










Services on Demand
article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.18 no.3 Guadalajara 2010
ARTÍCULOS
Formação de classes funcionais em cães domésticos (Canis familiaris): uma abordagem da aquisição de comportamento pré-simbólico1
Functional class formation in domestic dogs (Canis familiaris): an approach to the acquisition of pre-symbolic behavior
Liane Jorge de Souza Dahás2; Izabel Cristina da Silva Brasiliense; Romariz da Silva Barros; Thiago Dias Costa; Carlos Barbosa Alves de Souza
Universidade Federal do Pará
RESUMO
O presente estudo investigou a possibilidade de ensinar dois cães domésticos (Canis familiaris) a responderem adequadamente a duas classes de três estímulos (caixas contendo pedaços de alimento quando com função positiva) através de um procedimento de mudanças sucessivas de discriminação simples (MSDS). A coleta de dados foi realizada em uma sala na qual duas, quatro ou seis caixas de madeira eram dispostas lado a lado, em posições fixas. Inicialmente, os sujeitos responderam a dois estímulos por sessão (um positivo [S+] e outro negativo[S-]). Posteriormente quatro das seis caixas eram apresentadas, duas com função S+ e outras duas com função S-. Na fase seguinte, foi realizado um treino de MSDS com as quatro caixas. Em seguida os sujeitos foram expostos a um treino de MSDS com as seis caixas. Foram realizados sete testes. Cada um deles consistia em (1) uma sessão de mudança de função de estímulo com quatro caixas, e então (2) eram re-inseridas as duas caixas restantes na sala experimental. Um sujeito respondeu adequadamente a três desses sete testes e o outro a quatro deles. Discutem-se os efeitos da ênfase na modalidade visual de discriminação e o uso de posições fixas das caixas sobre o desempenho dos sujeitos.
Palavras chave: Cognição animal, classes funcionais, mudanças sucessivas de discriminação simples, cães domésticos.
ABSTRACT
The present study investigated the possibility of teaching two domestic dogs (Canis familiaris) to respond adequately to two classes of three stimuli (wooden boxes with pieces of food when having S+ function) through a procedure of successive shifts of simple discrimination. Sessions were carried out in a room where two, four, or six wooden boxes were placed side by side. Initially, the subjects responded to two stimuli (a positive [S+] and another negative [S-]) for session. Next, four of the six boxes were presented, two functioning as S+ and other two functioning as S-. In the following phase, successive shifts of simple discrimination were carried out with the four boxes. After that, the training with all six boxes was initiated. Seven tests were carried out. Each test consisted of (1) a discrimination shift with four boxes, with the two remaining boxes kept out; (2) then the two remaining boxes were re-inserted in the experimental room. One of the subjects adequately responded to three of these seven tests and the other to four of them. It is discussed the effect of the emphasis in the visual modality of discrimination and the use of fixed positions of the boxes on the performance of the subjects.
Key words: Animal cognition, functional classes, successive shifts of simple discrimination, domestic dogs.
O desenvolvimento de repertórios simbólicos pode ter representado uma vantagem evolutiva no processo de seleção das espécies (Deacon, 1997; Hauser, Chomsky, & Fitch, 2002). O comportamento simbólico pode ser definido como o responder a estímulos arbitrariamente relacionados e diferentes entre si, de maneira que estes se tornem substituíveis em determinados contextos (Barros, Galvão, Brino, Goulart, & McIlvane, 2005).
A formação de classes equivalentes de estímulos é um modelo experimental que tem sido amplamente usado na Análise do Comportamento para estudar o comportamento simbólico (e.g. Frank & Wasserman, 2005). Em um estudo típico desse fenômeno, primeiro são ensinadas relações arbitrárias entre estímulos (geralmente por meio de um procedimento de pareamento ao modelo3) e posteriormente são testadas relações não ensinadas diretamente (Sidman & Tailby, 1982). De acordo com Sidman e Tailby, os estímulos terão se tornado equivalentes se, após o ensino das relações arbitrárias entre eles, for observada em testes a emergência de relações reflexivas (ex. escolher X dado X como modelo), simétricas (ex. escolher Y dado X como modelo, após o ensino da relação X-Y) transitivas e simétrico-transitivas (ex. escolher Z dado X como modelo e vice versa, após o ensino das relações X-Y e Y-Z) entre esses estímulos.
Apesar de a equivalência de estímulos ser um modelo de estudo do comportamento simbólico utilizado há mais de 25 anos na área da Análise do Comportamento, algumas questões teóricas e empíricas relacionadas ao fenômeno ainda estão por serem elucidadas. Por exemplo, ainda não foi totalmente esclarecido se a equivalência de estímulos depende da existência de um repertório verbal (e.g. Horne, Lowe, & Harris, 2007) ou se a equivalência é um processo comportamental básico, pré-requisito para outros repertórios comportamentais (Sidman 2000).
Outra discussão na área trata da relação entre classes de estímulos funcionais (estímulos que exercem a mesma função sobre uma classe comportamental – ver Catania, 1999) e classes de equivalência. Vaughan (1988) questionou se o procedimento estipulado por Sidman e Tailby (1982) seria o único capaz de estabelecer classes de estímulos equivalentes. Segundo Vaughan (1988), um procedimento que formasse classes funcionais, comparado a outro que formasse classes de equivalência, na verdade estaria resultando no mesmo fenômeno através do procedimento de reforçamento diferencial de respostas a conjuntos de estímulos. A diferença estaria somente nos procedimentos empregados: via testes de reflexividade, simetria e transitividade para a classe de equivalência, e por meio da partição do conjunto de estímulos (responder de forma generalizada para os estímulos de um conjunto e discriminadamente entre estímulos de conjuntos diferentes) para a classe funcional.
Vaughan (1988) realizou um estudo de formação de classes funcionais em pombos. Um procedimento de discriminação simples Go/No-go4, com 40 slides de fotografi as de árvores divididos em 2 conjuntos de 20 slides, foi implementado. Foram utilizados seis pombos como sujeitos. Era reforçada a resposta de bicar duas vezes, em um intervalo de 2 segundos, na chave de resposta quando os estímulos do conjunto designado como positivo (S+) eram apresentados. Respostas de bicar quando os S-(s) eram apresentados produziam sua re-apresentação por mais 2 segundos, e não responder durante os S-(s) produzia o fim da tentativa. Após as 14 sessões iniciais (nas quais os estímulos do Conjunto 1 eram os S+), a cada sete sessões as funções dos estímulos foram revertidas. Foram realizadas 21 reversões no total. Após isso, passaram-se a reverter as funções dos estímulos a cada quatro sessões, o que ocorreu durante 95 reversões. Por fim, para evitar que os sujeitos respondessem à temporalidade das reversões, foram realizadas reversões de forma randômica (após cinco, seis ou sete sessões) durante 43 reversões.
Os resultados mostraram que, com o avanço no treino, os pombos passaram, gradualmente, a ajustar suas respostas às funções dos estímulos na sessão: logo no início da sessão, quando respostas aos S- não produziam conseqüência reforçadora, nas tentativas seguintes o pombo passava a responder aos S+ e a não responder aos S-. Para Vaughan (1988) esses resultados demonstravam que os sujeitos de fato respondiam aos estímulos de cada conjunto como sendo parte de uma classe, indicando a formação de classes funcionais ou de estímulos equivalentes.
No entanto, de acordo com Hayes (1989), o procedimento de Vaughan (1988) mostrou a formação de classes funcionais, mas não de equivalência, uma vez que não foram realizados os testes formais de relações emergentes (reflexividade, simetria e transitividade). Além disso, Hayes apontou que, devido ao fato de terem sido realizadas muitas sessões, é possível que cada uma das relações entre os estímulos pertencentes a cada conjunto tenha sido treinada explicitamente.
Sidman, Wynne, Maguire e Barnes (1989) buscaram avaliar com maior precisão a relação entre a formação de classes funcionais e classes de equivalência. Participaram do estudo uma mulher adulta com atraso no desenvolvimento e dois adolescentes autistas. Utilizando um procedimento de mudanças5 sucessivas das funções dos estímulos em discriminações simples simultâneas, inicialmente foram estabelecidas duas classes funcionais de estímulos. Depois, os mesmos estímulos pertencentes às classes funcionais foram utilizados em um procedimento de pareamento ao modelo (tanto por identidade quanto por relações arbitrárias), de maneira que, quando um estímulo de uma classe aparecia como modelo, os estímulos dessa mesma classe que apareciam como comparação, tinham função positiva, enquanto os estímulos da outra classe que aparecessem como comparação tinham função negativa. As respostas dos três participantes mostraram a emergência de relações condicionais entre os estímulos das mesmas classes funcionais.
