










Services on Demand
article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.20 no.2 Guadalajara 2012
ARTÍCULOS
Relación entre distintas densidades locales del agua y densidades locales de respuesta sobre la frecuencia total de respuesta en un programa temporal señalado
Relationship between different local densities of water and local densities of responses on the overall frequency of responding in a signaled temporal Schedule
Emilio Ribes Iñesta*; Sergio Arenas**; Alfredo Mayoral**
*Universidad Veracruzana (México)
**Universidad de Guadalajara (México)
RESUMEN
Se realizó un experimento con ocho ratas albinas Wistar machos. Se comparó el efecto de entregar el agua en los 20 ciclos iniciales o finales con su distribución en cada tercer ciclo de un programa definido temporalmente sobre el número total de respuestas por sesión. El agua se entregó contingente a la primera respuesta en el periodo tD de cada ciclo de 60 s y, además, en caso de no haber respuesta, de manera no contingente al final del periodo cada tercer ciclo en tres de las fases, o en los primeros veinte o últimos veinte ciclos en las dos fases restantes. Los periodos tD y tΔ se correlacionaron con estímulos exteroceptivos diferentes. Las ratas se distribuyeron en dos grupos, expuestos a las mismas fases experimentales, con la diferencia de que en un grupo la palanca permanecía todo el tiempo durante la sesión, mientras que en el otro se empleaba una palanca retráctil que se retraía después de la entrega del agua durante los periodos seleccionados para entregar de manera alternativa el agua de manera no contingente. Los resultados mostraron frecuencias semejantes de respuestas en los dos periodos del ciclo T, mayor número de respuestas en el grupo con palanca normal, y un elevado porcentaje de entregas de agua contingentes obtenidas, sugiriendo que la frecuencia total de respuesta fue una función de la frecuencia total de entregas de agua, y no de su localización durante la sesión.
Palabras clave: densidad local de agua, densidad local de respuesta, programas temporales, palanca retráctil, porcentaje de agua obtenido.
ABSTRACT
An experiment was conducted with eight male albino Wistar rats. It was compared the effect of delivering water in the twenty initial or final cycles or when distributed every third cycle of a temporally defined schedule upon the total number of responses per session. Water delivery was contingent to the first response during the tD period of each 60 s cycle and, in addition to that, if the rats did not respond, water was dispensed at the end on the first or final 20 cycles in two of the experimental, or in every third cycle in other three phases. tD and tΔ periods were signaled by different exteroceptive stimuli. Rats were distributed in two groups, exposed to the same experimental conditions, except for the fact that rats in one group had a retractable lever that was withdrawn during tD after the first response, in those selected cycles in which non contingent water delivery was also scheduled. Results of the experiment showed similar response frequencies in both periods of the T cycle, a larger number of responses in the group with normal, non-retractable lever, and a high percentage of obtained contingent-water deliveries, suggesting that overall frequency of responses was a function of overall frequency of water deliveries, irrespectively of its allocation during the session.
Keywords: local density of water, local density of responses, temporal schedules, retractable lever, percentage of obtained water deliveries.
Los programas temporales de estímulo (Schoenfeld & Cole, 1972) consisten en ciclos repetitivos (T) en los que se puede prescribir la entrega de un estímulo "neutro" y/o "reforzante" con base en dos parámetros independientes. Uno de estos parámetros es T, que describe el tiempo proporcional de entrega contingente de un estímulo (agua o comida) dada la ocurrencia de una respuesta, o la entrega de dicho estímulo independientemente de la respuesta, respecto del valor del ciclo T. El subciclo en que la disponibilidad del estímulo es igual a cero se denota como tΔ, mientras que el tiempo con disponibilidad del estímulo se denota como tiempo discriminativo o tD. El otro parámetro es P, que describe la probabilidad de que en un ciclo determinado esté disponible un estímulo, sea o no contingente. Mientras que P describe la frecuencia relativa de estímulos dado el total de ciclos, T describe el tiempo relativo de disponibilidad del estímulo dentro del ciclo.
En los programas temporales de estímulo, a diferencia de los programas de reforzamiento tipo Ferster y Skinner (Ferster & Skinner, 1957), la disponibilidad del estímulo denominado "reforzante" es limitada en tiempo, cuando su entrega se prescribe contingente a la ocurrencia de una respuesta. Su disponibilidad, delimitada por el periodo tD, permite que la primera respuesta que ocurre en dicho periodo produzca la entrega del estímulo. Sin embargo, de no ocurrir respuesta alguna en dicho periodo, el estímulo se "pierde", es decir, deja de estar disponible y no vuelve a presentarse, de manera que en una sesión experimental el número de estímulos entregados puede ser menor al número de estímulos programados. Por consiguiente, en los programas temporales, aunque la frecuencia de respuesta es un parámetro importante, el factor fundamental se relaciona con la oportunidad y ubicación temporal de la respuesta, la primera respuesta que ocurre en el tiempo discriminativo.
