










Services on Demand
article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.17 no.2 Guadalajara 2009
Consumo de soluciones endulzadas en octodones (Octodón-degú)1
Consumption of sweetened solutiones by octodones
Alma Gabriela Martínez*; Juan Antonio Madrid**; Antonio López-Espinoza*; Pablo Vivanco**
Universidad de Guadalajara *
Universidad de Murcia**
RESUMEN
Ratas expuestas a agua endulzada con glucosa presentan grandes bebidas, mientras que ratas expuestas a agua endulzada con sucralosa no presentan grandes bebidas. El resultado es similar a pesar de modificar la secuencia de presentación del endulzante. El objetivo de este trabajo fue replicar los resultados obtenidos en ratas con otra especie de roedor: el Octodón-degú, que tiene características alimentarias particulares como beber pocas cantidades de agua y no metabolizar adecuadamente los azúcares. Los resultados mostraron que los sujetos emitieron grandes bebidas de agua endulzada con glucosa, mientras que mostraron un incremento en el consumo de agua endulzada con sucralosa el primer día de acceso al endulzante sin calorías. Estos resultados confirman que el consumo de grandes cantidades de soluciones endulzadas con glucosa puede observarse experimentalmente en otra especie de roedor.
Palabras-clave: Consumo, soluciones endulzadas, glucosa, sucralosa, octodón-degú
ABSTRACT
Rats exposed to sweetened water with glucose show binge drinking response whereas rats exposed to sweetened water with sucralose do not show binge drinking response. This result is similar although it modified the sweetener presentation sequence. The objective of this work was to replicate the obtained results in rats with other species of rodent: Octodon-Degus, who has peculiar feeding characteristics like they drink few water and they do not metabolize properly the sugars. Results showed the subjects emit binge drinking response of sweetened water with glucose whereas they showed an increase of sweetened water with sucralose only the first day of access to the sweetener without calories. These results confirm that great quantity of sweetened solution intake can showed in another species of rodent experimentally
Keywords: Intake, sweetened solutions, glucose, sucralose, octodon-degusoperant variability; induced variability; extinction; behavioral history; positive reinforcement; animal behavior, rats
Desde un enfoque experimental, los endulzantes como la sucrosa, el aspartame, la sacarina y la sucralosa han sido utilizados para evaluar sus efectos en la conducta alimentaria (Hirsh, Duboise y Jacobs, 1982; Rothschild, 1971), ya sea por: su sabor (Booth, 1990), por su contenido energético (Bolles, Hayward y Crandall, 1981; Carper y Polliard, 1953), nutricional (Sclafani, 1990), o bien, para evaluar sus efectos como reforzador de conductas operantes (Guttman, 1954; Sheffield y Thornton, 1950). Le- Magnen (1999) afirmó que el sabor dulce activa ciertas funciones sensoriales que educen la conducta alimentaria. Señaló que los alimentos dulces son calificados como palatables y preferidos, pues estimulan la respuesta de comer y son percibidos como placenteros
Con base en estas evidencias, diversos investigadores se han interesado en estudiar la conducta de consumo de endulzantes en animales de laboratorio. Por un lado, se ha reportado que los animales muestran grandes consumos de soluciones endulzadas (Hamilton, Timmons y Lerner, 1980), y por el otro, también se ha reportado que los grandes consumos dependen de la calidad del endulzante (Sclafani, 1990). Martínez (2007) y Martínez, López-Espinoza, Díaz y Valdés (2009) demostraron que ratas expuestas a agua endulzada con glucosa (sabor dulce con calorías) incrementan su respuesta de consumo considerablemente respecto a la respuesta observada cuando son expuestas a agua o a agua endulzada con sucralosa (sabor dulce sin calorías). El resultado fue similar a pesar de modificar la secuencia de presentación del endulzante. No obstante, el potencial efecto de los endulzantes naturales o artificiales sobre la emisión de respuestas alimentarias no ha sido lo suficientemente estudiado aún. Al parecer, los efectos del consumo de bebidas endulzadas sobre la conducta alimentaria resulta una línea de investigación relevante para determinar qué propiedades de los endulzantes pueden modificar o desordenar el patrón alimentario de ciertos roedores. A partir de estas evidencias, el objetivo de este experimento es replicar los resultados obtenidos en ratas con otra especie de roedor: el Octodón-degú, que poseé características alimentarias particularmente especiales para el estudio experimental del consumo de soluciones endulzadas.
