocurriendo a la vez en lugares del cerebro tan diversos como las motoneuronas faciales, el cerebelo y el hipocampo. De acuerdo con la hipótesis que hemos propuesto en una revisión reciente (Delgado-García y Gruart, 2002) estos cambios en la actividad neuronal corresponderían a un estado funcional particular que abarcaría numerosas estructuras cerebrales y que sería específico para este tipo de aprendizaje.
En repetidas ocasiones se ha propuesto que el hipocampo sólo estaría relacionado con la adquisición de condicionamiento clásico utilizando paradigmas de traza, pero no en los de demora, para los que se requeriría el cerebelo (Clark y Squire, 1998; Thompson, 2005; Thompson y Krupa, 1994). Esta afirmación se podría poner en duda de acuerdo a los resultados experimentales ilustrados en la Figura 5B. Dicha figura muestra que las neuronas piramidales del hipocampo en el gato despierto responden prácticamente del mismo modo con los paradigmas de traza y de demora. Se puede afirmar incluso que la respuesta de las células piramidales del hipocampo parece independiente de la modalidad sensorial del estímulo condicionado utilizado [soplo de aire débil (B), tono breve (C), o tono prolongado (D), ver Figura 5]. De acuerdo, pues, con estos resultados, el hipocampo parece estar más relacionado con la relevancia, saliencia o valor predictivo del estímulo condicionado (Múnera et al., 2001; Rescorla, 1988) que con la determinación del intervalo de tiempo entre los estímulos condicionado e incondicionado (Thompson y Krupa, 1994), o con el paradigma de traza en exclusiva (Clark y Squire, 1998).
Recientemente, hemos emprendido el estudio de la actividad sináptica individual a lo largo del circuito del hipocampo en ratones silvestres y transgénicos (Gruart et al., 2005). Para ello, el animal se implanta con electrodos de estimulación en la rama supraorbitaria del nervio del trigémino y con electrodos de estimulación en el músculo orbicularis oculi. Además, el animal se implanta con electrodos de estimulación en las colaterales de Schaffer en el hipocampo contralateral, así como con electrodos de registro en la zona de las dendritas apicales de la capa CA1 del hipocampo (stratum radiatum). Se realiza un condicionamiento de traza similar al descrito más arriba, con la diferencia de que a los 300 ms de la presentación del estímulo condicionado se da un breve (50 ms) pulso eléctrico en las colaterales de Schaffer y se registra el potencial monosináptico extracelular que se produce en CA1. Así se puede seguir la evolución funcional de esta sinapsis a lo largo del condicionamiento.
Los resultados preliminares disponibles indican que el potencial sináptico evocado en CA1 por estimulación de las colaterales de Schaffer incrementa ligeramente en su pendiente (mV/s) a lo largo del aprendizaje. Esto sugiere la presencia de una facilitación de dicho paso sináptico, debida al proceso de condicionamiento (Gruart et al., 2005). También hemos podido demostrar que la inducción artificial de potenciación a largo plazo (LTP) en dicha sinapsis es capaz de bloquear el aprendizaje del animal experimental. Esta técnica tiene un interés especial porque permite ser aplicada en ratones transgénicos, knock-out condicionales, mutantes espontáneos, etc. Esto supone la disponibilidad de técnicas de estudios conductuales y de aprendizaje in vivo en estos pequeños roedores.
LÍNEAS FUTURAS DE EXPERIMENTACIÓN
Por lo dicho hasta aquí, una pregunta aún pendiente de respuesta es dónde y cómo se genera o elabora la respuesta palpebral condicionada. Estudios electrofisiológicos en gatos (Aou et al., 1992) y conejos (Leal-Camapanario et al., 2005) y estudios recientes sobre genes de expresión temprana (Gruart et al., 2000b) sugieren que amplias zonas de la corteza motora y sensorial podrían estar relacionadas con el aprendizaje de este tipo de respuesta motora y/o con los aspectos cognitivos de la misma. Así pues, parece necesario estudiar en detalle estas zonas corticales durante el momento mismo de la adquisición del movimiento palpebral condicionado; es decir, realizando experimentos in vivo, con el animal en perfectas condiciones fisiológicas. Más arriba se ha indicado la importancia que tendría la realización de registros de la actividad neuronal de modo simultáneo en muy diversas regiones corticales y subcorticales relacionadas con el movimiento palpebral (Morcuende et al., 2002) durante el proceso de aprendizaje. El objetivo a medio plazo sería determinar no ya el sitio, sino el estado funcional subyacente al aprendizaje motor. A esta línea experimental están dedicados, pues, todos nuestros esfuerzos.
