










Services on Demand
article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.19 no.2 Guadalajara 2011
Ajuste del comportamiento bajo programas definidos temporalmente de diferente complejidad concurrentemente disponibles
Adjustment of behavior under concurrently available temporally defi ned schedules of different complexity
Mario Serrano, Brenda Castellanos; Abril Cortés-Zúñiga; Enoc Obed De la Sancha Villa; Gelacio Guzmán-Díaz1
Universidad Veracruzana Instituto de Psicología y Educación
RESUMEN
Tres ratas fueron expuestas a un programa concurrente cuyos componentes entregaron agua de acuerdo con diferentes tipos de arreglos de contingencia: no contingente, contingente y contingente condicional. Un cuarto componente nunca entregó agua. Se calcularon índices de ajuste correspondientes a –y en su caso incluidos en- los diferentes tipos de arreglos contingenciales: diferencialidad, efectividad y precisión. Dos ratas concentraron su actividad en el componente no contingente, pero para cada una se observaron índices de diferencialidad distintos. Para una tercera rata los índices de ajuste correspondientes a cada componente disminuyeron con la complejidad del arreglo contingencial. Para esa misma rata los índices de diferencialidad y efectividad entre componentes también disminuyeron. Estos resultados se discuten en el contexto de la psicología interconductual y se contrastan con los observados en estudios previos en el análisis experimental del comportamiento.
Palabras clave: índices de ajuste, programas concurrentes, no contingencia, contingencia, discriminación condicional, ratas.
ABSTRACT
Three rats were exposed to a concurrent schedule whose components delivered water according to different kinds of contingency arrangements: non-contingent, contingent and conditional-contingent. A fourth component never delivered water. Adjustment indexes for, and if included in, each contingency arrangement were calculated: differentiallity, effectiveness and accuracy. Two rats concentrated their activity in the noncontingent component but different differentiallity indexes were observed. For a third rat adjustment indexes in each component decreased with the complexity of contingency arrangements. For that rat differentiallity and effectiveness indexes between components also decreased. These results are discussed in the context of interbehavioral psychology and contrasted with those observed in previous studies in the experimental analysis of behavior.
Keywords: adjustment indexes, concurrent schedules, non-contingent, contingent, conditional discrimination, rats
De acuerdo con la propuesta taxonómica de Ribes y López (1985), en el caso de los organismos no lingüísticos el comportamiento psicológico puede tener lugar en tres niveles de organización funcional progresivamente más complejos e inclusivos: a) contextual; b) suplementario; y c) selector. La diferencia entre estos tres niveles de organización funcional del comportamiento radica en la mediación, el nivel de desligamiento funcional y el criterio de ajuste implicados en cada uno de ellos. El término mediación se refi ere al proceso por el cual un elemento ya sea de estímulo o de respuesta -el mediador- se torna crítico como propiedad estructurante del campo de contingencias. El término desligamiento funcional hace referencia a la relativa autonomía del responder del organismo respecto de las propiedades fi sicoquímicas de los eventos de estímulo, así como de los parámetros espaciotemporales que las defi nen en situación. Por su parte, el término criterio de ajuste hace referencia al requerimiento conductual que el organismo debe satisfacer en cada nivel de organización funcional del comportamiento. El criterio de ajuste implicado en la función contextual es la diferencialidad o ajustividad, es decir, la adecuación de la actividad del organismo respecto de los parámetros que defi nen en tiempo y espacio a los eventos de estímulo. En el caso de la función suplementaria el criterio de ajuste es la efectividad, es decir, la adecuación espaciotemporal de las respuestas del organismo para regular el contacto entre los eventos de estímulo. Finalmente, el criterio de ajuste implicado en la función selectora es la precisión, es decir, la adecuación pertinente de la efectividad del responder del organismo respecto de la variabilidad funcional momento a momento de los eventos de estímulo, en medida de su relación con eventos selectores (véase también Carpio, 1994; Ribes, 2004; Ribes, Moreno & Padilla, 1996).