Los octodones (o degús) son roedores herbívoros y pertenecen al grupo de los octodóntidos. Estos animales proceden de América del sur, específicamente de Perú y Chile. Se han utilizado como animales de laboratorio para diversos estudios sobre ritmos circadianos y termorregulación en los que se ha señalado que presentan gran adaptabilidad a regímenes climáticos divergentes pero que muestran una limitada capacidad de termorregulación. Su temperatura corporal habitual es de 36-37º (Kenagy, Nespolo, Vásquez y Bozinovic, 2002).
Una de sus características de mayor interés científico es su dificultad para metabolizar los azúcares. Los octodones pueden consumir algunas frutas y vegetales, pero si tienen alto contenido de azúcares pueden sufrir alteraciones en su aparato digestivo. Cortés, Rosenmann y Bozinovic (2000) reportaron que los octodones representan una especie propensa a desarrollar diabetes. Otra de sus características alimentarias consiste en su limitada respuesta para beber agua. Generalmente, los octodones salvajes obtienen el agua de los vegetales que comen. Por el contrario, los octodones de laboratorio beben pocas cantidades de agua cuando son alimentados con comida especial para animales de laboratorio y dejan de beber agua cuando son alimentados con vegetales. Bozinovic y Gallardo (2006) observaron que dentro y fuera del laboratorio, los octodones son incapaces de mostrar grandes consumos de agua. En este sentido, es de nuestro interés conocer si el octodón puede modificar su conducta ingestiva de líquidos. Si el octodón muestra grandes consumos de agua endulzada podría indicar que ante fuertes estímulos ambientales (sabor, contenido energético) puede modificar conductualmente su patrón de consumo; o bien, si no se presentan las grandes bebidas, podría indicar que esta especie no modifica su respuesta de consumo ni siquiera ante estímulos poderosos como el sabor o el contenido energético de los endulzantes. Adicionalmente, será posible comparar los patrones alimentarios de bebida de endulzantes entre ratas y octodones. La finalidad es la de profundizar en el conocimiento de la respuesta de ingestión de bebidas endulzadas en roedores. Por ello, el objetivo de esta investigación es determinar el efecto del consumo de endulzantes (nutritivos y no nutritivos) sobre el consumo de alimento, agua, peso corporal y temperatura en octodones. El conocimiento de estos efectos permitirá responder otras interrogantes referentes a los grandes consumos de agua con glucosa observados en ratas, pues se establecerán nuevos parámetros útiles para describir este fenómeno.
MÉTODO
Sujetos
Ocho octodones: cuatro hembras y cuatro machos ingenuos y con un año y cuatro meses de edad al inicio del experimento. Los animales fueron proporcionados por el animalario de la Universidad de Alicante.
Aparatos y materiales
Se utilizaron ocho cajas-habitación individuales con una reja metálica en la parte superior y una división para comederos y bebederos. Para el registro del consumo de alimento y peso corporal se utilizó una báscula electrónica de precisión. Para el registro de la temperatura corporal se implantaron botones especiales para registros térmicos. Estos botones tienen un diámetro de 17mm y 3,3 g de peso, registran la temperatura de forma autónoma y no requieren de baterías o dispositivos especiales. Se implantan en el sujeto mediante un procedimiento quirúrgico. Pienso especial para roedores de laboratorio de la marca Harlan global diet fue el alimento proporcionado. Su fórmula nutricional es la siguiente: agua 12%, proteínas 14.6%, grasas 4%, fibra 4.5% y cenizas 4.7%. Como bebida se utilizó agua y durante la manipulación experimental se proporcionó una solución compuesta de glucosa diluida en agua al 8% (15g de glucosa por cada 200ml) y una solución de agua con sucralosa al 2% (4g de sucralosa por cada 200ml). El líquido se proporcionó en bebederos graduados de 200ml con boquillas antiderrames especiales para impedir el paso de aire.
