










Services on Demand
article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkActa Comportamentalia
Print version ISSN 0188-8145
Acta comport. vol.20 no.3 Guadalajara 2012
ARTÍCULOS
El consumo de comida en ratas controlado por un estímulo neutral
Food-intake in rats controlled by a neutral stimulus
Varsovia Hernández; Carlos A. Bruner1
Universidad Nacional Autónoma de México (México)
RESUMEN
Un estímulo neutral contiguo con un siguiente periodo de acceso a la comida incrementa la ingesta de comida en ratas relativo a cuando el estímulo se presenta ya sea contiguo al acceso precedente o aleatoriamente durante el intervalo entre accesos. En el presente experimento se determinaron los efectos de variar la ubicación temporal del estímulo durante el periodo entre accesos. Tres ratas vivieron 24/7 en sus respectivas cámaras experimentales y se les dieron periodos señalados de acceso a la comida de 10 minutos cada 160 minutos. Durante la línea base no se presentó ningún estímulo durante los periodos entre accesos. En condiciones subsecuentes, un estímulo de 5 minutos fue presentado en una ubicación temporal fija durante el periodo entre accesos, ya fuera 5, 10, 20, 40, 80 o 160 minutos antes del siguiente periodo de acceso. La ubicación de 5 minutos incrementó la ingesta relativo a la línea base. Alargar el periodo entre el estímulo y el acceso controló disminuciones correspondientes en la ingesta de comida incluso por debajo de la línea base. Se discuten las implicaciones de controlar la ingesta de comida al introducir un estímulo neutral en diferentes ubicaciones del periodo entre accesos.
Palabras clave: estímulo neutral, intervalo de acceso a la comida, intervalo entre accesos a la comida, ingesta de comida, ratas.
ABSTRACT
A neutral stimulus contiguous with a subsequent access-to-food period increase food intake in rats relative to when the stimulus is presented either contiguous to the preceding access or randomly during the interaccess interval. The present experiment determined the effects of varying the temporal placement of the stimulus during the inter-access period. Three rats lived 24/7 in their respective experimental chambers and were given signaled 10-minute access-to-food periods every 160 minutes. During baseline no stimulus was present during the inter-access periods. On subsequent conditions, a 5 minute stimulus was placed on a fixed temporal location during the inter-access period, either 5, 10, 20, 40, 80 or 160 minutes before the next access period. The 5-minute placement increased intake relative to baseline. Lengthening the stimulusto- access period controlled corresponding decreases in food intake even below baseline. The implications of controlling food intake by intruding a neutral stimulus at different locations of the inter-access period are discussed.
Keywords: neutral stimulus, access-to-food interval, inter-access-to-food interval, food-intake, rats.
En ratas privadas de comida, presentar un estímulo neutral justo antes de un periodo de acceso a la comida resulta en que coman más que cuando no se presenta el estímulo. A pesar de ser un fenómeno de considerable interés debido a sus posibles aplicaciones en el control de la ingesta, ha sido estudiado relativamente poco. Calvin, Bicknell y Sperling (1953) mostraron este efecto en ratas que habían comido durante una primera estancia en una cámara experimental y que al volverlas a colocar en la misma cámara comieron más que otras ratas a las que no se les dio comida en su primera estancia en la misma cámara experimental. A pesar de que Calvin et al. esperaban que comer dotaría a la cámara con la función de un reforzador condicionado y que éste disminuiría la pulsión por comer en el acceso siguiente a la manera de una pre-comida, sus resultados mostraron lo contrario. Diferente a su hipótesis original, Calvin et al. concluyeron que la presentación de un reforzador condicionado previo al acceso a la comida no disminuye la pulsión de hambre (véase también Valle, 1968).
En otro estudio, Zamble (1973) usó una misma cámara experimental para presentar repetidamente un estímulo contiguo a periodos de acceso a la comida. Estas ratas comieron más que otras a las que se les presentó el estímulo de manera aleatoria durante el periodo entre accesos sucesivos (Experimento 1) o al final del periodo de acceso anterior (Experimento 2). Zamble concluyó que un estímulo contiguo al acceso a la comida puede adquirir la función de una pulsión condicionada y así controlar un mayor consumo. En un estudio posterior, Lovibond (1980) probó la hipótesis de que un estímulo que precede al acceso a la comida podría evocar diversas conductas preparatorias a comer en ratas, como la búsqueda de comida y que éstas inducirían un mayor consumo (cf. Konorsky, 1967). En un primer experimento, encontró que introducir a las ratas en una cámara experimental 5 min antes de darles comida durante una primera sesión controló un mayor consumo durante sesiones posteriores en la misma cámara en comparación con la ingesta en una cámara en la que no se les entregó comida en la primera sesión. En un segundo experimento encontró que presentaciones repetidas de un estímulo contiguo con comida en una misma cámara experimental controlaron un mayor consumo que la presentación de un estímulo diferente al anterior o ningún estímulo. Lovibond concluyó que el estímulo contiguo a la comida preparó a las ratas para la posterior ingesta del alimento.