En el contexto de la propuesta taxonómica de Ribes y López (1985), recientemente Serrano (2009) argumentó que los supuestos de complejidad e inclusividad progresivas que caracterizan a dicha propuesta implican que, dadas condiciones paramétricas equivalentes, los indicadores cuantitativos de los ritmos de desarrollo y estados terminales de la confi guración de las funciones contextual, suplementaria y selectora deberían guardar una relación inversa con su correspondiente complejidad. Asimismo, señaló que al menos al nivel de los estados terminales, los indicadores cuantitativos de una misma función deberían disminuir progresivamente conforme ésta es incluida en niveles más complejos de organización funcional del comportamiento. Mediante el cálculo de índices basados en los criterios de ajuste correspondientes a -y en su caso incluidos en- las funciones contextual, suplementaria y selectora, dicho autor confirmó ambas implicaciones empíricas al exponer a ratas a la entrega de agua no contingente (NK), contingente (K) y contingente condicional (KC). Específicamente, utilizando una cámara de desplazamiento y un programa concurrente con tres componentes operativos y uno no operativo, observó que los índices de diferencialidad bajo la entrega de agua NK superaron los índices de efectividad bajo la entrega de agua K, los que a su vez fueron mayores que los índices de precisión bajo condiciones de entrega de agua KC. Adicionalmente, encontró que los índices de diferencialidad y efectividad disminuyeron conforme aumentó entre fases la complejidad de los arreglos contingenciales para la entrega del agua.
Se planeó un estudio para comparar los ritmos de desarrollo y estados terminales de las funciones contextual, suplementaria y selectora bajo situaciones en las que los arreglos contingenciales que las auspician están simultáneamente disponibles. Específicamente, el presente estudio evaluó los efectos de la entrega concurrente de agua NK, K y KC, sobre los índices de diferencialidad, efectividad y precisión del comportamiento descritos por Serrano (2009). Un estudio por el estilo se consideró importante por dos razones principales. Por un lado, las implicaciones empíricas descritas por dicho autor fueron previstas para situaciones en las que los arreglos contingenciales correspondientes a cada nivel de organización funcional del comportamiento son enfrentados sucesivamente por el organismo, por lo que valdría la pena determinar si las relaciones entre ritmos de desarrollo y estados terminales antes señaladas se observan bajo condiciones de concurrencia intercontingencial. Por el otro lado, el estudio permitiría abordar, al menos exploratoriamente, la ejecución de los organismos bajo programas concurrentemente disponibles que, en contraste con la mayoría de los estudios en el área (Baum. 1974; Catania, 1966; Davison & McCarthy, 1988; de Villiers, 1977; Mazur, 1991), difieren cualitativa y no cuantitativamente.
MÉTODO
Sujetos
Se utilizaron tres ratas Wistar macho, experimentalmente ingenuas y de aproximadamente tres meses de edad al inicio del estudio. Las ratas fueron sometidas a un régimen de privación de agua de 23 horas diarias con acceso libre al alimento (Purina Rodent Lab Chow 5001 ®). Después de cada sesión experimental las ratas recibieron 30 min de acceso libre al agua en sus jaulas hogar, ubicadas en una colonia con temperatura controlada y un ciclo luz-oscuridad 12:12.
Aparatos
Se utilizó una cámara de desplazamiento (Coulbourn Instruments ®, Habitest Labline L91-165) de 92 cm de ancho x 92 cm de largo y 33 cm de altura. A 2 cm del piso de rejilla y a 14.14 cm de la pared lateral derecha correspondiente a cada pared de la cámara de desplazamiento, se colocaron cuatro dispensadores de agua (Coulbourn H14-05R) provistos con un detector de entradas (MED ENV-254). Únicamente tres de los cuatro dispensadores fueron operativos, sin embargo, también se realizaron registros de entrada en el dispensador restante. Los dispensadores operativos dieron acceso a 0.01 cc de agua durante 3 s y en cada activación fueron iluminados por focos blancos de 5 w. En la parte superior de cada dispensador se colocaron tres módulos de estímulo triple (MED ENV-222M), compuestos por leds de color rojo, amarillo y verde. Adicionalmente, a los lados derecho e izquierdo de cada dispensador se colocó una palanca retráctil (MED ENV-110M) que requirió una fuerza de 0.2 N para el cierre del microinterruptor. Únicamente las palancas de tres de las cuatro paredes de la cámara fueron operativas, sin embargo, también se registraron las respuestas en las palancas de la pared restante. La programación y registro de los eventos se realizó mediante una computadora provista con una interfaz (MED SG-6010) y software (MED-PC ® IV) especializados.
PROCEDIMIENTO
Las ratas fueron expuestas a la entrega concurrente de agua NK, K y KC de acuerdo con tres programas defi nidos temporalmente paramétricamente equivalentes: T = 60 s, tD= 30 s y tΔ = 30 s. Los programas fueron independientes entre sí y cada uno se correlacionó con un panel operativo. El programa que entregó el agua de manera NK se correlacionó con el Panel 1, el programa que entregó el agua de manera K se correlacionó con el Panel 2 y el programa que entregó el agua de manera KC se correlacionó con el Panel 3. En el Panel 4 nunca se entregó agua. El estudio estuvo vigente durante 30 sesiones de 30 minutos de duración.