Procedimiento quirúrgico
Bajo inducción anestésica con Isofluorane se colocó al sujeto experimental en una plancha quirúrgica. Previa asepsia y antisepsia de la región se incidió por planos hasta cavidad peritoneal. Se colocó el botón para registro térmico (marca Ibuton). Se cerró por planos con Dexon 000, se afrontó la piel con Seda negra 00. Se aplicó asepsia con Isodine y se colocó al sujeto en su caja habitación para su recuperación. Este procedimiento se utilizó para colocar y extraer el botón térmico.
Procedimiento
Los sujetos fueron identificados con un número de registro, fecha de nacimiento y peso corporal al inicio del experimento y fueron colocados en cajas-habitación individuales. El consumo de agua y comida se registró diariamente a las 8:00 de la mañana. Para el pesaje se tomaba la caja habitación y se trasladaba a la mesa de trabajo en la que se encontraba la báscula utilizada para el registro. Se registró el consumo de alimento por gramo consumido y el consumo de agua por mililitro consumido. Para calcular las calorías consumidas en el alimento se determinó el número de calorías que aporta el alimento (Harlan global diet) por gramo (3.3 calorías). Esta cantidad se multiplicó por el número de gramos consumidos diariamente para obtener el consumo de calorías diario. El registro de la temperatura se obtuvo automáticamente con los botones implantados que se retiraron al finalizar el experimento y que fueron colocados con software especializado para ser analizados en computadora mediante el programa iButton viewer.
El experimento se dividió en cinco fases y tuvo una duración de 21 días. Todos los sujetos recibieron 50g de alimento y 200ml de bebida disponible diariamente bajo condiciones de libre acceso. Se formaron dos grupos conformados por cuatro sujetos (dos hembras y dos machos). En la primera fase, el Grupo A recibió alimento y agua durante cinco días consecutivos. En la segunda fase fueron expuestos alimento y una solución compuesta de agua+glucosa durante tres días. Retornaron a condiciones de línea base durante cinco días en la tercera fase. En la cuarta fase recibieron alimento y una solución compuesta de agua+sucralosa durante tres días. Finalmente recibieron alimento y agua en la última fase. El Grupo B recibió el mismo tratamiento, excepto que durante la segunda fase fueron expuestos a la solución de agua+sucralosa y durante la cuarta fase a la solución agua+glucosa. El número de días de acceso a los endulzantes fue más breve que el utilizado en las ratas albinas debido a las características alimentarias del octodón.
RESULTADOS
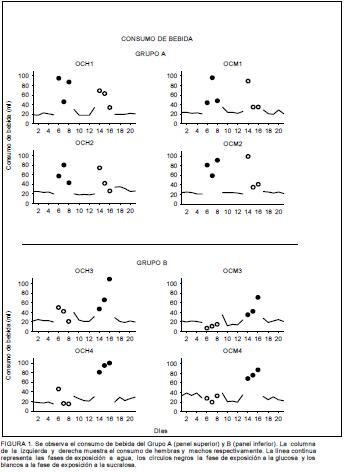
La Figura 1 muestra el consumo de líquido del Grupo A, que recibió la secuencia glucosa-sucralosa, en el panel superior. Los sujetos mantuvieron estable el consumo de agua durante las fases 1, 3 y 5, consumiendo un promedio de 20ml. No se observaron variaciones en el consumo de agua antes o después de adicionar glucosa o sucralosa. Durante la fase 2 en la que se adicionó glucosa al agua, los sujetos incrementaron su consumo de líquido respecto al consumo registrado en las fases sin el endulzante promediando un consumo de 65ml. El patrón de consumo de agua+glucosa fue variable en los cuatro sujetos. Para los sujetos OCH1 y OCM2 inició con un incremento seguido de un decremento y un nuevo incremento en el consumo. Para los sujetos OCH2 y OCM1 el consumo de agua+glucosa se incrementó gradualmente los primeros dos días de acceso, finalizando con un leve decremento. No obstante la reducción en el consumo de agua+glucosa no disminuyó hasta alcanzar el promedio de consumo de agua de las fases 1, 3 y 5. Durante la fase 4 los sujetos recibieron la sucralosa en el agua. Se observó que durante el primer día de acceso, todos los sujetos incrementaron su consumo de bebida que disminuyó gradualmente hasta alcanzar el promedio de consumo de agua de las fases 1, 3 y 5 en las hembras, mientras que en los machos alcanzó este promedio de consumo desde el segundo día de acceso