Ribes y colaboradores (e.g., Ribes, Mayoral, Torres & Ibañez, 2000; Ribes, Torres & Mayoral, 2002; Ribes, Zepeda, Arenas & Mayoral, 2007) al examinar las interacciones de P,T, y la presencia/ausencia de estímulos correlacionados con la entrega o ausencia del agua en programas temporales, han encontrado bajas frecuencias de respuesta, variaciones importantes de frecuencia de respuesta entre sesiones, y porcentajes significativos de pérdida de entregas de agua programada. Un hallazgo consistente es que la mayor parte de las respuestas ocurren durante el periodo tΔ (con probabilidad cero o menor que en el periodo tD), y que los incrementos o decrementos de respuesta tienen lugar durante tΔ, pues la frecuencia de respuesta en tD es usualmente baja y constante, a pesar de que pueden seguir ocurriendo respuestas en dicho periodo después de la entrega del agua mientras no se inicia el periodo tΔ.
En dos de los estudios realizados (Ribes et al. 2000; Ribes et al. 2002) empleando programas temporales con estímulos correlacionados diferenciales en ambos subciclos, se observó que las ratas con mayor frecuencia de respuesta y/o menor porcentaje de entregas de agua perdidas fueron aquellas que obtuvieron mayor proporción de entregas de agua en los primeros 15 ciclos de cada sesión. Esta observación sugiere que la frecuencia total de respuesta en una sesión puede depender del porcentaje de entregas de agua obtenidas en los primeros ciclos, de modo que si se pierden muchas entregas de agua en el principio de la sesión, la frecuencia total de respuesta puede disminuir.
La frecuencia total de respuesta en cada ciclo no es necesariamente el determinante único o principal en la obtención de las entregas de agua programadas. Como se mencionó, la mayor parte de las respuestas ocurren regularmente en el ciclo tΔ, en que no hay entregas programadas y, aunque pudiera plantearse que dichas respuestas pudieran funcionar de manera equivalente a una ejecución de intervalo fijo concluyendo con la primera respuesta en el subciclo tD y la entrega consiguiente de agua, el análisis de las latencias de las primeras respuestas en los estudios arriba citados descartó esta posibilidad. Se observaron latencias mayores a 4 s en la primera respuesta en el periodo tD, lo que contradice una ejecución terminal positivamente acelerada iniciada en el subciclo tΔ previo inmediato.
En dos experimentos adicionales, Ribes et al. (2007) exploraron el efecto de las entregas de agua en los primeros o los últimos veinte ciclos de la sesión sobre la regularidad de la frecuencia de respuesta entre sesiones, la pérdida de entregas de agua y la frecuencia relativa de respuesta en cada bloque de ciclos de entrega. En el primer experimento se evaluó el efecto de entregar efectivamente el agua en los 20 ciclos iniciales o terminales de un programa temporal sobre la frecuencia total de respuestas en la sesión, independientemente del número total de entregas de agua recibidas. El agua se entregó de manera contingente a la primera respuesta en cada subciclo tD y, además, en caso de que las ratas no respondieran en los bloques de ciclos iniciales o terminales seleccionados, el agua se entregó de manera no contingente a la respuesta al concluir el periodo tD particular. Los periodos tD y tΔ se correlacionaron con estímulos diferenciales. En el segundo experimento se mantuvo el mismo procedimiento, exceptuando que cuando las ratas respondían en el periodo tD de los bloques de ciclos seleccionados, se empleaba una palanca retráctil que se retiró inmediatamente después de la respuesta productora del agua, impidiendo más respuestas durante el subciclo. La palanca se reintrodujo al terminar el subciclo tD e iniciarse el periodo tΔ. La razón para emplear una palanca retráctil, en vez de una permanente, fue poder evaluar por separado los efectos de la frecuencia de entregas de agua de los de las frecuencias de respuestas asociadas a su entrega. Con la palanca retráctil solo pudo ocurrir una respuesta por entrega de agua en tD. Los resultados obtenidos en ambos experimentos sugirieron que el número total de respuestas en la sesión dependió, en primer término, de la frecuencia de respuestas asociadas a las entregas locales de agua y, complementariamente, del número total de entregas de agua, al margen de su localización al inicio o al final de la sesión, y de que una gran parte de ellas fuera de carácter no contingente a la respuesta. En la medida en que se observó una mayor frecuencia total de respuesta en el Experimento 1, en el que no se empleó palanca retráctil, que en el Experimento 2, es plausible suponer que la frecuencia total observada en la sesión fue una función primordial de las frecuencias locales de respuesta bajo densidades diferenciales de entrega de agua.