Em seguida, dois estímulos novos foram incluídos em cada classe. Cada participante recebeu um treino no qual o estímulo novo era o modelo e alguns estímulos da classe funcionavam como comparação correta (S+). Os testes ocorreram colocando-se os outros estímulos membros da classe funcional como modelos e o estímulo novo como comparação. Dois dos três participantes passaram nos testes de emergência de propriedades de equivalência.
Por último, dois novos estímulos foram incluídos em cada classe de equivalência (através do pareamento com os estímulos novos do teste anterior). Um teste foi realizado para verificar se os dois participantes que passaram nos testes de equivalência responderiam a esses estímulos novos como integrantes das classes funcionais correspondentes às classes de equivalência nas quais foram incluídos. Para isso, os estímulos novos foram adicionados aos anteriores em um teste de mudanças sucessivas de discriminações simples. Os desempenhos dos dois participantes mostraram evidência da inclusão dos novos estímulos nas classes funcionais correspondentes.
Kastak, Schusterman e Kastak (2001) realizaram um estudo semelhante ao de Sidman et al. (1989), utilizando como sujeitos dois leões-marinhos, com história experimental de pareamento arbitrário e teste de simetria e transitividade (Schusterman & Kastak, 1993). No Experimento 1, foi realizado um procedimento semelhante ao de Vaughan (1988), mas empregando um treino de mudanças sucessivas de discriminações simples simultâneas (com duas escolhas). Vinte fi guras (letras e números) foram divididas em dois conjuntos de 10 estímulos (letras vs. números). Os dois sujeitos passaram a responder de maneira que indicava a formação de classes funcionais, ou seja, após uma mudança, logo no início da sessão as conseqüências de responder aos S- (os S+ da sessão anterior) eram suficientes para estabelecer o responder aos novos S+. Nesse experimento, foi manipulado ainda (em diferentes fases) o uso de reforçadores específicos para as classes. Ambos os sujeitos responderam com maior precisão nas fases nas quais o reforçamento era específico para a classe.
O Experimento 2 avaliou inicialmente se as classes funcionais estabelecidas no Experimento 1 poderiam ser verificadas em uma situação de pareamento arbitrário ao modelo. Primeiro, foi retomada a linha de base do responder às tentativas de pareamento ao modelo com estímulos usados em estudos anteriores. Após essa retomada, foram realizados testes utilizando os estímulos das duas classes funcionais estabelecidas previamente: nesses testes o S+ na tentativa (ex. uma letra) era um estímulo comparação da mesma classe do estímulo modelo (ex. outra letra) e o S- era um estímulo da outra classe (ex. um número). Ambos os sujeitos responderam de acordo com as classes previamente estabelecidas. Uma vez verificado esse repertório, foram realizados testes de emergência das relações de equivalência, nos quais ambos os sujeitos passaram.
O Experimento 3 testou se estímulos inseridos nas classes de equivalência estabelecidas no Experimento 2 funcionariam como membros das classes de estímulos funcionais estabelecidas no Experimento 1, ao inseri-los nos treinos de mudanças sucessivas de descriminações simples com estímulos originais das classes. Primeiro os novos estímulos foram acrescentados às classes (anteriormente treinadas em reversões de discriminação simples) por meio de um treino de pareamento arbitrário entre eles e os estímulos das classes nas quais eles seriam inseridos. Depois esses estímulos novos foram usados, juntamente com estímulos originais das duas classes funcionais, em treinos de mudanças sucessivas de descriminações simples. Ambos os sujeitos mostraram evidências da inclusão dos novos estímulos nas classes previamente estabelecidas, com 90% de acertos nas sessões de teste.
Em uma primeira análise, os resultados dos estudos de Sidman et al. (1989) e Kastak et al. (2001) oferecem suporte à proposição de Vaughan (1988) de que classes funcionais e classes de equivalência podem ser vistas como um mesmo fenômeno. No entanto, certos aspectos desses dois estudos obrigam a relativização de seus resultados. No estudo de Sidman et al. um dos participantes mostrou evidências de formação de classes funcionais sem emergência das relações de equivalência. Além disso, não pode ser descartado o efeito do repertório verbal dos participantes (ainda que comprometido) sobre a formação dos repertórios investigados. Já no estudo de Kastak et al. ambos os sujeitos já tinham uma longa história de treinos de pareamento arbitrário e testes de relações de equivalência, o que pode ter facilitado os testes desses repertórios no estudo. Portanto, os estudos realizados até o presente momento ainda não são conclusivos sobre a relação entre classes funcionais e classes de equivalência.
Sidman (2000) afi rmou que a estrutura proposta por ele para verificar a formação de equivalência de estímulos (treinos de pareamentos arbitrários com testes de relações emergentes) poderia, de fato, ser somente uma das possibilidades para se verificar o fenômeno. Sendo, portanto, procedimentos como o realizado por Vaughan (1988) uma das outras possibilidades. Por exemplo, ao se reportar ao conceito de partição explicitado por Vaughan, Sidman defende que “relações de equivalência consistem em pares ordenados de todos os elementos positivamente relacionados que participam de uma contingência” (p.145).
Consoante com essa mudança na forma de definir, interpretar e investigar a formação de classes de estímulos, recentemente tem-se buscado avaliar a aquisição desse repertório em situações que procuram se aproximar mais das situações naturais nas quais possivelmente a formação de classes ocorre (Costa, 2008). Essa aproximação às condições naturais de aquisição de classes procura aumentar a validade ecológica do estudo desse fenômeno e também evitar possíveis problemas de procedimento que possam estar dificultando encontrar mais abundantemente esse fenômeno em não humanos.
Com objetivo de simular respostas de forrageamento (procura por alimento) em cativeiro, Costa (2008) expôs dois macacos-prego (Cebus apella) a um treino de mudanças sucessivas de discriminações simples. Caixas de madeira, divididas arbitrariamente em dois conjuntos, eram apresentadas aos sujeitos em posições fixas em uma estante (as caixas se diferenciavam pela cor e por uma fruta de cera fixada na parte superior). Procurar por alimento nas caixas pertencentes ao grupo designado como S+ em uma sessão,era consequenciado por achar um tipo de alimento em seu interior. Caso o sujeito, durante uma tentativa, procurasse por alimento somente nas caixas S+ e se afastasse das demais caixas, sem procurar nas caixas com função de S-, mais um pedaço de alimento era dispensado como bônus. Procurar por alimento em qualquer caixa designada como S- encerrava a tentativa sem o bônus. Uma vez que o critério de aprendizagem fosse alcançado, as funções dos estímulos eram revertidas.
As sessões experimentais se iniciaram com a apresentação de duas caixas, uma de cada grupo de estímulos. Posteriormente, sessões com quatro e seis caixas foram realizadas. Por fim, testes de formação de classes foram realizados. Nestes testes, quatro das seis caixas experimentais eram inicialmente apresentadas aos sujeitos com contingências revertidas se comparadas à sessão anterior. Quando o critério de aprendizagem era alcançado, as demais caixas eram reintroduzidas no contexto experimental e a primeira resposta era observada. Responder nas caixas reintroduzidas de forma condizente com a contingência em vigor era considerado pelos autores indicio de formação de classes. Seis testes foram realizados com diferentes caixas. Os dois sujeitos apresentaram respostas condizentes com a formação de classes em todos os seis testes.
A metodologia utilizada no estudo de Costa (2008) mostrou-se suficiente para treinar classes funcionais. Acredita-se que o treino e teste utilizados naquele estudo tenha sido uma maneira eficaz de avaliar a formação de classes, sem se ater ao paradigma de equivalência que exigiria testes de reflexividade, simetria e transitividade.
O presente trabalho visa realizar um procedimento semelhante ao de Costa (2008), utilizando como sujeitos cães domésticos (Canis familiaris), considerando os indícios recentes da possibilidade e das vantagens de utilizar esta espécie como um modelo animal em estudos sobre comportamentos humanos (e.g. Kaminski, Call, & Fischer, 2004; Range, Aust, Steurer, & Huber, 2008; Rossi & Ades, 2008; Udell & Wynne, 2008; Wobber, 2005).
Udell e Wynne (2008) apresentam uma revisão das linhas de pesquisa sobre os repertórios comportamentais dos cães. Eles apontam que grande parte dos estudos é realizada sob perspectivas etológicas e da psicologia do desenvolvimento. Esses estudos buscam verificar a existência de processos psicológicos semelhantes entre cães e humanos, tendo em vista o ambiente e as pressões seletivas que as duas espécies dividem há milhares de anos. Os autores concluem que os analistas do comportamento deveriam incluir mais frequentemente essa espécie como sujeito de suas pesquisas, visto que em determinadas tarefas os cães apresentam desempenhos superiores aos primatas não-humanos tradicionalmente utilizados como modelos animais do comportamento humano (com a vantagem adicional, não mencionada por eles, de que conseguir e manter os sujeitos para a pesquisa é bem menos dispendioso e mais fácil em comparação com a obtenção e manutenção de primatas não-humanos – ver Range et al., 2008)
Outros autores também apontam as vantagens na utilização dessa espécie em estudos sobre comportamentos simbólicos e pré-simbólicos. Tem sido discutido, por exemplo, que a evolução da espécie canina junto ao homem resultou em habilidades sociais e comunicativas que podem favorecer o estudo comparativo das duas espécies (Cooper, Ashton, Bishop, West, Mills, & Young, 2003; Wobber, 2005). Tem-se reportado também que cães resolvem tarefas envolvendo gestos comunicativos humanos mais facilmente que primatas não-humanos (e.g. Soproni, Miklósi, Csányi, & Topál, 2001).