A pesar de que en los trabajos descritos arriba se usaron procedimientos ostensiblemente diferentes, tienen algunos aspectos en común. Uno de estos es que todos se concentraron en el mismo fenómeno; i.e., el aumento en el consumo de comida en función de presentar estímulos contiguos al acceso a la comida. Otro aspecto en común es que todos intentaron probar que el estímulo que precede a la comida afecta a la pulsión por comer, ya sea como un sustituto de la comida (e.g., Calvin, Bicknell & Sperling, 1953) o bien porque adquiere la función de un estímulo condicionado que evoca la pulsión por comer. Es posible que la táctica de probar hipótesis, característica de la teoría de la motivación, haya distraído la atención de los investigadores del estudio de las variables independientes que en primer lugar controlan la ocurrencia del fenómeno.
Independientemente del aparente origen de este hallazgo en la teoría de la motivación, es claro que el conocimiento sobre la conducta de comer se encuentra repartido en el estudio de diferentes procesos psicológicos (Bruner, 2010). Un ejemplo notable relativo al efecto de estímulos neutrales que preceden a comer proviene del área de psicología social, específicamente del tema de facilitación social. En un experimento realizado por de Castro y Brewer (1992), se mostró que el número de calorías que consume una persona durante una comida es una función creciente del número de comensales en la mesa. Es muy probable que este descubrimiento involucre esencialmente las mismas variables que los estudios mencionados anteriormente con ratas y sirve para enfatizar el argumento concerniente a la importancia de estudiar a fondo las variables independientes que controlan un fenómeno. Una vez que se hace un intento por ignorar el origen de un fenómeno en algún apartado convencional de la psicología con algún enfoque particular y el investigador se concentra en las operaciones reales que controlan al fenómeno, es posible indagar con mayor profundidad sus variables independientes. En el caso del aumento en el consumo de comida en función de estímulos contiguos al acceso, es posible ver que la contigüidad entre el estímulo y la comida representa solamente un extremo de un continuo de operaciones consistentes en variar la ubicación temporal del estímulo en relación a la comida. En el estudio de Zamble (1973) descrito con anterioridad, se varió la relación temporal entre el estímulo y el acceso a la comida como un control del efecto de su contigüidad. En su Experimento 1 el estímulo neutral se presentó al azar durante el intervalo entre accesos y en su Experimento 2, el estímulo se presentó en contigüidad con la comida precedente. La contigüidad del estímulo con la siguiente comida o con la precedente representan de hecho los dos extremos de un continuo definido por el resto de las ubicaciones intermedias del estímulo entre dos comidas sucesivas. La variación sistemática de la ubicación temporal del estímulo en relación a los periodos de acceso a la comida es una manipulación que carece de precedentes en el estudio de la conducta de comer. Sin embargo, es una operación bien estudiada en condicionamiento, conocida como el "paradigma del estímulo intrusivo" (Farmer & Schoenfeld, 1966a). El propósito del presente trabajo fue determinar la cantidad de comida consumida durante los periodos de acceso en función de variar la ubicación temporal de un estímulo neutral entre accesos sucesivos. Dado que aparte de la condición de contigüidad "hacia atrás" en el estudio de Zamble no existe información sobre los posibles efectos de este procedimiento, un objetivo inmediato fue el de determinar el grado de separación entre el estimulo y la comida que aún permite notar un aumento en el consumo.
MÉTODO
Sujetos
Se utilizaron tres ratas Wistar macho de cuatro meses de edad y experimentalmente ingenuas. Durante todo el estudio los sujetos vivieron en las cámaras experimentales. El acceso al agua se mantuvo libre durante todo el estudio mientras que el tiempo de acceso a la comida fue de 80 minutos por día.
Aparatos
Se utilizaron tres cámaras experimentales de 49 cm de ancho x 40 cm de altura x 38 cm de profundidad. En el interior de cada cámara se instaló un panel frontal en el que se colocó una palanca (Modelo ENV-110RM, Med Associates Inc.) sensible a una fuerza de 0.15 N. La palanca se ubicó a un altura de 6.5 cm respecto del piso de la caja y a 2 cm respecto de la pared izquierda de la caja. A la derecha de la palanca se colocó un comedero de metal el cual se ubicó 3.5 cm por detrás del nivel del panel. El comedero se ubicó a 3.5 cm respecto del piso de la caja. En la parte posterior del comedero se conectó un dispensador de comida (Modelo ENV-203IR, Med Associates Inc.) que sirvió para entregar bolitas de comida de 25 mg. Las bolitas de comida se fabricaron remoldeando comida para ratas de la marca Zeigler. En la parte posterior del panel frontal se colocó una botella con agua. Al lengüetear la pipeta de la botella los sujetos tenían acceso al agua. La pipeta se colocó a 0.7 cm detrás del panel y se ubicó a 19.5 cm a la derecha de la palanca y a 8 cm respecto del piso de la caja. En el panel frontal también se colocó un generador de sonido (ENV-223AM) ubicado 22 cm por arriba del comedero. El generador produjo un tono de 60 dB. Para simular el ciclo de luz permanecieron encendidos dos focos de 28 V los cuales se ubicaron a 28 cm respecto del piso de la caja y estaban separados entre sí 18 cm. Cada caja estaba equipada con un ventilador que facilitó la circulación de aire y también sirvió para enmascarar ruidos ajenos a la investigación. El registro y control de los eventos experimentales se realizó por medio de una computadora con software MED-PC IV acoplada a una interfase Med-Associates Inc (Modelo SG-503), la cual se ubicó en un cuarto adyacente al lugar en el que se colocó a las ratas.