En el caso de la entrega de agua NK, el subciclo tDse señaló mediante los tres leds rojos y el subciclo tΔ se señaló mediante el led amarillo central de la matriz de leds correspondiente. El dispensador de agua se activó al final del subciclo tDindependientemente de la actividad de las ratas. En el caso de la entrega de agua K, la primera respuesta al interior del subciclo tDsobre cualquiera de las dos palancas del Panel 2 activó el dispensador de agua y, adicionalmente, canceló los tres leds rojos con los que se señaló dicho subciclo en ese panel. Durante el subciclo tΔ, que se señaló mediante el led amarillo central de la matriz correspondiente, las respuestas no tuvieron consecuencias programadas. Finalmente, en el caso de la entrega de agua KC, la primera respuesta sobre una u otra palanca del Panel 3 durante el subciclo tDtambién produjo la entrega del agua y la terminación de la señal correlacionada con dicho subciclo. La efectividad de la respuesta, sin embargo, estuvo determinada por las señales correlacionadas con el subciclo tD. Los tres leds rojos señalaron la operatividad de la palanca derecha, mientras los tres leds verdes señalaron la operatividad de la palanca izquierda. La señalización del subciclo tDpor una u otra triada de leds se determinó en forma aleatoria. Al igual que en los casos anteriores, las respuestas en el subciclo tΔ no tuvieron consecuencias programadas y el subciclo se señaló mediante el led amarillo central de la matriz correspondiente.
Análisis de datos
La ejecución de las ratas en el programa concurrente se determinó a partir de índices de ajuste correspondientes a los requerimientos conductuales o criterios de ajuste implicados en cada uno de los arreglos contingenciales: diferencialidad, efectividad y precisión. En línea con el supuesto de inclusividad progresiva de la propuesta taxonómica de Ribes y López (1985), el índice de diferencialidad se calculó bajo las condiciones de entrega de agua NK, pero también en los casos de entrega de agua K y KC. Asimismo, el índice de efectividad se calculó bajo las condiciones de entrega de agua K, pero también en el caso de la entrega de agua KC. El índice de diferencialidad se calculó como la diferencia entre la proporción de tiempo que las ratas interrumpieron los detectores de entrada a los bebederos en presencia del agua respecto del tiempo total de disponibilidad de agua, y la proporción de tiempo que interrumpieron los detectores de entrada a los bebederos en ausencia del agua respecto del tiempo total de la sesión.
El índice de efectividad se calculó como la proporción de entregas de agua producidas respecto de las programadas, multiplicada por el cociente de las entregas de agua producidas y el total de palancazos implicados en su producción. El índice de precisión se calculó como el número de aciertos, relativo a la suma de éstos y los errores por comisión y omisión.
RESULTADOS
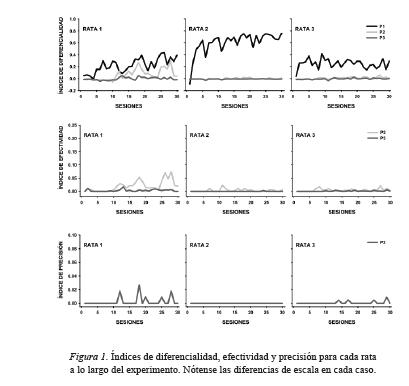
La Figura 1 muestra los índices de diferencialidad (primera fi la), efectividad (segunda fi la) y precisión (tercera fi la) observados para cada rata a lo largo del estudio. Los índices de diferencialidad en el Panel 1 fueron bajos para las ratas 1 y 3, mientras para la Rata 2 se observaron índices de diferencialidad más altos desde las primeras sesiones experimentales. Para la Rata 1 el índice de diferencialidad en el Panel 2 disminuyó respecto del observado en el Panel 1, y disminuyó aún más en el Panel 3. Para las otras dos ratas, los índices de diferencialidad en los paneles 2 y 3 fueron cercanos a cero a lo largo de las sesiones. Aunque ambos incipientes, el índice de efectividad fue más elevado en el Panel 2 que en el Panel 3 para todas las ratas. El índice de efectividad fue más cercano a cero para las ratas 2 y 3 que para la Rata 1. El índice de efectividad en el Panel 3 fue prácticamente igual a cero para la Rata 2, mientras para las otras dos ratas incrementó ligeramente en algunas sesiones. El mismo patrón se observó para el índice de precisión en el Panel 3, si bien los incrementos esporádicos en la precisión fueron mayores para la Rata 1 que para la Rata 3.