Se planeó un experimento para evaluar el efecto de la entrega de agua predeterminada, en distintos ciclos de un programa temporal, sobre la frecuencia total de respuestas en la sesión experimental. Para el efecto se entregó el agua en veinte ciclos repartidos de manera regular en toda la sesión, con el fin de determinar si el incremento en la frecuencia de la respuesta era una función de una proporción mínima de entregas por sesión, independientemente de su localización. A diferencia de otros estudios, el estimulo correlacionado con tD se mantuvo durante todo el periodo para evitar diferencias en la duración del estímulo entre las entregas contingentes y no contingentes de agua.
MÉTODO
Sujetos
Se emplearon 8 ratas albinas macho (cepa Wistar) de seis a doce meses de edad, experimentalmente ingenuas, que fueron asignadas aleatoriamente a dos grupos de cuatro ratas cada uno. Las ratas se mantuvieron bajo una privación de agua de 22 horas diarias, con acceso libre a alimento sólido (Nutricubos de Purina) en sus cajas habitación. Después de cada sesión experimental de una hora de duración, las ratas tuvieron acceso libre al agua por una hora. Las sesiones experimentales se realizaron seis días a la semana de las 10 a las 12 horas.
Aparatos
Se emplearon cuatro cámaras de condicionamiento operante de construcción propia (30 cm de largo por 25.5 cm de ancho y 32 cm de altura), colocadas en cubículos atenuadores de sonido y cada una con extractores de aire que funcionaron como ruido blanco. Cada caja disponía de un dispensador de líquido localizado en el centro del panel operativo. A 2.6 cm a la izquierda del dispensador se ubicaba una palanca que sobresalía 2 cm y requería de 0.13 N de fuerza para operar. Un foco blanco de 28-v, ubicado en la parte superior del panel operativo, procuró la iluminación general de la cámara durante toda la sesión, excepto durante las entregas de agua. En cada una de las entregas, el dispensador de líquido se iluminaba mediante el encendido de un foco blanco de 28-v, simultáneamente al apagado de la luz general de la cámara. En cada entrega se proporcionó una gota de agua de 0.01 cc, con una disponibilidad de 3 s. Se colocó una tecla translúcida 6.5 cm arriba de la palanca operativa, y detrás de ella se montaron dos focos de 5 w; uno que proyectaba luz verde y otro luz roja, cada una asociada con uno de los subciclos tD y tΔ, respectivamente. La programación y registro de los eventos se realizó mediante un sistema de cómputo AT386 PC-Turbo, empleando una interfase Paraport. El programa se diseñó utilizando Turbo Pascal 7.0.
Procedimiento
Las ratas fueron entrenadas a presionar la palanca mediante moldeamiento directo, hasta que obtuvieron 100 entregas de agua consecutivas en la sesión. Al término del moldeamiento, todas las ratas se expusieron a tres sesiones de reforzamiento continuo (100 entregas de agua), y después cada grupo fue sometido a un programa temporal de estímulos (Schoenfeld y Cole, 1972) con parámetros especiales en lo que toca a la distribución de la entrega de agua no contingente. El programa temporal empleado para todas las ratas consistió en un ciclo repetitivo T de 60 s, dividido en dos subciclos, tD y tΔ, de 30 s de duración cada uno. El subciclo tD se correlacionó con una luz verde y el subciclo tΔ con una luz roja. En ambos subciclos cada luz permaneció durante todo el periodo, incluso cuando se hubiera emitido una respuesta que produjera la entrega de agua en tD. La probabilidad de entrega de agua fue de 1.0 en tD y de 0.0 en tΔ, mientras el valor de T fue de 0.5 a lo largo del experimento. Cada sesión estuvo compuesta de 60 ciclos T de 60 s.
El experimento consistió de cinco fases, tres fases de 30 sesiones y dos fases de 10 sesiones, para un total de 110 sesiones. En tres de las fases se entregó el agua contingente o no contingente de manera regular cada tercer ciclo a lo largo de la sesión, mientras que en dos de las fases la entrega contingente o no contingente del agua se concentró en los primeros o últimos 20 ciclos de cada sesión. En ambos grupos de ratas, en las fases 1, 3 y 5 se seleccionaron 20 ciclos terciados (los ciclos 1, 4, 7, 10, 13, 16, 19, 22, 25, 28, 31, 34, 37, 40, 43, 46, 49, 52, 55, y 58) en los que la primera respuesta en tD producía la entrega del agua. De no darse respuesta alguna, el agua se entregaba de manera no contingente al final del subciclo tD. En dichos ciclos, para las ratas del Grupo 2 se retrajo la palanca solo después de la primera respuesta y se volvió a reintroducir al iniciar tΔ, mientras que para las ratas del Grupo 1 la palanca permaneció disponible todo el tiempo. En el resto de los ciclos, la primera respuesta en tD era seguida por la entrega de agua y la palanca permaneció disponible de manera que se podía continuar respondiendo en ambos grupos. En las fases 2 y 4, cada uno de los grupos se subdividió en dos, de modo que dos ratas recibieron el agua de manera contingente (al dar respuesta) o no contingente (al final de tD) en los primeros 20 ciclos (Ratas 1, 2, 5 y 6), y otras dos Ratas 3, 4, 7 y 8) en los últimos 20 ciclos de cada sesión. Al igual que en las otras fases, en el Grupo 2 la palanca se retrajo después de la primera respuesta en dichos bloques de ciclos (primeros o últimos veinte) mientras que en el Grupo 1 la palanca se mantuvo disponible.