Procedimiento
Establecimiento de las presiones a la palanca
Con el fin de establecer la respuesta de presión a la palanca, se expuso directamente a los sujetos a un programa de reforzamiento regular (RF1) conforme al cual por cada presión a la palanca se entregó una bolita de comida. Este programa estuvo vigente durante cinco días. No se dio ningún entrenamiento preliminar a los sujetos para acercarse al comedero. Todas las sesiones tuvieron una duración de 22 horas con 40 minutos por lo que los sujetos vivían en las cámaras experimentales. El tiempo restante del día se ocupó en pesar y rellenar las botellas con agua, verificar el funcionamiento de los dispensadores de comida, rellenarlos y limpiar las cámaras experimentales. Durante todas las fases, la mitad del tiempo que los sujetos se encontraron en la cámara experimental estuvo vigente el periodo de luz y la otra mitad el periodo de oscuridad.
En ninguna fase del estudio se encontraron residuos de comida en los comederos o en el piso de las cámaras experimentales.
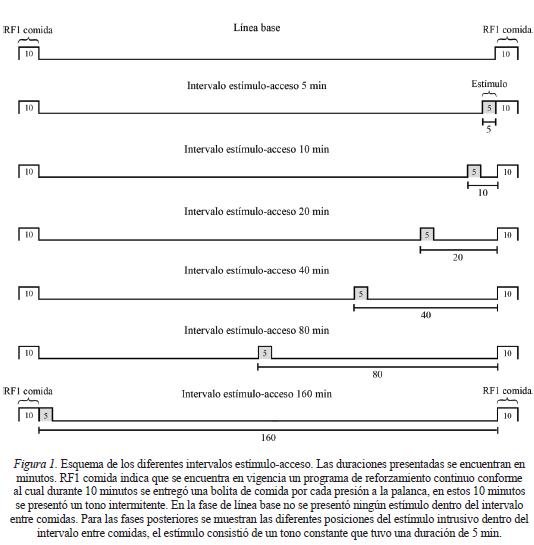
Línea base
En la Figura 1 se muestra un esquema del procedimiento. En cada día se programaron ocho periodos de acceso a la comida con duración de 10 minutos. Cada periodo de acceso se separó por un periodo sin acceso de 160 minutos. Los periodos de acceso a la comida fueron señalados con presentaciones de un tono intermitente que inició y terminó al mismo tiempo que el acceso a la comida. El tono intermitente consistió en el encendido y apagado del generador de sonidos cada segundo. Durante el periodo de acceso los sujetos obtuvieron una bolita de comida por cada presión a la palanca. La mitad de los periodos de acceso a la comida y de los periodos sin acceso se programaron en el periodo de luz y la otra mitad en el de oscuridad. Debido a que los periodos de acceso a la comida fueron ocho, el tiempo total de disponibilidad de comida fue de 80 minutos, el cual es el tiempo promedio que los sujetos con acceso libre al alimento ocupan para consumir comida en 24 horas (cf. Díaz & Bruner, 2007). El acceso a la botella con agua fue de manera libre. Durante toda la sesión experimental se registraron las presiones a la palanca. La línea base se mantuvo durante 30 sesiones.
Introducción del estímulo
Manteniendo constantes las mismas condiciones de la línea base, en diferentes fases se introdujo un tono continuo en una ubicación fija dentro de cada periodo sin acceso a la comida. El tono continuo siempre tuvo una duración de 5 minutos. En la primera fase se presentó el tono continuo durante los 5 minutos previos al periodo de acceso, por lo que al final de los 5 minutos de presentación del tono dio inicio el periodo de acceso a la comida. En fases subsecuentes, el intervalo entre el inicio de la presentación del tono continuo y el inicio de la entrega de comida se alargó conforme a una progresión geométrica en 10, 20, 40, 80 y 160 minutos. Después de introducir al tono en la última posición, se redeterminó el efecto de presentarlo en el intervalo estímulo-comida de 40 minutos y en el de 5 minutos. Cada ubicación del tono continuo se mantuvo fija durante 30 sesiones.
RESULTADOS
Para los tres sujetos la respuesta de obtención de comida se estableció en la primera sesión de entrenamiento. Todos los resultados se muestran como el promedio de las últimas 10 sesiones de cada condición. En todas las figura se muestran las desviaciones estándar correspondientes a cada promedio. Debido a que el patrón de alimentación de las ratas es predominantemente nocturno (Díaz & Bruner, 2007; Siegal, 1961), en cada figura se muestran los resultados por separado para el periodo de luz y el de oscuridad. El Sujeto 1 murió a los 5 días de iniciada la redeterminación del intervalo entre el estímulo y la comida de 40 minutos, por lo que los datos que se muestran se calcularon con base en esas sesiones.