Mais recentemente, estudos têm verificado em cães domésticos a aprendizagem por exclusão em uma tarefa de discriminação auditivo-visual – buscar objetos em um quarto após a sua nomeação (Kaminski et al., 2004), evidências da produção de respostas “comunicativas” por meio de lexigramas específicos colados em um teclado (Rossi & Ades, 2008) e a categorização de estímulos visuais apresentados na tela de um computador - 40 imagens de cães (S+) e 40 de paisagens (S-) (Range et al., 2008). Esses resultados indicam que essa espécie pode fornecer um novo modelo animal para estudar comportamento simbólico e pré-simbólico, com as vantagens já apontadas.
Portanto, o presente estudo objetivou expor dois indivíduos da espécie Canis familiaris a um procedimento que visou estabelecer a formação de duas classes funcionais através de um treino de mudanças sucessivas de discriminação simples, possibilitando: 1) uma avaliação do uso da espécie como modelo animal em estudos sobre formação de classes funcionais (um repertório pré-simbólico); e 2) uma comparação com o estudo de Costa (2008).
MÉTODO
Sujeitos: participaram do estudo dois cães domésticos (Canis familiaris): Lucke, um macho da raça Daschund, mestiço, com um ano e 8 meses de idade no início do experimento, e Yumi, uma fêmea da raça Yorkshire, com 6 meses no início do experimento. Lucke vive com a experimentadora e sua família, e Yumi com outra família. No início do estudo, Lucke já havia passado por treinos de reversões sucessivas de discriminação simples com estímulos visuais, enquanto Yumi era experimentalmente ingênua. Os cães eram levados diariamente para a Universidade Federal do Pará para a realização do experimento e trazidos de volta para suas casas pela experimentadora. Os sujeitos eram alimentados em suas casas no máximo até as 23 horas do dia anterior à sessão experimental, realizada entre 9:00 e 12:00 horas.
Ambiente e Equipamentos: os dados foram coletados em uma sala (5,0 X 5,50 m) do Laboratório de Psicologia Experimental da UFPA, climatizada e com iluminação artificial. Para a realização das sessões experimentais, foram usadas seis caixas de madeira (0.40 x 0.40 x 0.40 m) revestidas com EVA. Os lados frontais e posteriores das caixas tinham aberturas de 40 cm, cobertas com uma cortina de napa preta, que impedia a visão do conteúdo das caixas, mas permitia o acesso ao interior das mesmas. As caixas se diferenciavam pela cor, pela presença de um brinquedo infl ável diferente acima de cada uma e por formas (feitas em EVA) pregadas na parte superior do lado frontal das caixas (ver Figura 1). Uma câmera de vídeo foi usada para registrar algumas sessões de treino e os testes, e folhas de registro foram usadas para anotar as respostas dos sujeitos. Uma coleira foi utilizada para controlar a movimentação e permitir o acesso dos sujeitos às caixas, e para fazê-los retornar ao ponto inicial após cada tentativa.
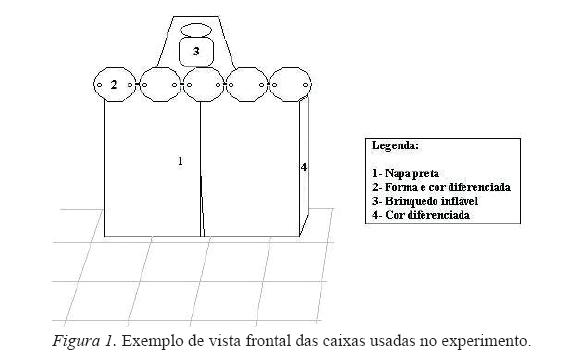
ESTÍMULOS
Discriminativos: foram utilizadas as seis caixas de madeira descritas no item “Ambiente e Equipamentos”. Elas foram divididas arbitrariamente em dois conjuntos: Conjunto 1 (A1, B1 e C1) e Conjunto 2 (A2, B2 e C2).
Conseqüências: após um teste de preferência com diversos itens alimentícios, foram estabelecidos dois itens para cada sujeito, os quais serviram como conseqüências específicas para respostas às caixas do Conjunto 1 (R1) e do Conjunto 2 (R2). O R2, para ambos os sujeitos, foi biscoito para cachorro (da marca Purina), enquanto os R1 de Lucke e de Yumi foram, respectivamente, copa e snacks para cães (tipo “bifinho”, da marca Purina).
PROCEDIMENTO
Considerando os objetivos de avaliar a formação de classes funcionais em arranjos experimentais com maior validade ecológica, e de comparar os resultados com os do estudo de Costa (2008), foi elaborada uma tarefa de discriminação simples com mudanças sucessivas nas funções dos estímulos (DSMS), semelhantes à daquele estudo, e que simula uma situação de busca por alimento (forrageio).
A tarefa consistiu em uma situação na qual duas, quatro ou seis caixas (dependendo da fase do experimento – ver descrição das fases a seguir) foram colocadas na sala, sendo inseridos dentro de algumas delas pedaços dos itens alimentícios correspondentes ao conjunto de cada caixa.
No início de cada tentativa de qualquer fase, o sujeito era mantido por um experimentador (E1) a uma distância de dois metros das caixas. As caixas ficavam dispostas no chão da sala, lado a lado (quando adjacentes, distantes 20 cm uma da outra), mantendo sempre a mesma posição na sala em todas as fases (ver Figura 2). Ou seja, em uma sessão na qual determinada caixa não estava presente, essa era retirada de sua posição, e as outras permaneciam em suas posições fixas.
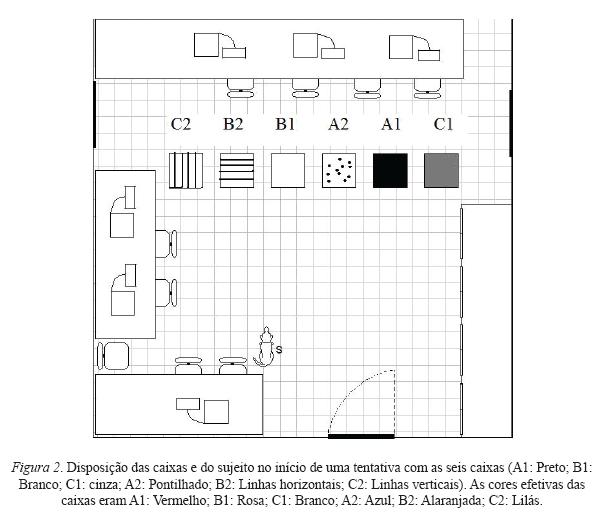
No início de cada tentativa, um segundo experimentador (E2), que permanecia próximo ao E1 durante o responder do sujeito, caminhava até a parte de trás das caixas e introduzia a mão em cada uma delas. O experimentador simulava colocar um item alimentício em todas, mas efetivamente colocava apenas o item específico que correspondesse ao conjunto da(s) caixa(s) designada(s) com a função de estímulo(s) positivo (S+) naquela sessão. Dentro das caixas do Conjunto 1 era sempre colocado um pedaço de alimento R1, e das caixas do Conjunto 2 um do alimento R2. Quando o E2 retornava para diante das caixas, E1 afrouxava a guia, permitindo que o sujeito se movimentasse pela sala.
Foi considerada uma resposta de escolha introduzir a cabeça ou o focinho no interior de uma caixa. Ao responder em uma caixa com função positiva (S+), encontrando o alimento específico do conjunto ao qual pertencia aquela caixa, o sujeito permanecia solto na sala, podendo responder a outra caixa. Uma tentativa correta consistia em responder a todas as caixas com função positiva e retornar ao E1. Quando isso ocorria, o sujeito recebia um pedaço de alimento bônus, específico do Conjunto S+ na sessão. Quando o sujeito respondia em uma caixa S-, ele era puxado de forma suave pela guia (por E1) para que se afastasse das caixas, indicando o fim da tentativa. Concomitantemente à resposta no S-, as luzes da sala eram desligadas, iniciando um time-out de 15 segundos.