RESULTADOS
La Figura 1 muestra el número total de respuestas en los subciclos tD y tΔ y el número total de entregas de agua por sesión para cada rata de ambos grupos. Los datos del Grupo 1 se muestran a la izquierda de la figura, mientras que los del Grupo 2 del lado derecho. Los datos están representados en una escala logarítmica por las variaciones en el número de respuestas observadas especial, pero no exclusivamente, en el subciclo tΔ. En el caso de las entregas de agua tuvieron un límite máximo de 60 por sesión.
En las ratas del Grupo 1, con palanca normal durante el subciclo tD de los ciclos seleccionados para entrega contingente o no contingente de agua ("forzosa"), se observaron ejecuciones poco variables entre sesiones. Con excepción de la Rata 1, en las demás ratas el número de respuestas en tD fue similar al número de entregas de agua, con una mayor ocurrencia de respuestas, en la mayoría de las fases, en el subciclo tΔ que en el subciclo tD. La excepción fue la Fase 5, en la que el número de respuestas en ambos subciclos tendió a ser similar o mayor en el subciclo tD. Las ratas 2, 3 y 4 obtuvieron la mayor parte de las entregas de agua, tanto en las sesiones en que su entrega forzosa se distribuía a lo largo de la sesión como en aquellas en las que se concentraba en los ciclos iniciales o terminales. La rata 1, en cambio, obtuvo solo una pequeña porción de las entregas de agua no forzosas, como se destaca especialmente en la Fase 3 en que no respondió o emitió escaso número de respuestas en el subciclo tD.
En las ratas del Grupo 2, con palanca retráctil, en la primera fase, con las entregas de agua "forzosas" programadas cada tercer ciclo, se observó que las Ratas 6 y 7 obtuvieron prácticamente todas las entregas de agua a partir de la cuarta o quinta sesión, mientras que las Ratas 5 y 8 oscilaron entre 30 y 60 entregas de agua por sesión durante esta fase, especialmente la Rata 8 que obtuvo la totalidad de entregas en muy pocas sesiones. En lo que respecta al número de respuestas, en general, para todas las ratas ocurrieron más respuestas en el subciclo tΔ que en el subciclo tD. En el caso de las ratas 6 y 7, el número de respuestas en el subciclo tD prácticamente igualó el número de entregas de agua, con poca variación en el número de respuestas en el subciclo tΔ. En cambio, en las ratas 5 y 8, se observó mayor variabilidad en la distribución de respuestas en ambos periodos del ciclo, en ocasiones con un número de respuestas en cada uno de los dos periodos inferior al número de entregas de agua. Con la presentación de las entregas forzadas en los primeros o últimos 20 ciclos de cada sesión en la Fase 2, y la posterior replicación de la primera fase en dos ocasiones (Fases 3 y 5) y de esta fase (Fase 4), se observó en todas las ratas mayor variabilidad en la entrega de agua y un menor número de respuestas en ambos subciclos, pero especialmente en el subciclo tΔ. Este efecto en cada rata dependió de la disminución observada en la Fase 2.
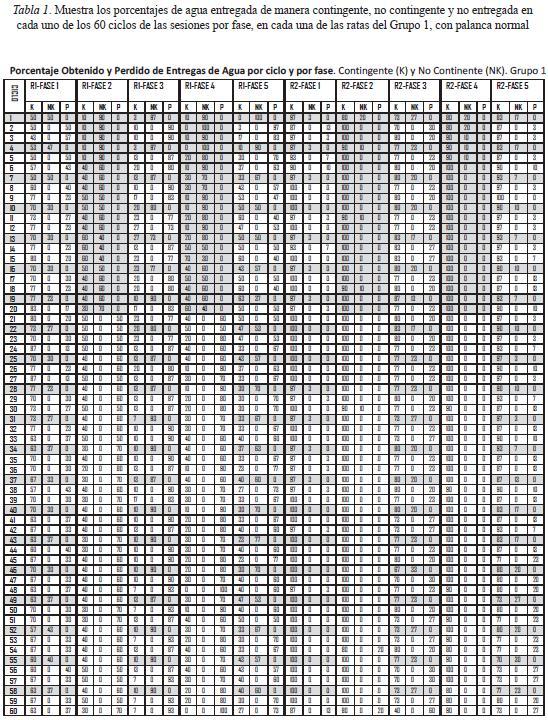
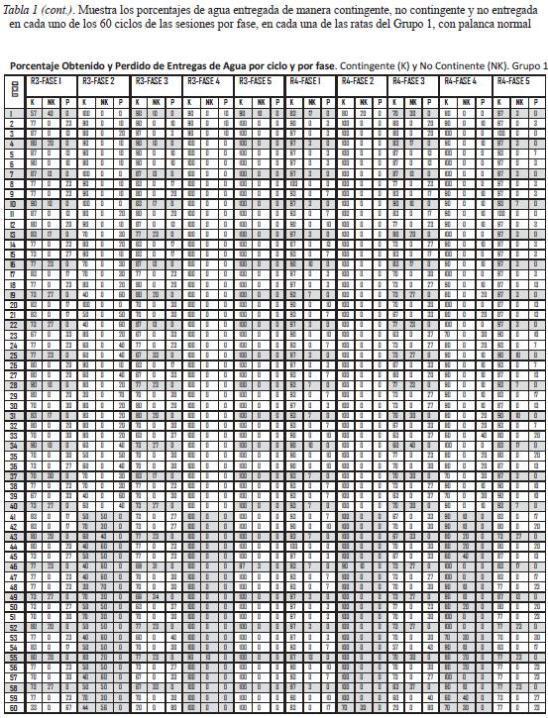