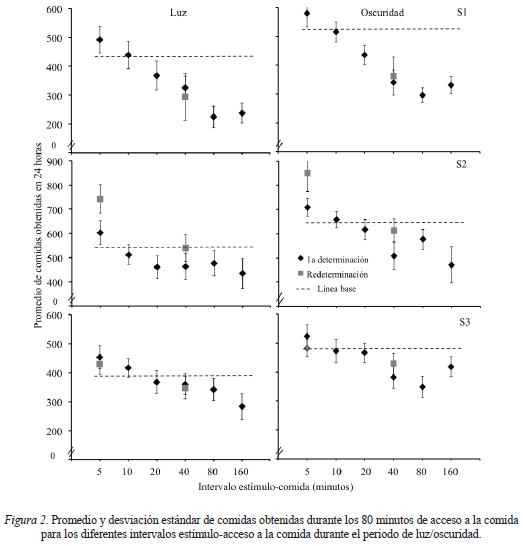
La principal variable dependiente en este estudio fue la cantidad de comida ingerida durante el periodo de acceso en función de presentar un estímulo entre accesos sucesivos y de los diferentes intervalos estímulo-acceso, por lo que en la Figura 2 se muestra el promedio de comidas obtenidas por cada sujeto durante los 80 minutos de acceso a la comida en cada fase del estudio. Con fines de comparación, en cada panel se presenta como una línea punteada el promedio de comidas obtenidas en la línea base en la que no se presentó el estímulo durante el periodo sin acceso. El eje de las ordenadas se muestra en una misma escala para los Sujetos 1 y 3 mientras que para el Sujeto 2 se muestra en una escala diferente. Durante el periodo de luz se encontró para los tres sujetos que la cantidad de comida consumida fue mayor para el intervalo entre la presentación del estímulo y el acceso a la comida de 5 minutos en comparación con la línea base. También para los tres sujetos se encontró que alargar el intervalo entre la presentación del estímulo y la comida controló disminuciones progresivas del número de comidas obtenidas con respecto al nivel de línea base. Al redeterminar el efecto de los intervalos estímulo-acceso de 40 y de 5 minutos se encontró que el número de comidas obtenidas regresó a niveles similares a los obtenidos la primera vez que se expuso a los sujetos a estas condiciones. Durante el periodo de oscuridad se encontró el mismo patrón en la ingesta que en el periodo de luz, con la diferencia de que la cantidad absoluta de comidas obtenidas fue mayor.
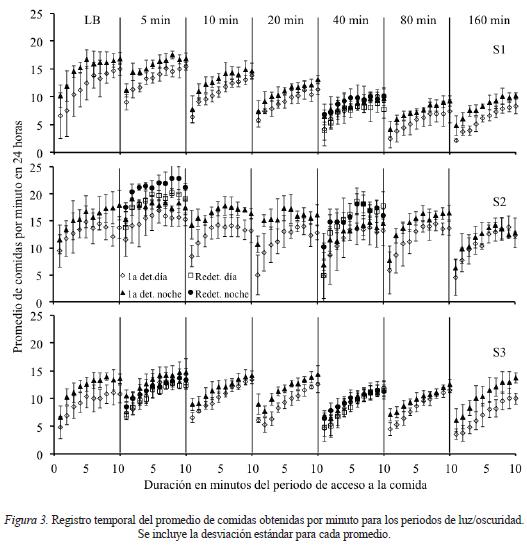
Un variable que ha sido descrita en estudios de motivación es el patrón temporal de ingesta durante el periodo de acceso a la comida. En la Figura 3 se muestra el registro temporal del promedio de comidas obtenidas por minuto durante la línea base y para los diferentes intervalos estímulo-acceso en el periodo de luz y oscuridad. La ingesta durante el periodo de luz se muestra con rombos blancos mientras que las redeterminaciones de los intervalos estímulo-comida de 5 y 40 minutos para este periodo se muestran con cuadros blancos. La ingesta durante el periodo de oscuridad se muestra con los triángulos negros y las redeterminaciones de los intervalos estímulo-comida de 5 y 40 minutos se muestran con círculos negros. Debido a que la única diferencia entre el patrón temporal de ingesta durante el periodo de luz y oscuridad fue que en el periodo de oscuridad la tasa global de comidas ingeridas fue más alta en comparación con el periodo de luz, se describen los datos en conjunto para ambos periodos. Para los tres sujetos se encontró que en todos los intervalos estímulo-comida la ingesta se distribuyó conforme a una curva negativamente acelerada. En el intervalo estímulo-comida de 5 minutos se encontró un nivel absoluto de la tasa de comidas obtenidas más alto en comparación con la línea base. También para los tres sujetos se encontró que al alargar el intervalo estímulo-comida la tasa global de comida consumida disminuyó pero no se modificó la forma de la curva de ingesta. En las redeterminaciones de los intervalos estímulo-acceso de 40 y de 5 minutos se encontró condiciones.