Portanto, uma tentativa iniciava quando o E2 já havia disposto os alimentos nas caixas, sendo a guia presa à coleira do sujeito liberada. A tentativa terminava quando o sujeito respondia a todas as caixas corretas, uma a pós a outra (em qualquer ordem) e retornava ao E1 (tentativa correta, seguida por liberação do bônus) ou quando respondia em uma errada, com função negativa, mesmo que tenha respondido em alguma caixa com função positiva anteriormente (tentativa incorreta, seguida de time-out). Se o sujeito respondesse em todas as corretas, mas não retornasse ao E1, permanecia solto na sala até que respondesse em uma caixa com função negativa (o que seria consequenciado com time-out). Outras respostas às caixas com função positiva não tinham conseqüência programada.
Após o consumo do bônus ou após o time-out, era iniciada uma nova tentativa. Foram realizadas de uma a três sessões diárias, cinco vezes por semana com cada sujeito. Quando havia mais de uma sessão no dia, era realizado um intervalo entre elas de cerca de dez minutos, durante o qual o cão era retirado da sala.
Uma sessão consistia em um total de 20 tentativas, sendo que se o sujeito alcançasse o critério de seis tentativas corretas consecutivas, a sessão era finalizada antes do término de 20 tentativas. Na sessão seguinte, sempre realizada um dia após o alcance de critério, a função dos estímulos era modificada.
Durante todas as sessões, o E2 utilizava óculos escuros para reduzir o risco de apresentação de dicas quanto à classe de estímulos com função positiva aos sujeitos quando o alimento era colocado nas caixas. Foram também tomadas três medidas para evitar que o comportamento de seleção da caixa ficasse sob controle do cheiro do alimento na mesma. Antes do início da sessão, era colocado um pedaço de alimento (referente à classe em vigor na sessão): 1) também nas caixas com função negativa, mas retirados logo em seguida; 2) acima de cada caixa (à frente dos brinquedos infl áveis); e (3) em um pedaço de pano anexado dentro de cada caixa, no tecido de napa que cobria a caixa).
Além disso, quando o critério de término das sessões (de seis tentativas corretas consecutivas) era alcançado nas primeiras dez tentativas de uma sessão, duas conseqüências eram apresentadas: o sujeito recebia pedaços extras de alimentos e passeava com os experimentadores na área externa do Laboratório de Psicologia Experimental. Dessa maneira, evitava-se a maximização de reforços pelo sujeito através de erros na sexta tentativa após cinco tentativas respondidas corretamente de maneira consecutiva. Esse procedimento foi realizado após se perceber que Lucke, em um estudo prévio, errava consistentemente na sexta tentativa após ter realizado cinco tentativas corretas consecutivas. Esse padrão foi também verificado em macacos-prego (Barros, Picanço, Costa, & Souza, em preparação). Ou seja, o sujeito com freqüência acertava cinco tentativas e errava a sexta, o que levou os pesquisadores à análise de que se o sujeito errasse na quinta tentativa, ele tinha a possibilidade de continuar a sessão, e assim, receber mais alimento. A inclusão do passeio e dos pedaços extras de comida funcionou como conseqüência reforçadora adicional para o acerto das seis tentativas corretas consecutivas.
O comportamento dos sujeitos de buscar os pedaços de alimentos nas caixas, introduzindo a cabeça dentro delas, foi modelado. Nas primeiras sessões, foram usadas dicas auditivas (assobios e/ou chamar o sujeito pelo nome), emitidas pelos experimentadores (E1 ou E2), procurando garantir que o sujeito retornasse para os experimentadores sem responder na(s) caixa(s) com função negativa (S-s) (para maiores detalhes do procedimento de modelagem realizado ver Dahás, 2009).
Em todas as fases, foi considerado como critério para que uma mudança de função dos estímulos fosse realizada o responder correto em seis tentativas consecutivas.
Fases do Procedimento: após a modelagem, foram realizadas sete Fases (ver Tabela 1)
- Fase 1- Treino de MSDS com duas caixas por sessão, até que quatro (A1, B1, A2 e B2) das seis caixas tivessem funcionado uma vez como estímulo positivo (S+) e outra como estímulo negativo (S-). O treino ocorreu de maneira que o S+ do par que estava sendo treinado não aparecia como S- após a mudança na função dos estímulos (procurando evitar um efeito de persistência comportamental – ver Rico, 2006). O estímulo anteriormente negativo adquiriu a função positiva com a mudança. As sessões tinham 20 tentativas, e o critério para realizar a mudança na função dos estímulos era de seis tentativas executadas corretamente de maneira consecutiva em uma sessão (sem considerar as respostas nas quais foram usadas dicas auditivas após a resposta ao S+).
- Fase 2- Treino de MSDS com as quatro caixas da Fase 1 (A1, B1, A2 e B2) apresentadas juntas na sessão. Inicialmente duas caixas de um conjunto eram designadas como estímulos positivos (S+) e as outras duas do outro conjunto como S-(s). As sessões tinham 20 tentativas e o critério para realizar a mudança na função dos estímulos era de seis tentativas executadas corretamente de maneira consecutiva em uma sessão (desconsiderando respostas com dicas). A resposta correta em uma sessão era a de responder nas duas caixas designadas como S+ (em qualquer ordem) e retornar para receber o bônus. Responder em uma caixa S+ (ou nas duas) e, em seguida, em uma caixa S-, ou responder primeiro em uma S- foi considerado erro, levando ao time-out e término da tentativa. Esse procedimento foi mantido até se obter o critério de aprendizagem (seis tentativas corretas consecutivas) em seis sessões consecutivas. Nesse caso, seis reversões seriam efetuadas em seis sessões consecutivas.
- Fase 3- História de reforçamento de respostas às duas caixas ainda não utilizadas (C1 e C2). Realizou-se uma sessão na qual as caixas C1 e C2 eram colocadas em suas posições na sala. Foram então contabilizadas 10 vezes nas quais os sujeitos, ao responder na Caixa C1, encontraram um alimento dentro dela (diferente dos específicos de cada Conjunto). O responder na caixa C2 não tinha conseqüência programada até então. Após encontrar alimento por 10 vezes na caixa C1, a condição foi revertida: os sujeitos encontraram o mesmo alimento na caixa C2. Dessa vez, respostas à caixa C1 não tinham conseqüência programada.
-Fase 4- Treino de discriminação simples com as seis caixas (A1/B1/C1 como Se A2/B2/C2 como S+). Foram realizadas sessões de 20 tentativas até que o critério de aprendizagem (seis tentativas consecutivas executadas corretamente) fosse alcançado. Nessa Fase, uma resposta correta foi o responder consecutivo (em qualquer ordem) nas três caixas designadas como S+ e retornar para receber o bônus. Uma resposta intermediária (ou inicial) em um S- foi considerada erro e a tentativa foi encerrada.
- Fase 5- Mudança da função no treino de discriminação das caixas na Fase 4 e Teste da Formação de Classes. Foram realizadas sessões de 20 tentativas que começavam somente com quatro caixas (A1/B1[S+] e A2/B2[S-]), com as funções revertidas em relação à Fase 4. Quando o sujeito alcançou o critério de aprendizagem (seis tentativas executadas corretamente) ele foi retirado da sala e as caixas C1(S+) e C2(S-) foram colocadas em seus lugares junto com as outras quatro. Após a inserção das duas caixas o sujeito foi trazido de volta para a sala e uma nova sessão com o Teste de Formação de Classes foi iniciado. Para ser aprovado no teste, o sujeito deveria ter respondido em seqüência às três caixas do Conjunto relacionado com o alimento após a mudança da função dos estímulos, sem responder em nenhuma caixa S- e retornando ao E1 para receber o bônus. Como o responder correto a todas as tentativas foi reforçado, somente a primeira tentativa após a inserção das caixas C1 e C2 tem validade estrita de teste6. O critério de seis tentativas consecutivas executadas corretamente ou o término da sessão de teste (com 20 tentativas) determinou o encerramento da Fase 5.
-Fase 6 – Caso não fossem encontradas evidências de formação de classes no primeiro teste da Fase 5, seria realizado um treino para avaliar a formação de classes funcionais durante o treino de reversões sucessivas das funções dos estímulos utilizando as seis caixas em cada sessão. Inicialmente três caixas de um conjunto seriam designadas como (S+) e as outras três do outro conjunto como S-. As sessões constituir-se-iam de 20 tentativas e o critério para realizar a mudança na função dos estímulos seria de seis tentativas consecutivas executadas corretamente em uma sessão (desconsiderando as respostas com dicas). A resposta correta em uma sessão seria responder nas três caixas designadas como S+ (em qualquer ordem) e retornar para receber o bônus. Uma resposta intermediária (ou inicial) em um S- seria considerada erro, e a tentativa seria encerrada. Esse procedimento foi mantido até que se observasse a ocorrência do critério de aprendizagem de seis tentativas consecutivas executadas corretamente nas dez primeiras tentativas em 11 de 12 sessões consecutivas.
