









Servicios Personalizados
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Compartir

 Permalink
PermalinkTemas em Psicologia
versión impresa ISSN 1413-389X
Temas psicol. vol.16 no.2 Ribeirão Preto 2008
ARTIGOS
Respostas de observação e respostas preparatórias em ratos
Preparatory responses and observing responses in rats
Gerson Yukio Tomanari
Universidade de São Paulo - Brasil
RESUMO
O presente estudo avaliou o papel de respostas preparatórias sobre contingências de respostas de observação. Seis ratos privados de água foram expostos a tentativas que eram encerradas com ou sem o acionamento do bebedouro. Respostas de observação podiam produzir estímulos discriminativos do tipo de tentativa corrente. Ao longo de diferentes condições experimentais, alternou-se o mecanismo de funcionamento do bebedouro de modo que o acesso ao mesmo podia ou não ser restrito temporalmente. Supostamente, a restrição no tempo de acesso à gota d'água aumentaria a probabilidade de ocorrência de respostas preparatórias, posto que permitiria maximizar o consumo de água. Os resultados mostraram que os efeitos dos diferentes mecanismos do bebedouro refletiram-se fundamentalmente na emissão de respostas na presença dos estímulos discriminativos, não na produção dos mesmos. De modo geral, observou-se um maior número de respostas na presença do estímulo discriminativo de ausência do que de presença de água, em particular na vigência do mecanismo restritivo temporalmente.
Palavras-chave: Respostas preparatórias, Respostas de observação, Reforço independente de resposta, Ratos.
ABSTRACT
The present study assessed the role of preparatory responses on observing-response contingencies. Six water-deprived rats were exposed to discrete trials that could end with or without water presentation. Observing responses produced discriminative stimuli of the current trial. Across experimental conditions, two dippers with different working mechanisms alternated. In one of them, but not in the other, access to water was temporally restricted. Supposedly, the temporal restriction would increase the occurrence of preparatory responses that led subjects to maximize water consumption. Results showed that dipper mechanisms affected the occurrence of responses in the presence of the discriminative stimuli, but not in their production. In general, there was a higher number of responses under the stimulus signaling absence of water, particularly when the time-restricted dipper was installed.
Keywords: Preparatory responses, Observing responses, Response-independent reinforcement, Rats.
Wyckoff (1969) demonstrou experimentalmente que, dada a oportunidade, animais emitem respostas que produzem estímulos discriminativos dos esquemas de reforçamento em vigor. Ele pretendia estudar como respostas que permitem a orientação do organismo a estímulos, as respostas de observação, são adquiridas ao longo do estabelecimento de uma discriminação (Wyckoff, 1952).
No procedimento pioneiro de Wyckoff (1969), pombos foram expostos a componentes nos quais alternavam-se esquemas em que ocorria reforçamento e esquemas em que não ocorria reforçamento. Um pedal próximo aos discos, quando acionado, produzia a apresentação dos estímulos relacionados ao componente em vigor. Os dados mostraram que, nessas condições, os pombos mantinham o pedal pressionado, o que fez Wyckoff denominar a resposta de pressionar o pedal de resposta de observação, posto que colocava o sujeito em contato com estímulos discriminativos.
Respostas de observação exercem um importante o papel nas situações em que é necessário que o sujeito comporte-se sob controle de estímulos. Vários procedimentos experimentais são planejados de forma que uma resposta é exigida apenas para garantir que o sujeito olhe para ou ouça um determinado estímulo (Dinsmoor, 1995a, 1995b; Sidman, 1994).
Uma característica importante em relação às respostas de observação é o fato de que elas não alteram a programação de reforços atribuída às respostas de reforçamento principal (água, comida, pontos etc.). A única conseqüência das respostas de observação é a produção de estímulos discriminativos. Em função disso, o procedimento de respostas de observação tem sido considerado a "melhor técnica disponível para demonstrar a legitimidade e a importância do próprio conceito de reforçamento secundário ou condicionado" (Dinsmoor, 1983, p. 696).
Por meio da análise das respostas de observação, dados têm apontado que o estímulo discriminativo de presença ou freqüência maior do reforçador primário (S+) adquire função reforçadora condicionada. Por outro lado, o estímulo discriminativo de ausência ou freqüência menor do reforçador primário (S-) não adquire função reforçadora condicionada ou, até mesmo, adquire função aversiva condicionada (Allen & Lattal, 1989; Case & Fantino, 1981, 1989; Case, Fantino, & Wixted, 1985; Case, Ploog, & Fantino, 1990; Dinsmoor, Bowe, Grenn, & Hanson, 1988; Dinsmoor, Browne, & Lawrence, 1972; Dinsmoor, Mulvaney, & Jwaideh, 1981; Fantino & Case 1983; Fantino, Case, & Altus, 1983; Jenkins & Boakes, 1973; Mueller & Dinsmoor, 1984, 1986; Preston, 1985; Tomanari, Machado, & Dube, 1998).