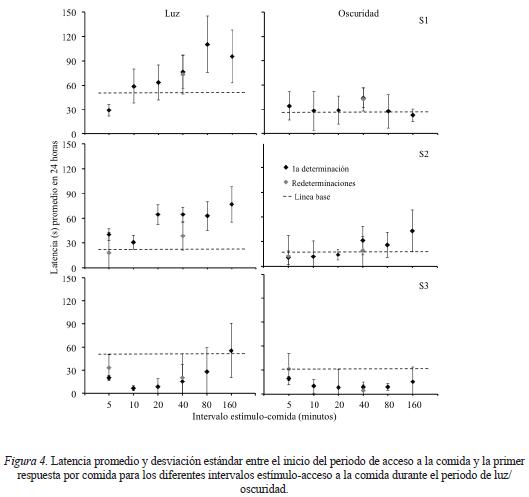
En algunos estudios se ha encontrado que la presentación de estímulos relacionados con comida tiene efectos sobre la latencia entre la presentación de la comida y el inicio del episodio de alimentación (e.g., Weingarten, 1983). En la Figura 4 se muestra para cada fase del estudio la latencia promedio entre el inicio del periodo de acceso a la comida y la primer respuesta de obtención de comida para el periodo de luz y el de oscuridad. Al igual que en la Figura 2, el nivel de línea base se muestra mediante una línea punteada.
Para obtener la latencia se descartaron los periodos de acceso en los que los sujetos no respondieron, este porcentaje como promedio de todas las fases equivalió a 2%, 1.3% y 1.5% para los Sujetos 1, 2 y 3, respectivamente. En el periodo de luz se encontró para los Sujetos 1 y 2 que alargar el intervalo estímulocomida controló incrementos progresivos en la latencia con respecto a la fase de línea base. Para el Sujeto 3, con excepción del intervalo estímulo-comida de 160 minutos, se encontró una duración menor de la latencia ante las diferentes ubicaciones del estímulo en comparación con la línea base aunque esta duración también incrementó al alargar el intervalo estímulo-comida. Redeterminar los intervalos estímulo-acceso de 5 y 40 minutos controló latencias similares a las obtenidas la primera vez que se expuso a los sujetos a estas condiciones. Durante el periodo de oscuridad las latencias para los tres sujetos fueron más cortas en comparación con el periodo de luz y no se encontró ningún patrón claro en los valores de esta variable en función de aumentar el intervalo estímulo-comida.
DISCUSIÓN
El propósito del presente trabajo fue determinar la cantidad de comida consumida durante los periodos de acceso en función de variar la ubicación temporal de un estímulo neutral entre accesos sucesivos. Se encontró que con respecto al nivel de línea base, presentar un estímulo en contigüidad con la siguiente comida controló aumentos en la cantidad de comida ingerida, mientras que alargar el intervalo entre la presentación del estímulo y el acceso a la comida controló disminuciones progresivas en la ingesta. Estos resultados fueron similares para el periodo de luz y oscuridad con la diferencia de que durante el periodo de oscuridad la cantidad de comida ingerida fue mayor. Si bien una posible crítica a los resultados de este estudio es que el efecto encontrado pudo deberse al orden de exposición de los tres sujetos a los diferentes valores de la variable independiente, el efecto de redeterminar la ubicación del estímulo en los intervalos estímulo-acceso de 40 y de 5 minutos descarta esa posibilidad. Al redeterminar los intervalos estímulo-acceso de 40 y de 5 minutos se encontraron resultados similares a los obtenidos la primera vez que se expuso a los sujetos a estas condiciones lo cual demuestra que, efectivamente, fue la ubicación del estímulo dentro del intervalo entre accesos lo que controló la ingesta y no el orden de presentación de éstos.
En diversos estudios se investigó el efecto de la presentación de un estímulo contiguo a un siguiente periodo de acceso a la comida o a un periodo de acceso antecedente al estímulo (e.g., Lovibond, 1980; Valle, 1968; Zamble, 1973), los cuales representan dos extremos de un continuo de ubicaciones del estímulo. Sin embargo, el procedimiento de variar de manera ordenada la ubicación de un estímulo neutral entre intervalos sucesivos utilizado en este estudio no tiene precedente en la literatura de conducta de comer. No obstante, en estudios de condicionamiento esta es una manipulación conocida como "paradigma del estímulo intrusivo" y ha sido utilizada en diversas investigaciones (e.g, Bruner, Gallardo, & Avila, 2002; Dews, 1966; Farmer & Schoenfeld, 1966a y b). La mayoría de éstas consistió en variar la ubicación temporal de un estímulo neutral entre un intervalo fijo de entregas de una sola bolita de comida. A diferencia de estos estudios, en la presente investigación los sujetos pudieron obtener más de una bolita de comida al final del periodo sin acceso por un tiempo similar al que ocupan consumiendo comida en condiciones de acceso irrestricto a la comida (Díaz & Bruner, 2007), lo cual permitió determinar el efecto de variar la ubicación temporal de los estímulos sobre la ingesta. Al utilizar este procedimiento se encontró que el grado de separación entre el inicio de la presentación del estímulo y la comida que aun permite notar un aumento en el consumo de comida se encuentra en el rango de 5 a 10 minutos.