- Fase 7 – Nessa fase, o teste da Fase 5, com a retirada inicial dos estímulos C1 e C2 e posterior inserção dos mesmos foi novamente realizado. Após a repetição do primeiro teste, foram realizados outros cinco testes, de maneira que todos os pares de estímulos (A1A2, B1B2 e C1C2) foram retirados duas vezes (para que cada caixa passasse por testes com a função tanto de S+ como de S-). Após cada sessão de teste, realizou-se uma mudança na função dos estímulos. Assim, os Testes 1, 3 e 5 ocorreram com os estímulos do Conjunto 1 como S+ e os testes 2, 4 e 6 com os estímulos do Conjunto 2 como S+ (ver Figura 3). O critério de seis tentativas consecutivas executadas corretamente ou o término da sessão de teste (com 20 tentativas) determinou o encerramento da Fase 7.
RESULTADOS
A Tabela 1 mostra o número de mudanças das funções dos estímulos, sessões e tentativas realizadas com cada classe de estímulos nas Fases 1, 2, 4 e 6, para cada sujeito. No total foram realizadas 91 sessões de treino com Lucke e 144 sessões com Yumi. Essa diferença de número total de sessões se deve à Fase 2, na qual Lucke necessitou 545 tentativas para alcançar o critério de mudança de fase, enquanto Yumi necessitou mais de 1.300 tentativas, totalizando 106 sessões. Essa discrepância se deve, provavelmente, ao fato de Lucke ter sido previamente exposto a um procedimento de mudanças sucessivas de discriminação simples.

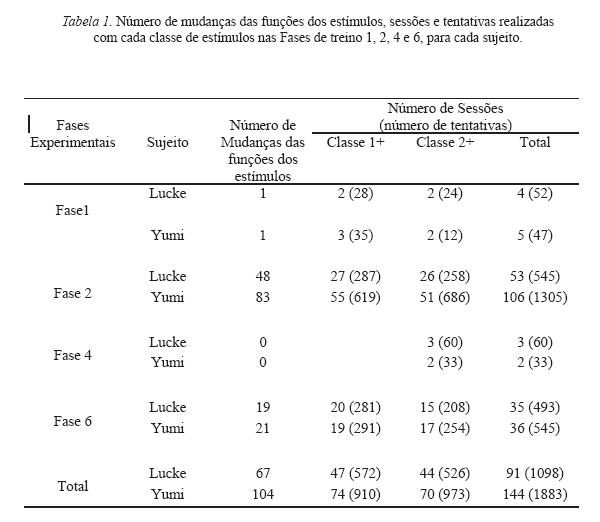
Na Tabela 1 pode-se observar uma diminuição no número de sessões/tentativas necessárias para a finalização da Fase 2 (com quatro caixas) para a Fase 6 (com seis caixas), indicando um efeito de learning set para a tarefa de discriminações simples com mudanças sucessivas nas funções dos estímulos. Ao mesmo tempo, o número de sessões/ tentativas realizadas com a Classe 1 e com a Classe 2 como positiva foi semelhante em todas as fases de treino. Isto indica que não havia uma classe de estímulos que dificultasse o responder discriminado mais do que a outra.
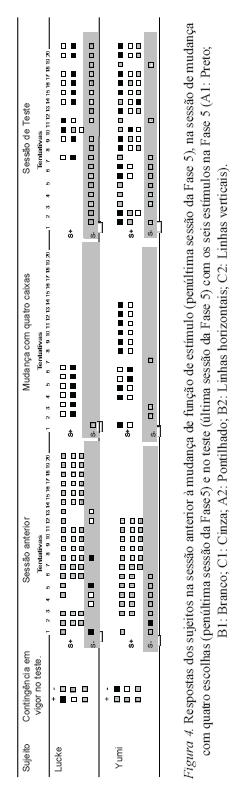
A Figura 4 mostra os dados das sessões dos testes de formação de classes (Fase 5), além da sessão imediatamente anterior à retirada das caixas e da imediatamente anterior aos testes. Ambos os sujeitos responderam adequadamente à mudança de função, como mostrado na quarta coluna da Figura 4. No entanto, as primeiras seis tentativas de teste (quinta coluna) não indicam formação de classe. Lucke respondeu às seis primeiras tentativas do teste erroneamente: nas caixas A2 e B2. Dentre as seis primeiras tentativas do teste, Yumi respondeu corretamente a segunda. Já nas tentativas 1 e 6 respondeu adequadamente em duas caixas corretas, mas não na terceira, e errou logo na segunda caixa nas tentativas 3, 4 e 5.
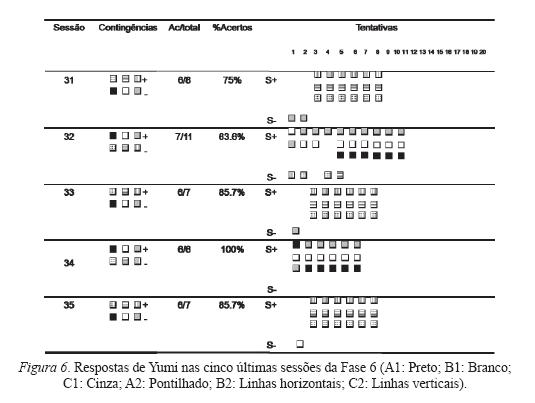
Os quadrados da Figura 4 indicam (conforme legenda) a caixa na qual o sujeito respondeu, e uma linha vertical de caixas indica, de cima para baixo, a ordem na qual o sujeito respondeu às caixas. Por exemplo, na terceira coluna da fi gura, Lucke respondeu as seis tentativas na mesma ordem: C2 (linhas verticais), A2 (pontilhado), B2 (linhas horizontais). As Figuras 5 a 8 devem ser lidas da mesma forma.
Os dados da Figura 4 apontam que o procedimento de mudança sucessivas nas funções dos estímulos com duas e quatro caixas não foi suficiente para estabelecer um responder adequado às classes de seis caixas.
As Figuras 5 e 6 apresentam as respostas de Lucke e Yumi, respectivamente, às últimas cinco sessões da Fase 6. Nota-se que os sujeitos apresentam poucos erros, concentrados nas primeiras tentativas. Vaughan (1988) defende que esse padrão de erro inicial e acerto consistente posterior seria um indício de formação de classes funcionais ou de equivalência. Inicialmente o sujeito responderia aleatoriamente, mas ao entrar em contato com a contingência em vigor naquela sessão (respondendo na caixa com função de S-) o sujeito passa rapidamente a responder adequadamente.
As Figuras 7 e 8 mostram os dados das sessões de testes de formação de classes realizados após o treino de mudança de função de estímulos com seis caixas, além da sessão imediatamente anterior à retirada das caixas e da imediatamente anterior ao teste (todas com, no máximo, 20 tentativas).
A Figura 7 mostra os dados dos testes realizados com Lucke. Em dois dos seis testes, o responder do sujeito foi adequado às contingências em vigor: testes C1 (100% de acerto) e A1 (erro somente na quinta tentativa). Os dois únicos testes que não necessitaram de sessões posteriores para que o sujeito alcançasse o critério de seis tentativas corretas consecutivas foram justamente aqueles com resultado positivo (C1 e A1). Além disso, ocorreram tentativas corretas entre as seis primeiras dos testes C2 (2, 3, 6), B1 (3, 5) e B2 (3, 4, 6). O teste B1 foi o realizado com mais erros por Lucke.
Algumas variáveis não controladas pelo experimentador afetaram o responder do sujeito durante as sessões de teste. Três sessões de teste foram canceladas, pois o sujeito logo no início da sessão não se dirigia às caixas. A primeira vez na qual isso ocorreu foi em uma sessão após o teste C2. Lucke não comia os pedaços de biscoito deixados dentro das caixas e ficou 10 minutos sem responder. Na segunda sessão, prévia ao teste A1, Lucke cheirava o local do chão onde Yumi tinha estado. Foi então verificado que Yumi estava no cio, e a partir de então, ela não foi mais levada ao laboratório até que a coleta com Lucke fosse finalizada. A terceira sessão cancelada foi uma sessão de mudança com quatro caixas prévia ao teste A2, na qual Lucke chorava e continuava cheirando o mesmo local do chão, assim como a calça jeans utilizada pelo E2 quando carregara Yumi no dia anterior.

Além disso, durante o teste B1, foi verificado que alguns pedaços de copa (Ref 1) não foram consumidos pelo sujeito. Na sessão seguinte, Lucke novamente não se dirigia às caixas. A copa foi então trocada pelo bifinho, Ref 1 utilizado inicialmente somente com Yumi, e assim, Lucke passou a responder adequadamente.