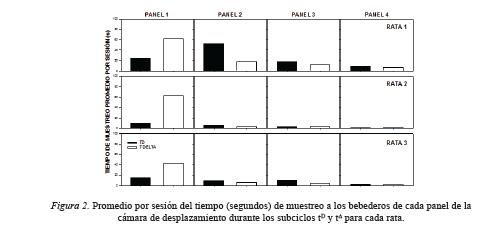
La Figura 2 muestra el tiempo promedio (segundos) de muestreo a los bebederos durante los subciclos tDy tΔ en los cuatro paneles de la cámara de desplazamiento para cada rata. Para las tres ratas el tiempo de muestreo en el Panel 1 fue mayor durante el subciclo tΔ que durante el subciclo tD, mientras en los paneles 2 y 3 se observó el efecto inverso para las ratas 1 y 3. Para la Rata 2 en los paneles 2 a 4 y las ratas 1 y 2 en el Panel 4, no se observaron diferencias claras entre ambos subciclos en el tiempo de muestreo a los bebederos.
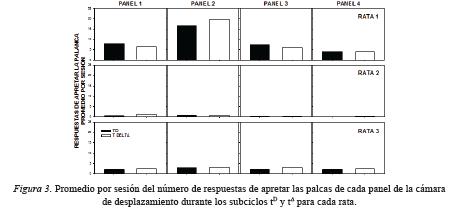
La Figura 3 muestra el promedio por sesión del número de respuestas de apretar la palanca durante los subciclos tDy tΔ en los cuatro paneles de la cámara de desplazamiento para cada rata. Para la Rata 1 el número de respuestas de apretar la palanca fue mayor en el subciclo tDque en el subciclo tΔ en los paneles 1, 3 y 4, mientras en el Panel 2 se observó el efecto inverso y el mayor número de respuestas totales. Para las otras dos ratas no se observaron diferencias claras entre los subciclos y paneles.
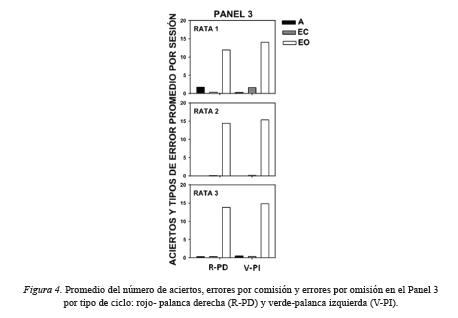
La Figura 4 muestra el promedio por sesión de los aciertos, errores por comisión y errores por omisión individuales en el Panel 3 por tipo de ciclo: rojo-palanca derecha y verde-palanca izquierda. Los errores por omisión fueron superiores que los aciertos y errores por comisión para las tres ratas. Para la Rata 1 se observaron más aciertos que errores por comisión en la palanca derecha, mientras en la palanca izquierda se observó el efecto inverso. Para la Rata 3 se registró un acierto y un error por comisión en la palanca derecha y dos aciertos y un error por comisión en la palanca izquierda. Para la Rata 2 sólo se observaron errores por comisión.
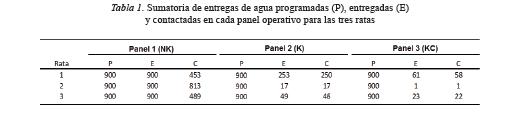
La Tabla 1 muestra la sumatoria de entregas de agua programadas, entregadas y contactadas en cada panal operativo para las tres ratas. El número de entregas de agua contactadas disminuyó conforme incrementó la complejidad de los arreglos contingenciales para la entrega del agua. En el caso del Panel 1, las ratas 1 y 3 contactaron alrededor del 50% de las entregas de agua, mientras la Rata 2 alcanzó el 90%. En el Panel 2, las ratas 2 y 3 produjeron menos del 10% de las entregas de agua programadas, mientras la Rata 1 produjo cerca del 30%. En ese mismo panel, la proporción de entregas de agua producidas respecto de las contactadas fue elevada para las tres ratas. El mismo efecto se observó en el Panel 3. Nuevamente para las ratas 2 y 3 se observó el menor porcentaje de entregas de agua, mientras para la Rata 1 las entregas de agua producidas fueron menores al 10%.