El resultado referente al aumento en la cantidad de comida ingerida ante presentaciones contiguas del estímulo con el siguiente periodo de acceso apoya lo reportado en investigaciones anteriores (e.g., Lovibond, 1980; Zamble, 1973). No obstante, la disminución gradual en la ingesta en función de alargar el intervalo estímulo-comida es un hallazgo novedoso. Probablemente este resultado no se reportó previamente debido a que en estudios anteriores la ubicación del estímulo con respecto a la comida no se varió de manera sistemática, mientras que el procedimiento utilizado en este trabajo permitió ver que dependiendo de los diferentes valores que una misma variable independiente puede tomar es posible encontrar efectos sobre la cantidad de comida ingerida. En este caso, los extremos de la presentación del estímulo en contigüidad al siguiente periodo de acceso a la comida o posterior a ésta controlaron aumentos y disminuciones en la ingesta respectivamente, mientras que presentaciones del estímulo en ubicaciones intermedias controlaron ya sea una ingesta similar o menor a la de línea base.
El incremento en la ingesta encontrado en este trabajo cuando se presentó el estímulo cercano a la comida es consistente con estudios en los que se mostró que a menor intervalo entre la presentación de un estímulo y la comida la magnitud de la respuesta condicionada es mayor (e.g., Pavlov, 1927). En otros estudios también se mostró que la presentación de estímulos condicionados modifica no sólo la magnitud de la respuesta condicionada sino también la magnitud de la respuesta incondicionada (como una revisión véase Domjan, 2005). También es consistente con lo propuesto por algunos autores (Keller & Schoenfeld, 1950; Woods & Seeley, 2002) que han mencionado que la presentación de estímulos antes de la entrega de comida puede tener la función de preparar a los sujetos para el consumo por lo que podrían facilitar la ingesta. En cuanto a la disminución en la ingesta, también en algunos estudios de condicionamiento Pavloviano se reportó que es posible controlar disminuciones en la magnitud de la respuesta en función de presentar estímulos antecedentes (Kimble & Ost, 1961; Pavlov, 1927; Siegel & Domjan, 1971). No obstante, debido a que no es posible realizar una comparación directa de este estudio con estudios previos debido a la diferencia en los procedimientos utilizados, es importante que en estudios posteriores se indague más sobre el efecto de disminuir la ingesta al alargar el intervalo estímulo-comida.
Una ventaja más del procedimiento utilizado en este estudio es que permitió observar cambios ordenados no sólo en la cantidad de comida ingerida sino también en el patrón de ingesta y en la latencia entre el periodo de acceso a la comida y la primer respuesta de obtención de comida. Referente al patrón de ingesta, se encontró que siguió la forma de una curva negativamente acelerada en el que la presentación del estímulo con diferentes ubicaciones temporales respecto al periodo de acceso controló la tasa global de ingesta, pero no afectó la forma de la curva. La descripción del patrón de ingesta como el de una curva negativamente acelerada fue previamente reportada por Skinner (1932) en un estudio en el que expuso a ratas a un periodo de acceso a la comida en el cual por cada presión a una palanca recibían una bolita de comida. Skinner encontró que la frecuencia de comer fue alta en un inicio y disminuyó gradualmente, de manera similar a la encontrada en el presente trabajo.
En cuanto a la latencia entre el periodo de acceso y la primera respuesta de obtención de comida, se encontró que en general la latencia durante el intervalo estímulo-acceso de 5 minutos disminuyó respecto al nivel de línea base mientras que al incrementar el intervalo estímulo-comida la latencia aumentó durante el periodo de luz. En el periodo de oscuridad no se encontraron cambios sistemáticos en esta variable. El control de la duración de la latencia mediante la presentación de estímulos antecedentes al acceso a la comida también fue reportado en un estudio realizado por Weingarten (1983). Este autor encontró que la latencia para iniciar la ingesta fue más corta ante presentaciones de un estímulo apareado con comida que ante un estímulo que se presentaba a la mitad del intervalo entre accesos a la comida, aunque no reportó por separado esta variable para el periodo de luz y oscuridad. Debido a su carácter paramétrico y a la observación de la conducta en ambos periodos del día, en la presente investigación se encontró que la latencia en iniciar un periodo de alimentación, al igual que la ingesta de comida, depende de la posición del estímulo respecto a la comida pero únicamente para el periodo de luz.
Es posible que los resultados encontrados en este estudio no se hayan reportado en investigaciones anteriores porque partieron de un enfoque hipotético-deductivo dirigido a apoyar o refutar diferentes teorías motivacionales (e.g., Lovibond, 1980; Zamble, 1973), dejando de lado el estudio de las variables físicas de las cuales la conducta de alimentación es una función. Si bien los estudios mencionados previamente mostraron aspectos en común como el aumento en la ingesta en función de la presentación de un estímulo contiguo al acceso a la comida, al enfocarse en apoyar o refutar hipótesis se dificultó su integración en base a sus manipulaciones. En cambio, el paradigma utilizado en este trabajo permitió enfocarse en las variables que controlan la conducta y realizar manipulaciones ordenadas de éstas independientemente de su origen en teoría de la motivación.