La Tabla 1 muestra, para cada fase, los porcentajes de agua entregada de manera contingente y no contingente, así como de agua no obtenida en cada uno de los 60 ciclos para cada una de las ratas del Grupo 1, que respondieron con palanca normal. Las ratas 1 y 2, expuestas a la programación de un bloque de entregas de agua no contingente en los primeros 20 ciclos de la sesión, mostraron los siguientes rangos de porcentaje de obtención de agua contingente: la rata 1, en la Fase 1, de 83 a 50%, en la Fase 2 de 50 a 10%, en la Fase 3 de 23 a 3%, en la Fase 4 de 50 a 0%, y en la Fase 5 de 63 a 0%; la rata 2, en la Fase 1 de 100 a 87%, en la Fase 2 de 100 a 80%, en la Fase 3 de 80 a 40%, en la Fase 4 de 100 a 80%, y en la Fase 5 de 100 a 73%. Esta última rata, a diferencia de la rata 1, obtuvo una gran cantidad de entregas de agua contingentes en los ciclos programados para entregarla también de manera no contingente al final del subciclo. Las ratas 3 y 4, que se expusieron a la programación del bloque de entregas de agua no contingente en los últimos 20 ciclos de la sesión, mostraron los siguiente rangos de porcentaje de obtención de agua contingente: la rata 3, en la Fase 1 de 83 a 57%, en la Fase 2 de 100 a 40%, en la Fase 3 de 90 a 63%, en la Fase 4 de 100 a 90%, y en la Fase 5 de 100 a 80; la rata 4, en la Fase 1 de 97 a 83%, en la Fase 2 de 100 a 70%, en la Fase 3 de 90 a 20%, en la Fase 4 de 100 a 60%, y en la Fase 5 de 100 a 73%. Los porcentajes más bajos se dieron en el primer y/o último ciclo o en los ciclos finales con entregas de agua no contingente programadas.
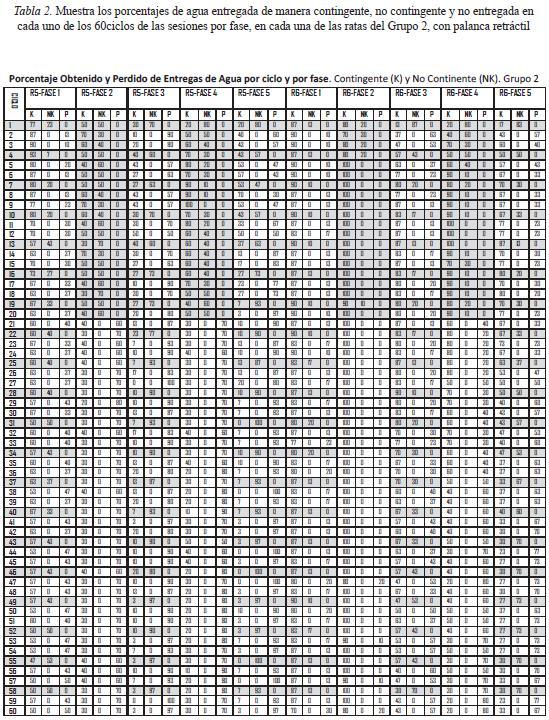
La Tabla 2 muestra el mismo tipo de datos para las ratas del Grupo 2 con palanca retráctil. En el caso de las Fases 1, 3 y 5, se muestran los datos de 30 sesiones, mientras que en las Fases 2 y 4 se muestran los datos de 10 sesiones. Las ratas 5 y 6, expuestas a la programación del bloque de 20 entregas de agua no contingente al inicio de la sesión, obtuvieron agua contingente en parte de los ciclos de dicho bloque. La rata 5 mostró los siguientes rangos de porcentaje de agua contingente obtenida: en la Fase 1 de 87 a 50%, en la Fase 2 de 70 a 50%, en la Fase 3 de 30 a 7%, en la Fase 4 de 100 a 20%, y en la Fase 5 de 70 a 3%. La rata 6 mostró los siguientes rangos: en la fase 1 de 90 a 70%, en la Fase 2 de 100 a 80%, en la Fase 3, de 80 a 13%, en la Fase 4 de 100 a 20%, y en la Fase 5 de 77 a 17%. Por su parte, las ratas 7 y 8 expuestas a la programación del bloque de 20 entregas no contingentes de agua en los últimos 20 ciclos de la sesión, mostraron los siguientes rangos de obtención de agua contingente: la rata 7, en la Fase 1 de 93 a 60%, en la Fase 2, de 100 a 60%, en la Fase 3 de 80 a 20%, en la Fase 4, de 100 a 10%, y en la Fase 5 de 83 a 10%; la rata 8, en la Fase 1 de 70 a 17%, en la Fase 2 de 100 a 10%, , en la Fase 3, de 79 a 23%, en la Fase 4, de 100 a 50%, y en la Fase 5 de 97 a 17%. En general, los porcentajes más bajos ocurrieron en los ciclos iniciales y/o finales de las sesiones, especialmente cuando se entregaba agua no contingente al final de los ciclos.