A Figura 8 mostra os dados dos testes realizados com Yumi. Em três dos seis testes, o responder do sujeito foi adequado às contingências em vigor: testes C1 (erro somente na primeira tentativa), C2 (erro somente na primeira tentativa) e A1 (erro somente na sexta tentativa).
Nota-se que os testes A2 e B1, que não apresentam resultados positivos diretos (nas seis primeiras tentativas), apresentaram poucos erros. No teste A2, Yumi respondeu adequadamente nas três primeiras tentativas, mas errou nas outras três. Em seguida, acertou mais cinco tentativas, errou outra e acertou seis, sendo assim encerrada a sessão.
No teste B1, Yumi acertou as tentativas 1, 2 e 4, mas errou as 3, 4 e 6. O restante da sessão mostra que o sujeito ainda respondeu mais quatro vezes na caixa A2 (S-), mas no restante das tentativas, respondeu corretamente. Yumi alcançou critério para mudança de função na sétima tentativa da sessão seguinte.
Ocorreram tentativas corretas entre as seis primeiras do teste B2 (3, 4, 6). No entanto, essa foi a sessão de teste com mais erros realizada por Yumi. Apenas os testes B1 e B2 necessitaram de sessões posteriores para que o sujeito alcançasse o critério de seis tentativas corretas consecutivas.
O responder dos sujeitos no decorrer do primeiro teste da Fase 7 foi consistente com as contingências em vigor, diferentemente do resultado do teste da Fase 5. Pode-se afi rmar que o treino de mudanças sucessivas de discriminação simples com seis caixas criou condições adequadas para que os sujeitos respondessem aos testes corretamente. No entanto, essas condições não foram suficientes para manter o responder adequado no decorrer de todos os testes da Fase 7.
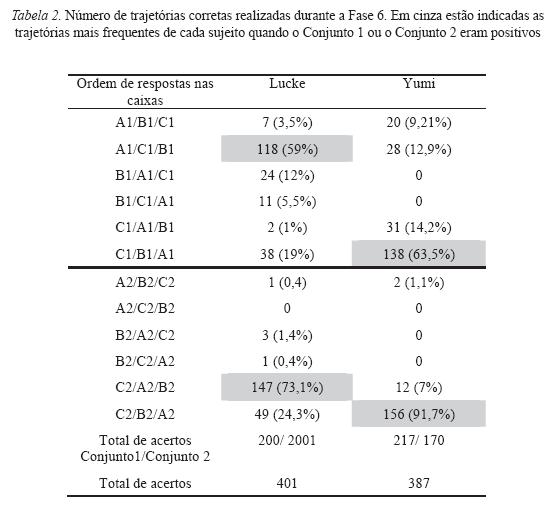
Durante a coleta de dados, o padrão de resposta dos sujeitos sugeria que eles poderiam estar respondendo sob controle da posição das caixas no chão, ou ainda, sob controle da trajetória percorrida de uma caixa para outra. Os dados da Tabela 2, referentes à Fase 6, mostram que os sujeitos respondiam mais seguindo determinadas trajetórias do que a outras. As marcações em cinza indicam as trajetórias mais frequentes de cada sujeito quando o Conjunto 1 ou o Conjunto 2 eram positivos: Lucke obteve 59% dos seus acertos quando o Conjunto 1 era positivo com a trajetória A1C1B1, e 73,1% com a trajetória C2A2B2 quando a Conjunto 2 era positivo. Yumi obteve 63, 5% dos seus acertos nas sessões com o Conjunto 1 como positivo com a trajetória C1B1A1, e 91,7% nas sessões com o Conjunto 2 como positivo com a trajetória C2B2A2.


Visto que os sujeitos respondiam às caixas repetindo um número limitado de trajetórias possíveis, resolveu-se analisar se os erros ocorridos durante as sessões de teste na Fase 7 estariam relacionados com a trajetória realizada na sessão anterior (com quatro caixas). Dessa forma, poder-se-iam explicar os erros realizados nos testes através do controle que a trajetória da sessão anterior exercia sobre o responder dos sujeitos.
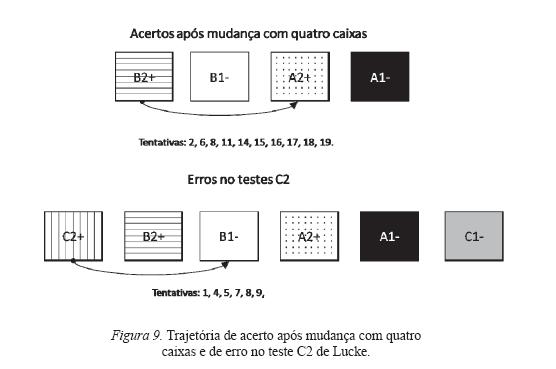
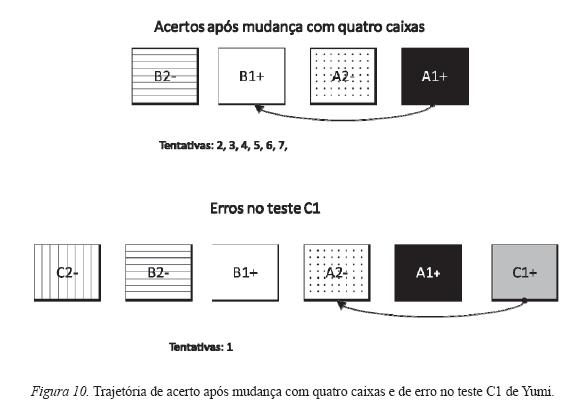
As Figuras 9 e 10 são exemplos desse controle pela trajetória. Elas mostram os resultados das trajetórias de acerto mais freqüentes nas sessões com quatro caixas e de erro mais freqüentes nas sessões de teste, de maneira que se averiguou a repetição do padrão das trajetórias realizadas em uma sessão de mudança com quatro caixas prévia ao teste e das realizadas no teste. Somente são apresentados dois exemplos de teste com cada sujeito.
A Figura 9 apresenta as trajetórias corretas realizadas por Lucke na sessão com quatro caixas anterior ao teste e as trajetórias erradas na sessão do teste C2. As caixas estão dispostas na posição em que ficavam durante a sessão: a seta indica a trajetória do sujeito: a ponta com um círculo preto indica a primeira caixa na qual o sujeito respondeu, e com um triângulo, a última caixa. Nota-se que, no teste, o sujeito realizou a mesma trajetória (responder na primeira e na terceira caixas da esquerda para a direita) que havia realizado durante a sessão prévia com quatro caixas, o que produziu seis erros.
A Figura 10 mostra as trajetórias corretas realizadas por Yumi na sessão com quatro caixas anterior ao teste e as erradas na sessão do teste C1. Nota-se que o único erro no teste foi na primeira tentativa, na qual o sujeito realizou a mesma trajetória (responder na primeira e na terceira caixas da direita para a esquerda), que durante a sessão prévia com quatro caixas foi a única utilizada das possibilidades de trajetória correta.
Estes resultados de ambos os sujeitos indicam que eles responderam nas sessões de teste da Fase 7, ao menos em parte, sob controle de uma trajetória que na sessão anterior consistia em um responder correto.
DISCUSSÃO
Os sujeitos demonstraram ser capazes de responder discriminadamente aos estímulos (ou às suas posições) nas Fases 1, 2, 3 e 6. Estes resultados reforçam as evidências recentes sobre a viabilidade de se utilizarem cães como sujeitos em procedimentos geralmente aplicados ao estudo de repertórios simbólicos e pré-simbólicos em primatas (e.g. Kaminski et al., 2004; Range et al., 2008; Rossi & Ades, 2008; Udell & Wynne, 2008).
O estudo de Range et al. (2008) já havia demonstrado que cães eram capazes de responder discriminadamente a estímulos visuais, a responder a classes desses estímulos e ainda a incluir membros nunca antes treinados a essas classes. Os dados do presente experimento, apesar de menos conclusivos, confi rmam a possibilidade de se utilizar essa espécie em estudos acerca do comportamento pré-simbólico e simbólico, tendo em vista a alta probabilidade de a co-evolução das espécies humana e canina ter rendido semelhanças comportamentais importantes (Cooper et al., 2003; Wobber, 2005).
Outro estudo que aponta para a formação de classes com estímulos visuais em cães é o de Domeniconi, Bortoloti, Antoniazzi e Mendes (2008). Três cães foram treinados a responder a três pares de estímulos (A1A2, B1B2, C1C2) em um procedimento de reversões sucessivas de discriminação simples com reforços específicos para os elementos 1 e 2 nos pares. Os treinos eram realizados com um par por sessão, e os testes consistiam em sessões com os três pares apresentados em blocos ou aleatoriamente. Os sujeitos responderam adequadamente às primeiras tentativas com cada par de estímulos durante os testes, o que os autores sugerem ser um indício de formação de classes (apesar de sugerirem também que o odor do ítem reforçador específico presente na sala poderia servir como dica para o responder adequado).