Recientemente, Bruner (2010) propuso que sin tomar en cuenta el origen de los estudios sobre conducta de alimentación, un enfoque basado en la variable independiente resulta beneficioso en la tarea de sistematizar el conocimiento establecido sobre esta conducta. De esta manera, diferentes hallazgos en conducta alimentaria pueden verse como efectos continuos de diferentes valores de una misma variable independiente (Bruner, 2010) lo cual permite integrar hallazgos en conducta alimentaria que parecen separados. Por ejemplo, en estudios anteriores se presentó al estímulo en contigüidad con la siguiente comida o, a manera de control se presentó en contigüidad con la comida precedente (Zamble, 1973). En el presente estudio, al variar de manera ordenada la ubicación del estímulo durante el intervalo entre accesos a la comida, se incluyó toda la serie de ubicaciones realizadas en trabajos anteriores y se pudo ver que las diferencias en los resultados de cada estudio se pudieron deber al momento en que presentaron el estímulo dentro del intervalo entre accesos. Asimismo, los resultados de este estudio referentes a aumentos y disminuciones en la cantidad de comida consumida apoyan lo dicho por Bruner, ya que son una muestra clara de que manipulando los valores de las variables independientes, en este caso el intervalo estímulo-comida, se pueden integrar hallazgos que vistos de manera parcial podrían parecer producto de diferentes operaciones.
Basándonos exclusivamente en las operaciones que controlan la ingesta de comida en ratas y extrapolando los resultados a la conducta humana, es posible que las mismas manipulaciones que controlan la ingesta de comida en ratas también controlen la ingesta de comida en personas. Por ejemplo, de Castro y Brewer (1992) reportaron que la cantidad de comida que consume una persona en compañía de otros comensales es mayor que la que consume cuando está solo. Asimismo, en otros estudios se reportó que estímulos como la hora del día (Schachter & Gross, 1968) o la vista de alimentos (Cornell, Rodin & Weingarten, 1989) también controlan la ingesta de comida. En estos estudios se puede presumir que los comensales, la hora del día y la vista de los alimentos funcionaron como estímulos que a través de una historia de ocurrencias en contigüidad con la comida controlaron un aumento en el consumo de alimento. En estudios posteriores sería importante determinar si el efecto encontrado en el presente trabajo con respecto a disminuciones en la ingesta de comida en ratas al alargar el intervalo estímulo-comida también se encuentra con humanos.
La importancia del control de estímulos sobre la ingesta en humanos es reconocida cuando se diseñan programas de control de la ingesta excesiva en los que una parte fundamental del éxito de éstos consiste en eliminar cualquier estímulo que pueda estar relacionado con comida (Loro, Fisher & Levenkron, 1979; Stuart, 1967). Retomando el resultado del presente estudio con respecto a disminuciones en la ingesta en ratas en función de alargar el intervalo estímulo-comida, resultaría interesante determinar si esta misma manipulación aplicada a humanos también controla disminuciones en la cantidad de comida ingerida. Por ejemplo, podría diseñarse un programa en el que se incremente progresivamente el intervalo entre la presentación del estímulo que "anuncia" una comida (e.g., la vista u olor de un alimento no nutritivo) con su consumo y determinar si la separación temporal tiene efectos en la ingesta.
Es necesario señalar que los resultados del presente estudio apoyan la idea de que el inicio y fin de un periodo de alimentación no puede explicarse simplemente conforme a teorías homeostáticas y modelos de depleción/repleción de diversas substancias en el organismo, como glucosa y lípidos (cf. Weingarten, 1985). Esto es, se mostró que aun manteniendo a los sujetos a un nivel de privación constante fue posible controlar cambios en la cantidad de comida ingerida en función de presentar un estímulo exteroceptivo en diferentes ubicaciones temporales con respecto a la comida.
Finalmente, es importante mencionar que debido a que las personas se exponen diariamente a diferentes estímulos relacionados con la comida es posible que éstos controlen parte de la ingesta de comida diaria (Petrovich & Gallagher, 2007), por lo que el estudio de las condiciones que controlan aumentos y disminuciones en la cantidad de comida consumida en función de la presentación de estímulos es una tarea fundamental. A partir del estudio de la manera en que los estímulos controlan la conducta alimentaria se podrían sugerir algunas de las condiciones que controlan que las personas coman de manera excesiva lo que en general puede llevar a problemas de obesidad, o por el contrario, que coman de manera deficitaria y se llegue a problemas como la anorexia.