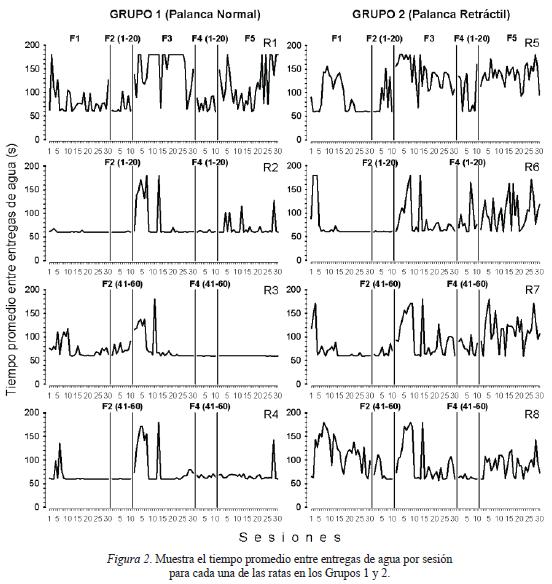
La Figura 2 muestra el tiempo promedio entre entregas de agua por sesión para cada una de las ratas de ambos grupos. En general, se observa mayor variación en los intervalos entre entregas de agua en las ratas del Grupo 2, con palanca retráctil, que en las ratas del Grupo 1, con palanca permanente, excepto en la rata 1 que también mostró gran variación. Aquellas fases, en ratas de ambos grupos, en las que se observan tiempos estables entre entregas de agua, el efecto se debe a la obtención de altos porcentajes de las entregas de agua programadas (en el rango del 80 al 100%).

DISCUSIÓN
En términos generales, los resultados de este estudio muestran desempeños globales diferentes de las ratas expuestas a una palanca normal y aquellas expuestas a una palanca retráctil solamente en los ciclos en que se programó de manera alternativa la entrega de agua no contingente a la respuesta, cuando no se había producido una respuesta en el subciclo tD. Las ratas expuestas a la palanca normal obtuvieron mayor cantidad de agua que las ratas expuestas a la palanca retráctil, y prácticamente perdieron pocas entregas, a diferencia de lo observado en otros estudios (Ribes & Torres, 1996; Ribes, Torres, Barrera % Mayoral, 1997; Ribes, Torres & Mayoral, 1999). Exceptuando a una rata (R1), las otras ratas del Grupo 1 obtuvieron en la mayoría de las sesiones las 60 entregas de agua programadas, mientras que en el Grupo 2, expuesto a la palanca retráctil, sólo en la primera y segunda fases dos de las cuatro ratas (R6 y R7) obtuvieron las 60 entregas de agua en la mayor parte de las sesiones. En el resto de las fases se observó –como en las primeras dos fases en las ratas 5 y 8- una disminución y variabilidad entre sesiones en el número de entregas de agua obtenidas. La entrega de agua libre o no contingente, en caso de no respuesta en los primeros o los últimos 20 ciclos de cada sesión, no tuvo efectos diferenciales aparentes, tal como se encontró en Ribes, Zepeda, Arenas y Mayoral (2007).
En este estudio no se observó una frecuencia mayor de respuesta en tΔ que en tD de manera sistemática, como es común en los programas temporales señalados con los valores de T y P aquí empleados (0.5 y 1.0, respectivamente). Sin embargo, las frecuencias altas de respuesta que tuvieron lugar en ambos grupos, y en distintas fases, ocurrieron usualmente en los subciclos tΔ. Como también se ha observado en otros estudios (Ribes & Torres, 1996), la cantidad obtenida de entregas de agua no dependió directamente de la frecuencia total de respuesta en el ciclo ni de la frecuencia de respuesta en el subciclo tD, como lo sugieren las ejecuciones de las ratas 1 y 4, principalmente. Los tiempos entre entregas de aguas fueron relativamente constantes en relación con el valor del ciclo T (60 s), con algunas excepciones, en tres de las ratas expuestas a la palanca normal. En cambio, en las ratas del Grupo 2, con palanca retráctil, sólo se observó cierta constancia en las primeras dos fases en dos de la ratas (6 y 7). En el resto de las fases y en las demás ratas se observó una gran variabilidad en los tiempos entre entregas de agua, con picos cercanos a los 200 s, de manera similar a lo que ocurrió con la rata 1.