El consumo de líquido del Grupo B se muestra en el panel inferior de la Figura 1. Los sujetos consumieron un promedio de 20ml de agua durante las fases 1, 3 y 5. Cuando recibieron la sucralosa en la Fase 2, las hembras consumieron 50ml en el primer día de acceso, es decir, duplicaron su consumo de bebida. Posteriormente disminuyeron su consumo gradualmente llegando a consumir cantidades inferiores a las registradas durante las fases 1, 3 y 5, en las que solo tenían disponible agua. Por su parte los machos no incrementaron su consumo de agua+sucralosa respecto a su consumo de agua. De forma particular, el sujeto OCM1 redujo su consumo de agua+sucralosa respecto a su consumo de agua durante la fase 1. Cuando los sujetos tuvieron disponible la glucosa en la Fase 4, incrementaron su consumo de bebida gradualmente durante los tres días. El sujeto OCH3 rebasó los 100ml de consumo el último día de acceso, mientras que los sujetos OCH4 y OCM4 registraron 100ml de consumo máximo durante el último día de acceso. El promedio de consumo de agua+glucosa del Grupo B fue de 75ml.
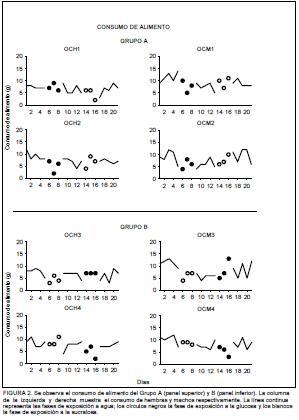
La Figura 2 muestra el consumo de alimento del Grupo A, que recibió la secuencia glucosa-sucralosa, en el panel superior. Durante las fases 1, 3 y 5 los sujetos consumieron un promedio de 10g de alimento diariamente. Durante la manipulación experimental no se observaron variaciones en el consumo. Cuando tuvieron disponible la glucosa en la Fase 2, los sujetos OCM1 y OCH2 disminuyeron su consumo por debajo del promedio mencionado en el segundo día de acceso de forma general, el Grupo A consumió un promedio de 6g durante la fase de acceso a la glucosa. Por su parte, cuando recibieron la sucralosa en la Fase 4, el sujeto OCH1 disminuyó su consumo de alimento durante el tercer día de acceso, mientras que el sujeto OCH2 lo hizo en el primer día de acceso.
El consumo de alimento del Grupo B, que recibió la secuencia sucralosa-glucosa se representa en el panel inferior de la Figura 2. Los sujetos consumieron un promedio de 10g de alimento durante las fases 1, 3 y 5. El promedio de consumo se mantuvo igual durante la Fase 2 en la que se adicionó la sucralosa al agua. Específicamente el sujeto OCM3 disminuyó su consumo de alimento el primer día de acceso a la sucralosa. Durante la Fase 4, los sujetos mostraron variabilidad en su patrón de consumo. De forma particular, el sujeto OCH3 mantuvo estable su consumo de alimento de 10g, el sujeto OCH4 disminuyó su consumo durante el último día de acceso, el sujeto OCM3 incrementó gradualmente su consumo durante los tres días de acceso hasta llegar a los 15g; finalmente el sujeto OCH4 disminuyó gradualmente su consumo durante los tres días de acceso a la glucosa hasta alcanzar 1g de consumo de alimento.
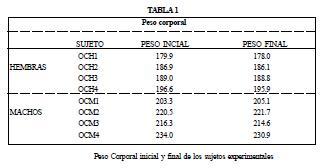
En la tabla 1 se muestra el peso corporal inicial y final de cada sujeto. Se observó que todos los sujetos no mostraron aumentos o decrementos importantes de su peso corporal final respecto al peso corporal inicial registrado al inicio del experimento.