DISCUSIÓN
En el contexto de los supuestos de complejidad e inclusividad progresivas que caracterizan a la propuesta taxonómica de Ribes y López (1985), el presente estudio evaluó los efectos de la entrega concurrente de agua NK, K y KC sobre los índices de diferencialidad, efectividad y precisión del comportamiento (Serrano, 2009). Se observó que dos de las tres ratas escogieron más frecuentemente el Panel 1, es decir, aquel en el que el agua se entregó de manera NK. La rata restante distribuyó su actividad entre los tres paneles operativos de la cámara de desplazamiento, aunque dicha actividad disminuyó con la complejidad de los arreglos contingenciales para la entrega del agua. Para esta rata, se observó que el índice de diferencialidad en el Panel 1 fue superior al índice de efectividad en el Panel 2, es decir en el que el agua se entregó de manera K, así como que dicho índice de efectividad fue superior al índice de precisión en el Panel 3, en el que el agua se entregó de manera KC. Adicionalmente, para esa misma rata se observó que los índices de diferencialidad y efectividad calculados en los paneles 2 y 3 disminuyeron con la complejidad de los arreglos contingenciales para la entrega del agua.
Los resultados del presente estudio indican que las relaciones entre ritmos de desarrollo y estados terminales descritas por Serrano (2009) en torno de la confi guración de las funciones contextual, suplementaria y selectora reconocidas en la propuesta taxonómica de Ribes y López (1985), son posibles bajo condiciones en las que los arreglos contingenciales que auspician dichos niveles de organización funcional del comportamiento están simultáneamente disponibles. Sin embargo, al mismo tiempo indican que bajo tales circunstancias los organismos "prefi eren" los arreglos contingenciales que implican los criterios de ajuste de menor complejidad. Resultados análogos a los del presente estudio han sido reportados previamente en la literatura experimental del análisis de la conducta. Bajo programas de reforzamiento concurrente, por ejemplo, se ha observado que los organismos concentran su actividad en el componente que implica el menor número de respuestas por reforzador, principalmente cuando los requisitos de respuesta difieren notablemente (e.g., Herrnstein, 1958; Herrnstein & Loveland, 1975). Por el otro lado, el hecho de que una rata haya distribuido su actividad entre los tres paneles operativos y que la segunda haya disminuido conforme se incrementó la complejidad de los arreglos contingenciales, concuerda con diversos estudios en los que se ha reportado que los organismos persisten en la producción de "reforzadores" bajo condiciones en las que éstos se entregan libremente, así como que dicha persistencia disminuye con el esfuerzo implicado en la producción (véase Inglis, Forkman & Lazarus, 1997). En el caso del presente estudio, sin embargo, debe destacarse que el término esfuerzo estaría referido al nivel de desligamiento funcional implicado en la organización funcional del comportamiento, más que a los incrementos en los parámetros intensivos de un mismo tipo de procedimiento.
En lo que respecta a las ejecuciones locales en los paneles y durante los subciclos tDy tΔ, las observaciones descritas en el presente reporte contrastan con las realizadas en estudios previos. Al respecto, destacan los tiempos de muestreo a los bebederos de los paneles 1 y 2, en los que el agua se entregó de manera NK y K, respectivamente. Estudios anteriores sobre condicionamiento pavloviano en los que se utilizó la entrada al comedero como respuesta condicional, reportaron que el muestreo al comedero fue mayor durante el estimulo condicional (EC) que durante periodos equivalentes inmediatamente anteriores a la presentacion de dicho estimulo (e.g., Bueno & Alvarez, 2001). Adicionalmente, estudios en los que se analizo momento a momento la actividad de los organismos bajo programas de reforzamiento NK y K basados en el paso del tiempo, senalaron que la actividad vinculada morfologica y geografi camente con la entrega del reforzador ocurre al final (i.e., conductas terminales) y no al principio (i.e., conductas interinas) del intervalo ente reforzadores (Staddon & Simmelhag, 1971). En el presente estudio, sin embargo, en el Panel 1 se registro un mayor tiempo de interrupcion de los detectores de entrada al bebedero durante el subciclo tΔ que durante el subciclo tD, mientras que solo en el Panel 2 se observo el efecto inverso.
Es posible suponer que en el caso del Panel 1, la concentracion del tiempo de muestreo durante el subciclo tΔ haya estado regulada por los valores parametricos utilizados. En el experimento de Bueno y Alvarez (2001), por ejemplo, el numero de muestreos fue mayor durante el periodo previo al EC que durante la presentacion de dicho estimulo mientras menor fue la razon entre el intervalo entre ensayos y el intervalo entre estimulos. Una interpretacion por el estilo, sin embargo, no explicaria el hecho de que en el Panel 2, bajo los mismos valores parametricos pero condiciones de entrega de agua K, el tiempo de muestreo haya sido menor que en el Panel 1 y la distribucion al interior del ciclo T se invirtiera. En el contexto de estos resultados, pareciera que la posibilidad de ubicar las llamadas conductas interinas y terminales descritas por Staddon y Simmelhag (1971) al inicio y final del intervalo entre reforzadores, respectivamente, estaria condicionada por los valores parametricos de los procedimientos implementados y el tipo de arreglo contingencial, mas que por estados motivacionales reales o inferidos.