Al examinar los subciclos tD y tΔ con respuesta en las distintas fases, se encontraron diferencias importantes en ambos grupos de ratas. En el grupo con la palanca normal, exceptuando a la rata 1, se observaron respuestas en todos los ciclos, obteniendo la mayor parte de la entregas de agua de manera contingente. Solo en los ciclos iniciales y finales dejaron de ocurrir respuestas, probablemente debido a los bloques de entrega no contingente de agua (en caso de no respuesta) en las Fases 2 y 4. Sin embargo, las ratas 2, 3 y 4 (y la rata 1 en la primera fase) respondieron en la mayoría de los ciclos en que había también programadas entregas no contingentes de agua. En cambio, las ratas del Grupo 2, con palanca retráctil, obtuvieron menos entregas contingentes de agua que las del Grupo 1, aunque el efecto también fue más notorio en los primeros y últimos ciclos de cada sesión, lo que puede deberse de igual manera a un efecto de los bloques iniciales o terminales de entregas no contingente de agua en las Fases 2 y 4. En un elevado porcentaje de los ciclos con entregas de agua no contingente programada, se observó entregas de agua contingente, aunque no equivalente al del Grupo 1, en el que en algunas fases el 100% de entregas de agua en un ciclo determinado por fase fue producido por la respuesta de la rata.
Los resultados de este experimento confirman que la frecuencia de respuesta en un programa temporal depende de la obtención de un porcentaje (en este caso la tercera parte) de las entregas de agua (o comida) programadas bajo un valor de probabilidad de 1.0 (Ribes, Zepeda, Arenas & Mayoral, 2007). Este valor quizá indica un umbral de consecuencias por bajo del cual la ejecución se torna poco efectiva, de manera semejante a los valores de T y P que Schoenfeld y Cole (1972) identificaron como transiciones entre las ejecuciones de "razón" y las de "extinción". A diferencia de los estudios clásicos con programas temporales, en este experimento se emplearon ciclos T de un minuto con valores de P y T independientes, por lo que el valor de P "límite" podría ser mayor que el encontrado en estudios con ciclos más cortos y subciclos tD breves.
También se confirmaría la propuesta de que la frecuencia de respuesta en un programa temporal depende del contacto global en las sesiones iniciales con una densidad mínima de entregas de agua (o comida), pues de lo contrario se darán frecuencias bajas de respuesta, localizadas principalmente en tΔ, y con un gran porcentaje de entregas de agua (o comida) perdidas. En un principio se consideró que la frecuencia de respuesta en cada sesión y en el conjunto del experimento probablemente dependía del contacto inicial con las entregas de agua (o comida) en los primeros quince o veinte ciclos (Ribes, Mayoral, Torres & Ibañez, 2000; Ribes,Torres & Mayoral, 2002). Sin embargo, tal como se encontró en un estudio previo (Ribes et al. 2007), la entrega no contingente de agua en los primeros y últimos 20 ciclos de la sesión tuvo efectos equivalentes, por lo que el factor determinante es la densidad relativa de contactos durante la sesión completa y no en su primera parte. En el presente estudio, la distribución de las entregas de agua no contingentes, en caso de no respuesta, de manera proporcional a lo largo de la sesión, produjo un mejor desempeño que el observado en el estudio antes mencionado, especialmente en lo que corresponde al porcentaje de entregas de agua perdidas, la variación en la frecuencia de respuesta entre sesiones y la constancia en el tiempo entre entregas de agua.
Cuando se analiza el porcentaje de entregas contingentes y no contingentes en los primeros 20 ciclos de la primera fase para cada una de las ratas, los datos sugieren que la constancia y efectividad del desempeño depende del número de entregas contingentes obtenidas, como lo ilustran los casos negativos de las ratas 1 y 8. Sin embargo, esto no es suficiente para entender el por qué de la disminución de la frecuencia de respuesta y de entregas contingentes en las ratas restantes del Grupo 2, con palanca retráctil, a partir de la segunda o tercera fases. Una posibilidad, apoyada en la frecuencia de respuestas durante el subciclo tD, cuando se respondió en dicho periodo en que estaba programada alternativamente una entrega de agua no contingente, hubo un intervalo sin palanca hasta el inicio del subciclo tΔ, lo que impidió que las ratas respondieran, y que dicho responder fuera temporalmente contiguo a la entrega de agua obtenida. Esto no ocurrió en el Grupo 1, con palanca normal, en el que correlacionaron varias respuestas con la entrega de agua. En este grupo hubo mayor número de respuestas en tD que entregas de agua por sesión, al contrario de lo que ocurrió en el Grupo 2, especialmente a partir de la segunda y tercera fases. Las densidades locales de entrega de agua se correlacionaron con densidades mayores de respuesta en tD en el grupo con palanca normal que en el grupo con palanca retráctil, lo que apoya la interpretación previa (Ribes et al, 2007) acerca de interacciones bidireccionales entre la densidad de entregas de agua y la densidad de respuestas correlacionadas (Catania & Reynolds, 1968; Reynolds & Limpo. 1968).