REFERENCIAS
Aou, S., Woody, C.D. & Birt, D. (1992). Changes in the activity of units of the cat motor cortex with rapid conditioning and extinction of a compound eye blink movement. Journal of Neuroscience, 12, 549-559. [ Links ]
Becker, W. & Fuchs, A.F. (1988). Lid-eye coordination during vertical gaze changes in man and monkey. Journal of Neurophysiology, 60, 1227-1252. [ Links ]
Bliss, T.V.P. & Collingridge, G.L. (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature, 361, 31-39. [ Links ]
Bracha, V. & Bloedel, J.R. (1996). The multiplepathway model of circuits subserving the classical conditioning of withdrawal reflexes. En J.R. Bloedel, T.J. Ebner y S.P. Wise (Eds.). The Acquisition of Motor Behavior in Vertebrates (pp. 175-204). Cambridge: The MIT Press. [ Links ]
Clarck, R.E. & Squire, L.R. (1998). Classical conditioning and brain systems: The role of awareness. Science, 280, 77-81. [ Links ]
Delgado-Garcia, J.M. (2000). Why move the eyes if we can move the head? Brain Research Bulletin, 52, 475-482. [ Links ]
Delgado-García, J.M. (2003). Participación del cerebelo y del hipocampo en el condicionamiento clásico del reflejo corneal. Salud Mental, 26, 1-10. [ Links ]
Delgado-García, J.M., Evinger C., Escudero, M. & Baker, R. (1990). Behavior of accessory abducens and abducens motoneurons during eye retraction and rotation in the alert cat. Journal of Neurophysiology, 64, 413-422. [ Links ]
Delgado-García, J.M. & Gruart, E. (2002). The role of interpositus nucleus in eyelid conditioned responses. The Cerebellum, 1, 289-308. [ Links ]
Edwards, F.A. (1995). LTP ma structural model to explain the inconsistencies. Trends in the Neurosciences, 18, 250-255. [ Links ]
Evinger, C., Manning, K.A. & Sibony, P.A. (1991). Eyelid movements. Mechanisms and normal data. Investigative Ophthalmology & Visual Science, 32, 387-400. [ Links ]
Gordon, G. (1951) Observations upon the movement of the eyelids. British Journal of Ophthalmology, 35, 339-351. [ Links ]
Gormezano, I., Kehoe, E.J. & Marshall, B.S. (1983). Twenty years of classical conditioning research with the rabbit. Progress in Psychobiology and Physiological Psychology, 10, 197-275. [ Links ]
Gormezano, I., Schneiderman, N., Deaux, E. & Fuentes, I. (1962). Nictitating membrane: classical conditioning and extinction in the albino rabbit. Science, 138, 33-34. [ Links ]
Gruart, A., Blázquez, P. & Delgado-García, J.M. (1995). Kinematics of spontaneous, reflex, and conditioned eyelid movements in the alert cat. Journal of Neurophysiology, 74, 226-248. [ Links ]
Gruart, A., Blázquez, P., Pastor, A.M. & Delgado-García, J.M. (1994). Very short term potentiation of climbing fiber effects on deep cerebellar nuclei neurons by conditioning stimulation of mossy fiber afferents. Experimental Brain Research, 101, 173-177. [ Links ]
Gruart, A. & Delgado-García, J.M. (1994). Discharge of identified deep cerebellar nuclei neurons related to eye blinks in the alert cat. Neuroscience, 61, 665-681. [ Links ]
Gruart, A., Guillazo-Blanch, G., FernándezMas, R., Jiménez-Díaz, L. & Delgado-García, J.M. (2000a). Cerebellar posterior interpositus nucleus as an enhancer of classically conditioned eyelid responses in alert cats. Journal of Neurophysiology, 84, 2680-2690. [ Links ]
Gruart, A., Morcuende, S., Martínez, S. y Delgado-García, J.M. (2000b). Involvement of cerebral cortical structures in the classical conditioning of eyelid responses in rabbits. Neuroscience, 100, 719-730. [ Links ]
Gruart, A., Muñoz, M.D. & Delgado-García, J.M. (2005). Involvement of the CA3-CA1 synapse in the acquisition of associative learning in behaving mice. 35th Society for Neuroscience Meeting, Washington, USA. [ Links ]
Gruart, A., Pastor, A.M., Armengol, J.A. & Delgado-García, J.M. (1997). Involvement of cerebellar cortex and nuclei in the genesis and control of unconditioned and conditioned eyelid motor responses. Progress in Brain Research, 114, 511-528. [ Links ]
Gruart, A., Schreurs, B.G., Domínguez del Toro, E. & Delgado-García, J.M. (2000c). Kinetic and frequency-domain properties of reflex and conditioned eyelid responses in the rabbit. Journal of Neurophysioogy, 83, 836-852. [ Links ]