Tambien sobre el tiempo de muestreo al bebedero del Panel 1, es de destacar que al menos para la rata que distribuyo su actividad entre los tres paneles operativos, la Rata 1, la distribucion de dicha variable al interior del ciclo T fue similar a la distribucion de respuestas de apretar la palanca en el Panel 2, la que a su vez replico los resultados de varios experimentos anteriores sobre control del estimulo en los que se utilizaron programas defi nidos temporalmente similares a los del presente estudio (e.g., Ribes, Mayoral, Torres & Ibanez, 2000; Ribes, Torres & Mayoral, 2002). En tales experimentos se reporto que las ratas emiten un mayor numero de respuestas durante el subciclo tΔ que durante el subciclo tD, cuyas senales agregadas se correlacionan negativa y positivamente con la entrega del agua, respectivamente. A diferencia de los argumentos vertidos en tales experimentos en los que el agua se entrego de manera K, la ausencia de control del estimulo observada en el presente estudio bajo condiciones de entrega de agua NK no puede explicarse en terminos de reforzadores perdidos. Aunque para la Rata 1 las entregas de agua contactadas en el Panel 1 fueron solo el 50% de las entregas de agua programadas, en el caso de la rata para la que se observo el indice de diferencialidad mas elevado en ese mismo panel, la Rata 2, el porcentaje de entregas de agua contactadas fue del 90% y, no obstante, el tiempo de muestreo al bebedero del Panel 1 tambien fue mayor durante el subciclo tΔ que durante el subciclo tD. Estos resultados son de llamar la atencion si se considera que para la Rata 3, que al igual que la Rata 2 concentro su actividad en el Panel 1, el indice de diferencialidad y el porcentaje de entregas de agua contactadas en dicho panel fueron similares a los observados para la Rata 1, y la distribucion del tiempo de muestreo igualmente fue mayor durante el subciclo tΔ que durante el subciclo tD.
Dicho en otros terminos, la ausencia de control por parte de las senales correlacionadas con los subciclos tDy tΔ sobre el tiempo de muestreo al bebedero en el caso de la entrega de agua NK, no es atribuible a la proporcion de entregas de agua no contactadas, la cantidad absoluta de agua consumida ni tampoco a la disponibilidad de otros dispositivos de respuesta, se entre o no en contacto con ellos. Dado que disminuir las dimensiones del espacio experimental incrementa la actividad en torno de los dispositivos de respuesta (e.g., Skuban & Richarson, 1975), no es osado suponer que incrementarlo produce el efecto inverso. Sin embargo, atribuir los resultados del tiempo de muestreo al bebedero del Panel 1 directamente a las dimensiones del espacio experimental, solo explicaria los bajos valores de la variable en cuestion observados durante el sub ciclo tDy no los altos valores registrados durante el subciclo tΔ. Una posibilidad explicativa, complementaria a las dimensiones del espacio experimental y que requeriría de una exploración empírica más detallada, radica en el uso del agua como estímulo motivacionalmente relevante. Estudios anteriores han reportado que la actividad en torno de los dispositivos de respuesta está altamente determinada por las características de las respuestas de consumo de los estímulos, tanto bajo condiciones de entrega NK (e.g., Jenkins & More, 1973) como K (e.g., Wolin, 1948) y KC (e.g., Brodigan & Peterson, 1976). En esa medida, es posible suponer que las respuestas inmediatamente posteriores al consumo de los estímulos motivacionalmente relevantes también difieren dependiendo del estímulo del que se trate. Estudios venideros deberán atender a dicha posibilidad. En cualquier caso, lo cierto es que una interpretación por estilo no sólo explicaría los altos tiempos de muestreo al bebedero del Panel 1 durante el subciclo tΔ para todas las ratas, sino igualmente la inversión en la distribución de dicha variable al interior del ciclo T en el bebedero del Panel 2 para la Rata 1. Al respecto, cabe destacar que el control por parte de las señales correlacionadas con los subciclos tDy tΔ sobre el tiempo de muestreo se tornaría un artefacto del estímulo motivacionalmente relevante utilizado. En esa medida, la ausencia de control del estímulo en programas defi nidos temporalmente antes señalada podría extenderse a respuestas diferentes a la de apretar la panca.