Finalmente, en este experimento, de manera similar al estudio previo examinado (Ribes et al, 2007), no se observaron frecuencias más elevadas de respuesta en tΔ de manera sistemática. A partir de la fase 3, en muchas de las sesiones en las ratas de ambos grupos, no ocurrieron respuestas durante los subciclos tΔ. Sin embargo, mientras que en el Grupo 1 este efecto tuvo lugar con respuestas ocurriendo prácticamente en todos los subciclos tD, en el Grupo 2 se vio acompañado de la no ocurrencia de respuestas también en ese subciclo. El desempeño observado en las ratas del Grupo 2 durante las dos últimas fases, sería probablemente el primer hallazgo de una "discriminación" ideal con programas temporales.
Se puede concluir que este experimento sugiere influencias bidireccionales entre las densidades locales de entrega de agua y la frecuencia de respuesta, relaciones que, sin embargo, no dependen de la ubicación temporal de la entrega de agua durante la sesión. Por ello es importante examinar en un futuro las interacciones moleculares bidireccionales que tienen lugar al interior de los distintos ciclos durante la sesión, pues tienen que ser determinantes de las transiciones particulares que resultan del proceso (Leslie, 1981).
REFERENCIAS
Catania, A.C., & Reynolds, G.S. (1968). A quantitative analysis of the responding maintained by interval schedules of reinforcement. Journal of the Experimental Analysis of Behavior, 11, 327-383. [ Links ]
Ferster, C.B. , & Skinner, B.F. (1957). Schedules of Reinforcement. New York: Appleton Century Crofts. [ Links ]
Leslie, J. C. (1981). Effects of variations in local reinforcement rate on local response rate in variable interval schedules. Journal of the Experimental Analysis of Behavior, 35, 45-53. [ Links ]
Reynolds, G.S., & Limpo, A. (1968). On some cause of behavioral contrast. Journal of the Experimental Analysis of Behavior, 11, 543-547. [ Links ]
Ribes, E., & Torres, C. (1996). Efectos de la variación de la probabilidad de reforzamiento correlacionada con dos estímulos neutros en un programa definido temporalmente. Revista Mexicana de Análisis de la Conducta, 22,41-78. [ Links ]
Ribes, E., Torres, C.,& Mayoral, A. (1999). Efectos de la ausencia y presencia de estímulos correlacionados con distintas probabilidades de reforzamiento y con extinción en programas definidos temporalmente. Revista Mexicana de Análisis de la Conducta, 26, 327-354. [ Links ]
Ribes, E., Torres, C., & Mayoral, A. (2000). Señalización no diferencial de distintas probabilidades de entrega de agua en dos subciclos de un programa definido temporalmente. Acta Comportamentalia, 8, 5-21. [ Links ]
Ribes, E., Torres, C., & Mayoral, A. (2002). Extended exposure to a discriminated, limited-hold temporal schedule does not produce stimulus control. Behavioural Processes, 59, 131-146. [ Links ]
Ribes, E., Torres, C., Barrera, A.,& Mayoral, A. (1997). Control discriminativo con estímulos compuestos en programas temporales de reforzamiento. Revista Mexicana de Análisis de la Conducta, 23, 25-54. [ Links ]
Ribes, E., Mayoral, A., Torres, C., & Ibañez, F.J. (2000). Effects of auditory stimuli correlated with different probabilities of water delivery in a limited-hold temporal schedule. Behavioural Processes, 52,49-59. [ Links ]
Ribes, E., Zepeda, I., Arenas, S. & Mayoral, A. (2007). Efecto de la densidad local de entrega de agua en la frecuencia total de respuesta en un programa temporal señalado. Revista Mexicana de Análisis de la Conducta, 33, 139-166. [ Links ]
Schoenfeld, W.N., & Cole, B.K. Stimulus schedules: the T-t systems. Nueva York: Harper & Row Publishers. [ Links ]
Received: September, 17, 2011
Accepted: November, 29, 2011
Dirigir correspondencia al primer autor a: Centro de Estudios e Investigaciones en Conocimiento y Aprendizaje Humano. Av. Orizaba No. 203 (Fraccionamiento Veracruz). Xalapa, Veracruz. México, C. P. 91020 o al correo electrónico: eribes@uv.mx