Para os sujeitos de um grupo (G1), o reforço era liberado contingente à seqüência
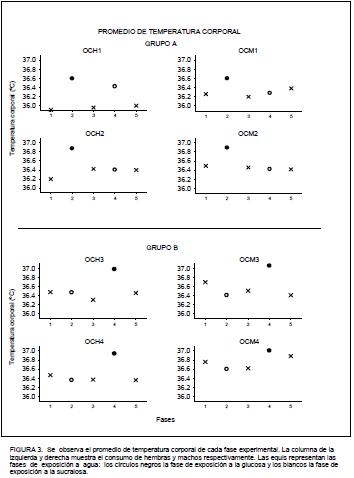
En la Figura 3 se representa el promedio de temperatura corporal de cada sujeto. Se obtuvo el registro de la temperatura corporal diariamente durante cada hora. Para obtener la media que se observa en la figura se promediaron las veinticuatro mediciones diarias y posteriormente se separaron los días en lo que se tuvo acceso a agua, a la glucosa y a la sucralosa para obtener el promedio final. De forma general, se observó que todos los sujetos incrementaron la temperatura cuando tuvieron acceso a la glucosa, mientras que cuando fueron expuestos a la sucralosa no mostraron diferencias en el registro de temperatura observado durante las fases de acceso a agua.
DISCUSIÓN
Los resultados obtenidos mostraron: 1) incremento gradual en el consumo de agua con glucosa respecto al consumo de agua o agua con sucralosa; 2) incremento en el consumo de agua con sucralosa el primer día de acceso al endulzante en el Grupo A; 3) decremento gradual en el consumo de agua endulzada con sucralosa en hembras del Grupo A; 4) decremento abrupto en el consumo de agua endulzada con sucralosa a partir del segundo día de acceso al endulzante en machos del Grupo B; 5) consumo de alimento estable durante todo el experimento; 6) ninguna modificación en el peso corporal registrado al inicio y final del experimento; y, 7) incremento de la temperatura corporal durante las fases de acceso a la glucosa respecto a las fases de acceso a agua y sucralosa Estos resultados confirman que el consumo de grandes cantidades de soluciones endulzadas con glucosa puede observarse en otra especie de roedor. Los estudios realizados anteriormente con ratas muestran una similitud en las respuestas ingestivas de agua endulzada con glucosa observadas en los octodones. Esta respuesta, al igual que la de consumir grandes cantidades de agua ha sido denominado por algunos autores como gran bebida y a pesar de que no se ha definido operacionalmente, se ha reportado en otras especies: gerbiles (Bouskila, 2001), chimpancés (Bonthius, Bonthius y Napper, 1996), iguanas (Carpenter, 1966) y diversas especies de aves (Cade y Green- Wold, 1966; Poulsen, 1953; Wickler, 1985). Se ha señalado que esta conducta es resultado de la adaptación conductual de los animales a climas áridos (Hallager, 1994). Sin embargo, otros investigadores afirmaron que la presencia de esta conducta en animales y humanos no depende únicamente de factores geográficos, sino que involucra otros factores relacionados con el horario, disponibilidad y sabor de los alimentos (Bolles, 1983 Por su parte, Bozinovic y Gallardo (2006) señalaron que la modificación de los estímulos ambientales puede alterar el patrón alimentario de diversos animales, y par-ticularmente, del octodón. En este experimento, la adición de un sabor dulce y un efecto postingestivo en la dieta recibida por el sujeto determinó su modificación de respuestas ingestivas. Los sujetos experimentales mostraron consumos considerables de agua endulzada con glucosa. Esta conducta suele presentarse cuando los animales reciben un alimento nuevo y palatable. Numerosos autores coinciden en afirmar que los animales prefieren alimentos dulces o palatables por las propiedades hedónicas y postingestivas que contienen (Bolles, Hyward y Crandall, 1981; Booth, 1990; Sclafani, 1990). No obstante, los resultados obtenidos mostraron que el sabor dulce no es suficiente para que los animales mantengan las "grandes bebidas". Cuando se adicionó la sucralosa a la dieta, los animales mostraron grandes bebidas solo el primer día de acceso y posteriormente se observó un decremento en el consumo de bebida gradual (en las hembras) o abrupto (en los machos). Es decir, los