Sobre la ejecución en el panel en el que el agua se entregó de manera KC, por un lado, los resultados del presente estudio concuerdan con los descritos por Serrano (2009) en lo referente a los bajos índices de ajuste observados en el caso de arreglos contingenciales que implican discriminaciones condicionales para la entrega del agua. Por el otro, indican que tales índices de precisión obedecen principalmente a errores por omisión (ausencia de respuestas) y no a errores por comisión (respuestas incorrectas). Además de que en el estudio anteriormente citado no se analizó la ejecución en términos del número de aciertos y del tipo de errores cometidos, ambos puntos son importantes por al menos dos razones. En primer lugar, porque en conjunto, los resultados de ambos estudios impiden atribuir los bajos índices de precisión a la disponibilidad de arreglos contingenciales que implican discriminaciones condicionales exclusivamente, o bien arreglos contingenciales de menor complejidad. En esa medida, en segundo lugar, los resultados de ambos estudios sugieren que futuras exploraciones deberán enfocarse al papel jugado por la dimensión de las señales correlacionadas con los subciclos tDy tΔ, los valores de ambos períodos y, principalmente, el tamaño del espacio experimental, ya que al parecer el uso de espacios de un tamaño mayor a los usualmente utilizados propicia el despliegue de respuestas o segmentos de actividad incompatibles con los requeridos por los arreglos contingenciales.
Dado que los índices de ajuste más bajos fueron los índices de precisión, y en la medida que éstos se debieron principalmente a errores por omisión y no a errores por comisión incluso en el caso de la Rata 1, los resultados del presente estudio también sugieren que los efectos del tamaño del espacio experimental en términos de las respuestas incompatibles antes mencionadas son relativos a la complejidad de los arreglos contingenciales. De ser este el caso, por un lado, debe destacarse que los resultados aquí descritos apoyan la idea de que los arreglos de entrega agua implementados en cada panel operativo corresponden a las contingencias de ocurrencia (Ribes, 1995) que auspician el desarrollo de las funciones que se pretendió evaluar. Por el otro lado, y dado que la disminución progresiva de los índices de ajuste constituye el resultado conceptualmente esperado, al menos los resultados observados para la Rata 1 del presente estudio sugieren que el moldeamiento de la respuesta de apretar la palanca no es requisito para analizar el comportamiento bajo programas concurrentemente disponibles. Desde el punto de vista interconductual, ésta posibilidad es de suma importancia en la medida que dicho procedimiento necesariamente "contamina" el desarrollo de nuevas funciones psicológicas.
Finalmente, en lo tocante al Panel 4, es decir, aquel en el que nunca se entregó agua, los resultados del presente estudio no sólo replican efectos bien conocidos sobre los procedimientos de extinción, sino igualmente los relativos a la intermitencia en la entrega de los estímulos motivacionalmente relevantes. Específicamente, aunque en el Panel 4 se registraron los menores tiempos de muestreo al bebedero y respuestas de apretar la palanca para todas las ratas, ambas variables fueron más altas para la Rata 1, es decir, la rata expuesta a la mayor intermitencia de entrega de agua en medida de la distribución de su actividad entre los tres paneles operativos y las características de los arreglos contingenciales correspondientes. Estos resultados confi rman la viabilidad de utilizar panales no operativos para detectar posibles respuestas incompatibles y/o irrelevantes para la satisfacción de los criterios implicados en los arreglos contingenciales, así como la inclusión de los registros correspondientes en el cálculo de los índices de ajuste. Nuevos estudios deberán evaluar el incremento o disminución en el número de paneles no operativos.