REFERENCIAS
Bruner, C. A. (2010). Conducta de comer: variables comunes a través del condicionamiento y la motivación. Revista Mexicana de Análisis de la Conducta, 36(2), 1-11. [ Links ]
Bruner, C. A., Gallardo, L., & Ávila, R. (2002). Variaciones en la ubicación temporal de una señal dentro de un ciclo de reforzamiento independiente de la respuesta sobre la actividad de las ratas. Revista Mexicana de Análisis de la Conducta, 28(1), 3-18. [ Links ]
Calvin, J. S., Bicknell, E. A., & Sperling, D. S. (1953). Effect of a secondary reinforcer on consummatory behavior. Journal of Comparative and Physiological Psychology, 46, 176-179. [ Links ]
Cornell, C., Rodin, J., & Weingarten, H. (1989). Stimulus-Induced eating when satiated. Physiology & Behavior, 45, 695-704. [ Links ]
De Castro, J. M., & Brewer, E. M. (1992). The amount eaten in meals by humans is a power function of the number of people present. Physiology and Behavior, 51, 121- 125. [ Links ]
Dews, P.B. (1966). The effect of multiple S delta periods on responding on a fixed-interval Schedule: IV. Effect of continuous S delta with Only short SD probes. Journal of the Experimental Analysis of Behavior, 9, 147-151. [ Links ]
Díaz, F., & Bruner, C. (2007). Comer y beber en ratas con libre acceso a la comida y al agua. Acta Comportamentalia, 15, 111-130. [ Links ]
Domjan, M. (2005). Pavlovian conditioning: A functional perspective. Annual Review of Psychology, 56, 179-206. [ Links ]
Farmer, J., & Schoenfeld, W. N. (1966a). Varying temporal placement of an added stimulus in a fixed-interval Schedule. Journal of the Experimental Analysis of Behavior, 9, 369-375. [ Links ]
Farmer, J., & Schoenfeld, W. N. (1966b). The effect of a response-contingent stimulus introduced into a fixed-interval Schedule at varying temporal placement. Psychonomic Science, 6, 15-16. [ Links ]
Keller, S., & Schoenfeld, N. (1950). Principles of psychology. Massachusetts, EE. UU.: Copley Publishing Group. [ Links ]
Kimble, G., & Ost, J. (1961). A conditioned inhibitory process in eyelid conditioning. Journal of Experimental Psychology, 61, 150-156. [ Links ]
Konorski, J. (1967). Integrative activity of the brain. An interdisciplinary approach. Chicago, EE.UU.: The university of Chicago press. [ Links ]
Loro, A. Fisher, E., & Levenkron, J. (1979). Comparison of established and innovative weight-reduction treatment procedures. Journal of Applied Behavior Anlaysis, 12, 141-155 [ Links ]
Lovibond, P. F. (1980). Effects of long- and variable-duration signals for food on activity, instrumental responding and eating. Learning and Motivation, 11, 164-184. [ Links ]
Pavlov, I. P. (1927). Conditioned reflexes. Londres: Oxford University Press. [ Links ]
Petrovich, G., & Gallagher, M. (2007). Control of food consumption by learned cues: A forebrain-hyphotalamic network. Physiology and Behavior, 91, 397-403. [ Links ]
Schachter, S., & Gross, P. (1968). Manipulated time and eating behavior. Journal of Personality and Social Psychology, 10(2), 98-106. [ Links ]
Siegal, P. S. (1961). Food intake in the rat in relation to the dark-light cycle. Journal of Comparative and Physiological Psychology, 54, 294-301. [ Links ]
Siegel, S., & Domjan, M. (1971). Backward conditioning as an inhibitory procedure. Learning and Motivation, 2, 1-11. [ Links ]
Skinner, B. F. (1932). Drive and reflex strength. The Journal of General Psychology, 6, 22-37. [ Links ]
Stuart, R. (1967). Behavioral control of overeating. Behavior Research Therapy, 5, 357-365. [ Links ]
Valle, F. (1968). Effect of exposure to feeding-related stimuli on food consumption in rats. Journal of Comparative and Physiological Psychology. 66(3), 773-776. [ Links ]
Weingarten, H. P. (1983). Conditioned cues elicit feeding in sated rats: A role for learning in meal initiation. Science, 220, 431-433. [ Links ]
Weingarten, H.P. (1985). Stimulus control of eating: Implications for a two-factor theory of hunger. Appetite, 6, 387-401. [ Links ]
Woods, S., & Seeley, R. (2002). Hunger and energy homeostasis. En R. Gallistel & H. Pashler (Eds.) Stevens Handbook of Experimental Psychology. (pp. 633 - 668). Hall Pushler. [ Links ]
Zamble, E. (1973). Augmentation of eating following a signal for feeding in rats. Learning and Motivation, 4, 138-147. [ Links ]
Recived: September, 21, 2011
Accepted: February, 7, 2012
1) Los datos de este artículo forman parte de la tesis de doctorado del primer autor cuyos estudios fueron apoyados por la beca 223216 del CONACYT. Dirigir correspondencia a cualquiera de los autores a: Laboratorio de Condicionamiento Operante, Facultad de Psicología, UNAM. Av. Universidad 3004, Cd. Universitaria, México, D.F. 04510. Correo electrónico: varsovia07@gmail.com o cbruner@servidor.unam.mx