animales no obtuvieron una consecuencia postingestiva de la solución endulzada con sucralosa y dejaron de beber grandes cantidades. Este registro fortalece el argumento de que el contenido energético del alimento es un estímulo más fuerte que el sabor para observar y mantener respuestas de consumo. Adicionalmente, no se observó ningún cambio en el peso corporal de los sujetos, a pesar de que aumentaron considerablemente su ingestión de calorías durante las fases en las que recibieron la glucosa. Esto indica que la expedición de energía tuvo que incrementarse para evitar el aumento de peso corporal en los sujetos. Posiblemente, la medición del nivel de actividad de los sujetos podría haber aclarado este hecho Otra evidencia que parece fortalecer este argumento es el aumento en la temperatura corporal registrada durante las fases de acceso al endulzante respecto a las fases de acceso a la sucralosa y a agua. Se ha señalado que la temperatura corporal habitual del octodón oscila entre los 36 y 37º. Si bien el registro de temperatura obtenido los días en que los animales fueron expuestos a la glucosa no sobrepasó este límite, sería importante indagar si, efectivamente, la glucosa representa un estímulo potencial para modificar la temperatura del octodón, pues el resultado observado es cuestionable. De forma general, los estudios realizados sobre la termorregulación en octodones consisten en modificar la temperatura ambiente y determinar cómo se lleva a cabo este proceso (Kenagy, Nespolo, Vásquez y Bozinovic, 2002; Kenagy, Vásquez, Nespolo y Bozinovic; 2002). Los datos obtenidos en el presente experimento demuestran que el cambio de dieta también podría involucrarse en su adaptación a diversos climas. Por otro lado, parece que la presentación del endulzante puede modificar también la respuesta ingestiva. La historia previa de sabor dulce pudo establecer la presencia de grandes bebidas de agua con sucralosa en los sujetos que recibieron la secuencia glucosa-sucralosa, respuesta que no se presentó en el grupo que recibió la secuencia sucralosa-glucosa. Sin embargo, es necesario establecer nuevas manipulaciones experimentales para determinar si la historia de alimentación previa puede intervenir en el consumo, selección, preferencias o aversiones de bebidas endulzadas. De igual forma, es importante establecer un criterio para definir operacionalmente el concepto de gran bebida y poder aplicarlo adecuadamente en la investigación experimental de respuestas ingestivas
REFERENCIAS
Bolles, R.C. (1983). Teoría de la motivación. México: Trillas. [ Links ]
Bolles, R.C., Hayward, L., y Crandall, C. (1981). Conditioned taste preferences based on caloric density. Journal of Experimental Psychology:Animal Behavior Processes, 7, 59-69. [ Links ]
Bouskila, A. (2001). A habitat selection game interactions between rodents and predators. Annals of Zoology, 38, 55-70. [ Links ]
Bonthius, D. A., Bonthius, F. H. y Napper, M. A. (1996). Alcohol induced neuronal lost in developing rat: Increase brain damage. Alcoholism: Clinical and Experimental Research, 114, 107-111. [ Links ]
Booth, D. (1990). Learned tastes in eating motivation. En E.D. Capaldi, y T.L. Powley (eds.). Taste, experience & feeding: development and learning. USA: American Psychological Association. [ Links ]
Bozinovic, F. y Gallardo, J. C. (2006). Fisiología ecológica de la alimentación y digestión en vertebrados: modelos y teorías. Revista Chilena de Historia Natural, 66, 375-382). [ Links ]
Cade, T. J. y Green-Wold, L. I. (1966). Drinking behavior of mouse birds in the Namibidesert, southern Africa. Wilson Bulletin, 83, 126-128 [ Links ]
Carper, J., y Polliard, F. (1953). A comparison of the intake of glucose and saccharin solutions in conditions of caloric need. American Journal of Psychology, 66, 479-482. [ Links ]