Referencias
Baum, W. M. (1974). On two types of deviation from the matching law: Bias and undermatching. Journal of the Experimental Analysis of Behavior, 22, 231-242. [ Links ]
Brodigan, D. L., & Peterson, G. B. (1976). Two-choice conditional discrimination performance of pigeons as a function of reward expectancy, prechoice delay and domesticity. Animal Learning and Behavior, 4, 121-124. [ Links ]
Bueno, M., & Álvarez, R. (2001). El efecto de las duraciones del intervalo entre ensayos y entre estímulos en el condicionamiento pavloviano apetitivo en ratas. Psicológica, 22, 205-215. [ Links ]
Carpio, C. (1994). Comportamiento animal y teoría de la conducta. En L. J. Hayes, E. Ribes, & F. López Valadez (Eds.), Psicología interconductual: Contribuciones en honor a J. R. Kantor (pp. 45-68). México: Universidad de Guadalajara. [ Links ]
Catania, A. C. (1966). Concurrent operants. En W. K. Honig (Ed.), Operant behavior: Areas of research and application (pp. 213-270). New York: Appleton Century Crofts. [ Links ]
Davison, M., & McCarthy, D. (1988). The matching law: A research review. Hillsdale, NJ: Lawrence Erlbaum Associates. [ Links ]
de Villiers, P. A. (1977). Choice in concurrent schedules and a quantitative formulation of the law of effect. In W. K. Honig & J. E. R. Staddon (Eds.), Handbook of operant behavior (pp. 233–287). Englewood Cliffs, NJ: Prentice Hall.
Herrnstein, R. J. (1958). Some factors infl uencing behavior in a two-response situation. Transactions of the New York Academy of Science, 21, 35-45. [ Links ]
Herrnstein, R. J., & Loveland, D. H. (1975). Maximizing and matching on concurrent ratio schedules. Journal of the Experimental Analysis of Behavior, 24, 107-116. [ Links ]
Inglis, I. R., Forkman, B., & Lazarus, J. (1997). Free food or earned food? A review and fuzzy model of contrafreeloading. Animal Behaviour, 53, 1171-1191. [ Links ]
Jenkins, H. M., & Moore, B. R. (1973). The form of auto-shaped response with food or water reinforcers. Journal of the Experimental Analysis of Behavior, 20, 163-181. [ Links ]
Mazur, J. E. (1991). Choice. En I. H. Iversen & K. A. Lattal (Eds.) Techniques in the behavioral and neural sciences: Experimental analysis of behavior, Part 1 (pp. 219-250). Amsterdam: Elsevier. [ Links ]
Ribes, E. (2004). Acerca de las funciones psicológicas: Un post-scriptum. Acta Comportamentalia, 12, 117- 127. [ Links ]
Ribes, E. (1995). Causalidad y contingencia. Revista Mexicana de Análisis de la Conducta, 21(Mono), 133-150. [ Links ]
Ribes, E., & López, F. (1985). Teoría de la conducta: Un análisis de campo y paramétrico. México: Trillas. [ Links ]
Ribes, E., Mayoral, A., Torres, C., & Ibáñez, F. J. (2000). Effects of auditory stimuli correlated with different probabilities of water delivery in a limited-hold temporal schedule. Behavioural Processes, 52, 49-59. [ Links ]
Ribes, E., Moreno, R., & Padilla, A. (1996). Un análisis funcional de la práctica científica: extensiones de un modelo psicológico. Acta Comportamentalia, 4, 203-232. [ Links ]
Ribes, E., Torres, C., & Mayoral, A. (2002). Extended exposure to a discriminated, limited-hold temporal schedule does not produce stimulus control. Behavioural Processes, 59, 131-146. [ Links ]
Serrano, M. (2009). Complejidad e inclusividad progresivas: Algunas implicaciones y evidencias empíricas en el caso de las funciones contextual, suplementaria y selectora. Revista Mexicana de Análisis de la Conducta, 35 (monográfico), 161-178. [ Links ]
Skuban, W. E., & Richardson, W. K. (1975). The effect of the size of the test environment on behavior under two temporally defi ned schedules. Journal of the Experimental Analysis of Behavior, 23, 271-275. [ Links ]
Staddon, J. E. R., & Simmelhag, V. L. (1971). The "superstitious" experiment: A reexamination of its implications for the principles of adaptive behavior. Psychological Review, 78, 3-43. [ Links ]
Wolin, B. R. (1948). Difference in manner of pecking a key between pigeons reinforced with food and water. En A. C. Catania (Ed.), Contemporary research in operant behavior (p. 286). New York: Scott, Foresman. [ Links ]
Received: November 9, 2010
accepted March 24, 2011
1) La presente investigación fue posible gracias al proyecto PROMEP/103.5/10/5006 otorgado al primer autor. Brenda Castellanos y Gelacio Guzmán-Díaz recibieron, respectivamente, las becas 236107 y 236118 del Consejo Nacional de Ciencia y Tecnología para cursar la Maestría en Investigación en Psicología Aplicada a la Educación. Enviar correspondencia a Mario Serrano, Universidad Veracruzana, Instituto de Psicología y Educación, Agustín Melgar y Juan Escutia s/n. Col. Revolución. Xalapa, Veracruz. México, C. P. 91100 o al correo electrónico mserrano@uv.mx