Hawkins, R.D., Abrams, T.W., Carew, T.J., Kandel, E.R. (1983). A cellular mechanism of classical conditioning in Aplysia: activitydependent amplification of presynaptic facilitation. Science, 219, 400-405. [ Links ]
Inda, M.C., Delgado-García, J.M. & Carrión, A.M. (2005). Acquisition, consolidation, reconsolidation, and extinction of eyelid conditioning responses require de novo protein synthesis. Journal of Neuroscience, 25, 2070-2080. [ Links ]
Ito, M. (1989). Long-term depression. Annual Review of Neuroscience, 12, 85-102. [ Links ]
Jiménez-Díaz, L., Navarro-López, J. de D.; Gruart, A. & Delgado-García, J.M. (2004). Role of cerebellar interpositus nucleus in the genesis and control of reflex and conditioned eyelid responses. Journal of Neuroscience, 24, 9138-9145. [ Links ]
Lander, T., Wirtschafter, J.D., J.M. & McLoon, M.K. (1996). Orbicularis oculi muscle fibers are relatively short and heterogeneous in length. Investigative Ophthalmology and Visual Science, 37, 1732-1739. [ Links ]
Leal-Campanario, R., Delgado-García, J.M. & Gruart, A. (2005) Substitution of tactile stimuli for somatosensory cortex stimulation during an associative learning task in alert behaving rabbits. 35th Society for Neuroscience Meeting, Washington, USA. [ Links ]
Lennertrand, G. (1975). Motor units in the eye muscles. En G. Lennertand y P. Bach-y-Rita (Eds.). Basic Mechanisms of Ocular Motility. Oxford: Pergamon Press. [ Links ]
Magariños-Ascone, C., Núñez, A. & Delgado-García, J.M. (1999). Different discharge properties of rat facial nucleus motoneurons. Neuroscience, 94, 879-886. [ Links ]
Malenka, R.C. (1995). LTP and LTD: dynamic and interactive processes of synaptic plasticity. The Neuroscientist, 1, 35-42. [ Links ]
McLoon, L.K. & Wirtschafter, J.D. (1991). Regional differences in the orbicularis oculi muscle: conservation between species. Journal of Neurological Science, 104, 197-202. [ Links ]
Morcuende, S., Delgado-García, J.M. & Ugolini, G. (2002). Neuronal premotor networks involved in eyelid responses: retrograde transneuronal tracing with rabies virus from the orbicularis oculi muscle in the rat. Journal of Neuroscience, 22, 8808-8818. [ Links ]
Múnera, A., Gruart, A., Muñoz, M.D., FernándezMas, R., & Delgado-García, J.M. (2001). Hippocampal pyramidal cell activity encodes conditioned stimulus predictive value during classical conditioning in alert cats. Journal of Neurophysiology, 86, 2571-2582. [ Links ]
Navarro-López, J. de D., Alvarado, J.C., Márquez-Ruiz, J., Escudero, M., Delgado-García, J.M., & Yajeya, J. (2004). A cholinergic synaptically triggered event participates in the generation of persistent activity necessary for eye fixation. Journal of Neuroscience, 24, 5109-5118. [ Links ]
Rescorla, R.A. (1988). Behavioral studies of pavlovian conditioning. Annual Review of Neuroscience, 11, 329-352. [ Links ]
Thompson, R.F. (2005). In search of memory traces. Annual Review of Psychology, 56, 1-23. [ Links ]
Thompson, R.F. & Krupa, D.J. (1994). Organization of memory traces in the mammalian brain. Annual Review of Neuroscience, 17, 519-549. [ Links ]
Trigo, J.A., Gruart, A. & Delgado-García, J.M. (1999a). Discharge profiles of abducens, accessory abducens, and orbicularis oculi motoneurons during reflex and conditioned blinks in alert cats. Journal of Neurophysiology, 81, 1666-1684. [ Links ]
Trigo, J.A., Gruart, A. & Delgado-García, J.M. (1999b). Role of proprioception in the control of lid position during reflex and conditioned blink responses in the alert behaving cat. Neuroscience, 90, 1515-1528. [ Links ]
Trigo, J.A., Roa, L., Gruart, A., & Delgado-García, J.M. (2003). A kinetic study of blinking responses in cats. Journal of Physiology (London), 549, 195-205. [ Links ]
Woody, C.D. (1986). Understanding the cellular basis of memory and learning. Annual Review of Psychology, 37, 433-493. [ Links ]
Recepción: Julio de 2005
Aceptación final: Octubre de 2005
1 Este trabajo ha sido realizado con una ayuda del MCyT (BFI2002-00936).
2 Correspondencia: Prof. José M. Delgado-García, División de Neurociencias, Universidad Pablo de Olavide. Ctra. de Utrera, Km. 1. 41013Sevilla, España Tlfno: + 34954349374. Fax: + 34954349375. Correo electrónico: jmdelgar@upo.es