Carpenter, C. C. (1966). The main iguanas of the Galápagos Islands, its behavior and ecology. California Academic Sciences, 4, 329-375. [ Links ]
Cortés, A., Rosenmann, M. y Bozinovic, F. (2000). Water economy in rodent: evaporative water loss and metabolic water production. Revista Chilena de Historia Natural, 73, 24-31. [ Links ]
Ebensperguer, L.A., Hurtado, M., Soto-Gamboa, E. A., Lacey, A. T. y Chang, M. (2004). Comunal nesting and kinship in degus (Octodón degus). Naturwissenschaften, 91, 391-395. [ Links ]
Guttman, N. (1954). Equal-reinforcement values for sucrose and glucose solutions compared with equal sweetness values. Journal of Comparative and Physiological Psychology, 47, 358-361. [ Links ]
Hallager, S. L. (1994). Drinking methods in two species of bustords. The Wilson Bulletin, 106, 764-766. [ Links ]
Hamilton, L.W., Timmons, C.R., y Lerner, S.M. (1980). Caloric consequences of sugar solutions: A failure to obtain gustatory learning. American Journal of Psychology, 93, 387-407. [ Links ]
Hirsch, E., Duboise, C., y Jacobs, H. L (1982). Overeating, dietary selection patterns and sucrose intake in growing rats. Physiology and Behavior, 28, 819-833. [ Links ]
Kenagy, G. J., Nespolo, R. F., Vásquez, R. A. y Bozinovic, F. (2002). Daily and seasoneal limits of time and temperature to activity of degus. Revista Chilena de Historia Natural, 75, 567-581. [ Links ]
Kenagy, G. J., Vásquez, R. A., Nespolo, R. F. y Bozinovic, F. (2002). A time-energy analysis of daytime surface activity in degus, Octodon degus. Revista Chilena de Historia Natural, 75, 149-156. [ Links ]
Le-Magnen, J. (1999). Role of dietary odour in the short-term control of intake in the white rat. Appetite, 33, 30-32. [ Links ]
Martínez, A. G. (2008). Análisis experimental de la gran bebida de endulzante es ratas (Rattus norvegicus) y degús (Octodón-degus). Tesis Doctoral, Universidad de Guadalajara, Jalisco, México. [ Links ]
Martínez, A. G., López-Espinoza, A., Díaz, F. y Valdés, E. (aceptado). Consumo de soluciones endulzadas en ratas albinas, Psicothema. [ Links ]
Mook, D. G. (1974). Saccharin preference in the rat: some unpalatable findings. Psychological Review, 81, 475-490. [ Links ]
Poulsen, H. (1953). A study of incubation responses and some other behavior patterns in birds. Wilson Bulletin, 115, 11-31. [ Links ]
Rothschild, G.H. (1971). Patterns of concentration preference for glucose and sodium chloride in rats. The psychological record, 21, 487-496. [ Links ]
Sclafani, A. (1990). Nutritionally based learned flavor preferences. En E.D. Capaldi, y T.L. Powley (eds.). Taste, experience & feeding: development and learning. USA: American Psychological Association. [ Links ]
Sheffield, F. D., y Thornton, R. (1950). Reward value of a non-nutritive sweet taste. Journal of Comparative and Physiological Psychology, 43, 471-481. [ Links ]
Veloso, C. y Bozinovic, F. (2000). Effect of food quality on the energetics of reproduction in a presocial rodent, Octodon degus. Journal of Mammalogy, 81, 971-978. [ Links ]
Wickler, W. 1(985). Coordination of vigilance in bird groups. Z.Tierpsychol. 69, 250-253. [ Links ]
Received January 22, 2009
Accepted March 11, 2009
1Esta investigación fue financiada por la Fundación SENECA (código PB/43/FS/02) y por el Programa de Movilidad Estudiantil para realizar estancias de investigación de la Coordinación General de Cooperación e Internacionalización de la Universidad de Guadalajara.
Correspondencia: Dra. Alma Gabriela Martínez Moreno.Centro de Investigaciones en Comportamiento Alimentario y Nutrición.Centro Universitario del Sur. Universidad de Guadalajara. Prolongación Colón s/n, Edificio X-3,Zapotlán el Grande, Jalisco. México.
Tel. 01(341) 57 522 22 extensión 6127 e-mail alma.martinez@cusur.udg.mx