De forma diferente do presente estudo, Domeniconi et al. (2008) apontam que não houve indício de learning set no desempenho de seus três sujeitos. À parte outras diferenças de procedimento (como número maior de estímulos no nosso estudo), a diferença que pode explicar o surgimento de learning set no presente estudo e não no de Domeniconi et al. pode ser o maior número de sessões de treino realizadas no estudo atual (os sujeitos de Domeniconi et al. realizaram entre 33 e 39 sessões de treino, contra as 91 sessões de Lucke e as 144 de Yumi).
Os dados dos treinos com seis caixas (Figuras 5 e 6) apontam que o responder dos sujeitos estava de acordo com o conceito de formação de classes funcionais de Vaughan (1988), já que, logo nas primeiras tentativas das sessões, o responder se adequava às contingências. No entanto, não há indícios fortes de formação de classes nos resultados dos testes de Lucke e Yumi. Lucke respondeu adequadamente a dois dos testes (testes C1 e A1) e Yumi a três deles (C1, C2 e A1). Como foi mencionado antes, a falta de controle de algumas variáveis como a saciação alimentar e o cio de Yumi parece ter alterado fortemente o responder de Lucke durante a fase de teste.
Costa (2008) propõe que, em um procedimento como o do presente experimento, pode-se aceitar como indício de formação de classes o fato de que, durante um teste, o primeiro responder em uma das caixas positivas é seguido pelo responder adequado nas outras caixas pertencentes à classe da primeira. De acordo com esse critério de precisão para aceitar resultados de testes como evidência de classes funcionais, pode-se dizer que há indícios de formação de classes somente em um dos 6 testes realizados por Lucke (teste A1) e em quatro dos testes realizados por Yumi (testes C1, A1, A2, B1).
Comparando os resultados do presente estudo com os do primeiro experimento de Costa (2008), verifica-se que os dois macacos-prego apresentaram responder mais consistente com as classes de estímulos do que os dois cães do presente experimento. Na fase equivalente à Fase 6 do presente experimento, um dos sujeitos respondeu corretamente a 100% dos seis testes, e outro sujeito efetuou somente um erro na quinta tentativa do terceiro teste. Essa diferença se deu mesmo considerando que os sujeitos participaram de menos sessões de treino do que os cães do presente experimento. Enquanto Lucke e Yumi participaram de 35 e 36 sessões na Fase 4, respectivamente, os dois sujeitos de Costa participaram de 12 e 11 sessões na fase equivalente. Esse aparente déficit dos sujeitos do presente experimento, quando comparados com os macacos-prego do experimento de Costa, pode se dever a diferenças perceptuais inter-específicas discutidas mais adiante.
No entanto, diferenças metodológicas dos dois estudos e/ou características das espécies utilizadas podem explicar os resultados. O custo do responder às caixas no estudo de Costa (2008) pode ser maior do que no presente estudo, visto que naquele estudo os sujeitos precisavam subir ou descer de uma caixa a outra, escalando a parede da gaiolaviveiro (2,5 m de altura), enquanto neste estudo as caixas permaneciam mais próximas e no mesmo plano. Essa característica da apresentação dos estímulos pode ter facilitado o responder persistente em caixas com função negativa mesmo após o responder correto nas caixas com função positiva, emitidos pelos sujeitos do presente estudo. O uso dos mesmos reforçadores por longo período de coleta causou saciação em um dos sujeitos do segundo estudo, o que não foi relatado por Costa. Por último, os sujeitos do presente estudo responderam mais sob controle da trajetória do que das características visuais dos estímulos, enquanto um dos sujeitos de Costa respondeu adequadamente às classes mesmo no Experimento 2, no qual as caixas mudavam de lugar a cada sessão.
O responder inadequado em alguns dos testes pode também se dever a algumas características do planejamento experimental do atual estudo. Primeiro, visto que a cada alcance de critério a função das caixas era mudada, a situação de teste era diferenciada de todo o treino, não havendo mudança de função em relação à sessão anterior. Ou seja, na sessão que ocorria antes do teste, com quatro caixas, se a Conjunto 1 era positivo, a sessão de teste teria o mesmo conjunto como positivo. Talvez melhores resultados fossem adquiridos planejando-se manter a mesma função dos estímulos por mais de uma sessão, mesmo com alcance de critério, ao menos aleatoriamente. Dessa maneira, a situação de teste não seria tão diferenciada da de treino.
Segundo, a posição relativa das caixas não permanecia a mesma nas sessões de mudança com quatro caixas prévia ao teste e de teste. Por exemplo, a primeira caixa da esquerda para a direita era a B2 na sessão com quatro caixas antes do teste C1, enquanto era a C1 no teste C1. Um procedimento no qual a posição das caixas mudasse a cada sessão ou tentativa poderia minimizar esse problema. Estando sempre as caixas em uma nova posição, o responder sob controle das características visuais da caixa seria mais provável do que sob controle da posição dela na sala.
Terceiro, não foram controlados, no presente experimento, os odores deixados pelos sujeitos na sala experimental. Assim, quando Yumi entrou no cio, o responder de Lucke foi alterado. Essa variável poderia ser controlada através do uso de salas diferenciadas para machos e fêmeas, da limpeza da sala com produtos específicos para retirada de odores orgânicos, ou ao menos não se realizando sessões com a fêmea durante o cio.
Quarto, o uso repetido dos mesmos itens alimentícios fez com que Lucke parasse de responder em diversas sessões. Um experimento posterior poderia alternar itens semanalmente, ou fazer diversos testes de preferência no decorrer do experimento.
Por último, e mais importante, sabe-se que a visão canina não é tão apurada quanto a audição e a olfação. Há dados indicando que a visão, sendo apropriada para a caça, adaptou-se para ambientes pouco iluminados, e é facilitada pela movimentação dos objetos. Mas, comparando-se com a visão humana, a acuidade visual de cães é de três a quatro vezes pior, além de serem dicromatas, enquanto os humanos são tricromatas (Micklósi, 2007). O uso de modalidade sensorial olfativa em estudos subsequantes poderá ser mais bem sucedido. Mesmo tendo-se relatos de resultados positivos de discriminação visuais com cães na literatura (Range et al., 2008), o presente estudo exigia uma discriminação simultânea, diferentemente de Range et al. (2008), o que pode ter dificultado a discriminação. A opção pelo uso de estímulos visuais no presente estudo é justificada pela tentativa de replicar de maneira mais direta possível o estudo de Costa (2008).
Já a sensibilidade olfativa em cães é extensamente relatada na literatura. Cães são capazes de detectar a presença das mais diversas substâncias, como drogas (Furton & Myers, 2001), explosivos (Gazit, Goldblat, & Terkel, 2005) e até células cancerígenas (Balseiro & Correia, 2006). Williams e Johnston (2008) treinaram cães a responder diferencialmente na presença de uma classe de estímulos olfativos e a não responder na presença de outra classe.
Não se sabe ao certo que resultados podem ser advindos da presença do dono do sujeito Lucke, nas sessões. Acredita-se que o controle ambiental foi exercido de maneira a não permitir dicas dadas ao sujeito; além do uso dos óculos escuros pelo E2 (que podia ou não ser o dono do sujeito), os sujeitos respondiam de costas para os experimentadores, não podendo se valer de nenhuma dica, ao menos visual, dada por eles. No entanto, sugere-se que, em estudos posteriores, o dono do sujeito esteja ausente do ambiente experimental. Outra possibilidade é da presença do dono gerar respostas emocionais que atrapalhem o desempenho do sujeito, o que seria outro motivo para realizar sessões na ausência do dono.
Considerando o exposto antes, a presente pesquisa terá continuidade com a realização de um estudo que utilizará estímulos olfativos em um procedimento semelhante ao aqui descrito. Acredita-se que a utilização de estímulos de modalidade mais saliente para a espécie, somada a um maior controle de algumas variáveis (como a posição dos estímulos, o contato entre os sujeitos de gêneros diferentes e preparações mais cuidadosas para a transição das situações de treino para as de teste), são alterações metodológicas que devem favorecer o treino de formação de classes funcionais em cães domésticos (Canis familiaris). Espera-se assim avançar na avaliação do uso da espécie como modelo animal em estudos sobre aquisição de repertórios pré-simbólicos.
REFERÊNCIAS
Balseiro, S., & Correia, H. (2006). Is olfactory detection of human cancer by dogs based on major histocompatibility complex-dependent odor components? – A possible cure and a precocious diagnosis of cancer. Medical Hypothesies, 66, 270-272
Barros, R. S., Galvão, O. F., Brino, A. L. F., Goulart, P. R. K., & McIlvane, W. J. (2005). Variáveis de procedimento na pesquisa sobre classes de equivalência: contribuições para o estudo do comportamento simbólico. Revista Brasileira de Análise do Comportamento, 1,15-27. [ Links ]
Barros, R. S., Picanço, C. R. F., Costa, T. D., & Souza, C. B. A. (em preparação). Learning set de reversões de discriminações simples conjugadas: explorando procedimentos para formação de classes em macacos-prego. [ Links ]
Catania, A. C. (1999). Aprendizagem: Comportamento, linguagem e cognição (Trad. de Deisy das Graças de Souza et al.). Porto Alegre: ArtMed. [ Links ]
Cooper, J. J., Ashton, C., Bishop, S., West, R., Mills, D. S., & Young. R. J. (2003). Clever hounds: Social cognition in the domestic dog (Canis familiaris). Applied Animal Behaviour Science, 81, 229-244. [ Links ]
Costa, T. D. (2008). Repetidas mudanças de discriminações simples e formação de classes funcionais em animais. Tese de Doutorado, Programa de Pós-Graduação em Teoria e Pesquisa do Comportamento, Universidade Federal do Pará, Belém, PA. [ Links ]
Dahás, L. J. S. (2009). Formação de classes funcionais em cães domésticos (Canis familiaris). Dissertação de Mestrado, Programa de Pós-Graduação em Teoria e Pesquisa do Comportamento, Universidade Federal do Pará, Belém, PA. [ Links ]
Deacon, T. W. (1997). The symbolic species: The co-evolution of language and the brain. New York: Norton. [ Links ]
Domeniconi, C., Bortoloti, R., Antoniazzi, L. C. K., & Mendes, T. E. N., (2008). Treinos de discriminação simples e formação de classes funcionais de estímulos por cães. Interação em Psicologia, 12, 235- 243. [ Links ]
Frank, A. J., & Wasserman. E. A. (2005). Associative symmetry in the pigeon after successive matching-tosample training. Journal of the Experimental Analysis of Behavior, 84, 147-165. [ Links ]
Furton, K. G., & Myers, L. J., (2001) The scientific foundation and efficacy of the use of canines as chemical detectors for explosives. Talanta, 54, 487-500. [ Links ]
Gazit, I., Goldblat, A., & Terkel, J., (2005). The role of context specificity in learning: The effects of training context on explosives detection in dogs. Animal Cognition, 8, 143-150. [ Links ]
Hauser, M. D., Chomsky, N., & Fitch, W. T. (2002). The faculty of language: What is it, who has it, and how did it evolve? Science, 298, 1569-1579. [ Links ]
Hayes, S. C. (1989). Nonhumans have not yet shown stimulus equivalence. Journal of the Experimental Analysis of Behavior, 51, 385-392. [ Links ]
Horne, P. J., Lowe, C. F., & Harris, F. D. A. (2007). Naming and categorization in young children: V. Manual sign training. Journal of the Experimental Analysis of Behavior, 87, 367-381. [ Links ]
Kaminski, J., Call, J., & Fischer. J. (2004). Word learning in a domestic dog: Evidence for “fast mapping”. Science, 304, 1682-1683.
Kastak, C. R., Schusterman, R. J., & Kastak, D. (2001). Equivalence classifications by california see lions using class-specific reinforcers. Journal of the Experimental Analysis of Behavior, 76, 131-158. [ Links ]
Miklósi, A. (2007). Dog: behavior, evolution and cognition. New York: Oxford University Press. [ Links ]
Range, F., Aust, U., Steurer, M., & Huber, L. (2008). Visual categorization of natural stimuli by domestic dogs. Animal Cognition, 11, 339-347. [ Links ]
Rico, V. V. (2006). Persistência comportamental e topografi a de controle de estímulos coerente em treino de discriminação simples e escolha condicional por identidade ao modelo com quatro escolhas em macacos-prego (Cebus apella). Dissertação de Mestrado, Programa de Pós-Graduação em Teoria e Pesquisa do Comportamento, Universidade Federal do Pará, Belém, PA. [ Links ]
Rossi, A. P., & Ades, C. (2008). A dog at the keyboard: Using arbitrary signs to communicate. Animal Cognition, 11, 329-338. [ Links ]
Schusterman, R. J., & Kastak, D. (1993). A California sea lion (Zalophus californianus) is capable of forming equivalence relations. The Psychological Record, 43, 823-839. [ Links ]
Sidman, M. (2000). Equivalence relations and the reinforcement contingency. Journal of the Experimental Analysis of Behavior, 74, 127–146.
Sidman, M., & Tailby, W. (1982). Conditional discrimination vs. matching-to-sample: An expansion of the testing paradigm. Journal of the Experimental Analysis of Behavior, 37, 5-22. [ Links ]
Sidman, M., Wynne, C.K., Maguire, R.W., & Barnes, T. (1989). Functional classes and equivalence relations. Journal of the Experimental Analysis of Behavior, 52, 261-274. [ Links ]
Silva, F. T. N., & Souza, C. B. A. (2009). Discriminação simples com mudanças sucessivas na função dos estímulos: aprendizagem em bebês. Psicologia: Teoria e Pesquisa, 25, 569-580. [ Links ]
Soproni, K., Miklósi, A., Csányi, V., & Topál, J. (2001). Comprehension of human communicative signs in pet dogs (Canis familiaris). Journal of Comparative Psychology, 115, 112-126. [ Links ]
Souza, C. B. A., Borges, R. P., Goulart, P. R. K., Barros, R. S., & Galvão, O. F. (2009). Testes de identidade generalizada com objetos em macaco-prego (Cebus apella). Psicologia: Teoria e Pesquisa, 25, 169- 177. [ Links ]
Udell, M., & Wynne, C., (2008). A review of domestic’s dogs (Canis familiaris) human-like behaviors: or why behavior analysts should stop worrying and love their dogs. Journal of the Experimental Analysis of Behavior, 89(2), 247-261.
Vaughan, W. Jr. (1988). Formation of equivalence sets in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 14, 36-42. [ Links ]
Williams, M., & Johnston, J. M. (2002). Training and maintaining the performance of dogs (Canis familiaris) on an increasing number of odor discriminations in a controlled setting. Applied Animal Behaviour Science. 78, 55–65.
Wobber, V. E. (2005). The evolution of cooperative signal comprehension in the domestic dog (Canis familiaris). Tese de Doutorado, Department of Anthropology, Harvard University, Cambridge, Massachusetts. [ Links ]
Received: January 21, 2010
Accepted: June 11, 2010
1 Este trabalho corresponde a uma versão da dissertação de mestrado da primeira autora orientada pelo último autor, apresentada ao Programa de Pós-Graduação em Teoria e Pesquisa do Comportamento, da Universidade Federal do Pará, em 2009, com apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes), por meio de bolsa de estudo.
2 Correspondência para: Liane Dahás - lianedahas@gmail.com / Carlos Souza –carlos.souza@pesquisador.cnpq.br
3 O procedimento de pareamento ao modelo mais usado nos estudos sobre equivalência de estímulos é o de pareamento com atraso zero. Nesse procedimento, um estímulo modelo é apresentado, uma resposta a ele produz o seu desaparecimento e ocorre a apresentação imediata (daí o atraso zero) de dois ou mais estímulos (estímulos comparação). Um dos estímulos é arbitrariamente definido como correto (S+) e os demais como incorretos (S-). A escolha do S+ é reforçada e a do(s) S-(s) não. Ao longo do treino, dois ou mais estímulos modelos são positivamente relacionados com S+(s) específicos estabelecendo uma relação arbitrária entre eles.Texto integrante da tese de Livre-Docência do autor (Tomanari, 2008).
4 O procedimento padrão de discriminação simples Go/No-go consiste em apresentar de forma sucessiva estímulos com função positiva (S+) e com função negativa (S-). Respostas ao S+ são reforçadas e encerram uma tentativa. A não emissão de respostas durante a apresentação de um S- encerra a tentativa. Responder ao S- ou não responder ao S+ produz time-out.
5 Tradicionalmente, o termo utilizado para esse procedimento é “reversões sucessivas de discriminação simples” ou RSDS. No entanto, a terminologia “mudanças sucessivas de discriminação simples” ou MSDS passou a ser adotada recentemente por alguns pesquisadores (e.g. Silva & Souza, 2009; Souza, Borges, Goulart, Barros, & Galvão, 2009). O uso do novo termo se deve à maior adequação dele em procedimentos que utilizam mais de dois estímulos, ou em procedimentos que descartam a apresentação do estímulo previamente com função positiva (ver Rico, 2006, ou a seção Método do presente trabalho).
6 É necessário certificar-se que o responder de um sujeito em testes não foi treinado Durante o próprio teste. Portanto, muitos testes na área são realizados em extinção. Em outros casos, a conseqüência ao responder correto é mantida, mas somente a primeira tentativa com cada estímulo testado é levada em conta, já que o responder às outras pode ter sido ensinado com a consequenciação da primeira.