Os estudos em respostas de observação são unânimes em mostrar o valor reforçador de S+. No entanto, em relação ao papel exercido por S-, divergências são freqüentes. Em determinadas condições, o estímulo relacionado com ausência ou com menor probabilidade de reforçamento (S-) parece manter respostas de observação. Nesse caso, os dados indicariam a possibilidade de que S- exerça função reforçadora condicionada (Carvalho & Machado, 1992; Lieberman, 1972; Lieberman, Cathro, Nichol, & Watson, 1997; Perone & Baron, 1980; Roper & Baldwin, 2004; Shrier, Thompson, & Spector, 1980).
Com o objetivo de analisar o papel de S- na manutenção de respostas de observação, Tomanari e Machado (1996) submeteram ratos a tentativas que, em igual probabilidade, podiam ou não terminar com apresentação de reforço primário independentemente de qualquer resposta do sujeito. Respostas de observação emitidas em uma barra disponível no interior da caixa operante eram conseqüenciadas pela apresentação do estímulo discriminativo do tipo de tentativa em vigor. Salvo distinções em alguns parâmetros, esse mesmo procedimento foi também empregado por Blanchard (1975), Tomanari et al. (1998), Tomanari (2001) e Benvenuti e Tomanari (no prelo) com pombos, e Tomanari (2004), com humanos.
Os resultados obtidos por Tomanari e Machado (1996), diferentemente dos estudos com pombos e humanos, mostraram uma frágil manutenção de respostas de observação pelos estímulos discriminativos. Nas sessões iniciais, ocorreu um número alto de respostas, mas estas diminuíram gradualmente ao longo das sessões seguintes.
Para compreender os dados pouco robustos, Tomanari e Machado (1996) levantaram hipóteses acerca do papel das respostas de observação (e, portanto, da produção dos estímulos discriminativos) sobre respostas preparatórias que favoreceriam o consumo do reforço. No procedimento utilizado, o mecanismo de acionamento do bebedouro era tal que, em repouso, o bebedouro era normalmente mantido ao alcance dos ratos, no nível do chão da caixa operante. Ao ser acionado, mergulhava no tanque de água e retornava imediatamente à situação original. O bebedouro sempre disponível aos ratos, com praticamente nula restrição temporal para o consumo do reforço, pode ter estabelecido uma condição em que os estímulos discriminativos tornaram-se irrelevantes, pois não promoviam desempenhos diferenciais em sua presença. Por exemplo, mantendo-se o bebedouro sempre disponível, respostas de aproximação poderiam ocorrer a qualquer momento, uma vez que a gota d'água lá permaneceria até que fosse consumida. Ao contrário, caso houvesse uma restrição no tempo de acesso ao bebedouro, apenas uma aproximação imediata ao bebedouro logo após seu acionamento permitiria o consumo de água. Nesse caso, a produção dos estímulos discriminativos da tentativa ganharia relevância na medida em que controlaria as respostas preparatórias.
Considerando a possibilidade de que respostas de observação produzam e sejam mantidas pela produção de estímulos discriminativos que controlem respostas preparatórias, o presente experimento manipulou sistematicamente diferentes modos de atuação do mecanismo do bebedouro e analisou os efeitos correspondentes sobre as respostas de observação.
Método
Sujeitos
Foram utilizados 6 ratos albinos (Rattus novergicus), Wistar, machos, identificados como Ratos 19, 20, 21, 22, 5 e 6 com idade em torno de 120 dias no início do experimento. Os animais foram mantidos alojados em gaiolas individuais no biotério do Departamento de Psicologia Experimental da Universidade de São Paulo, local onde foram criados. Durante o experimento, os animais estiveram sob regime de restrição de acesso à água de modo que seus pesos fossem mantidos a cerca de 85% do valor ad libitum.
Equipamento
Foram utilizadas duas caixas de condicionamento operante modulares, modelo ENV-008, fabricadas por Med Associates Inc.. As caixas eram equipadas com um bebedouro (ENV-202M) localizado no centro da parede direita. A cada acionamento do bebedouro, uma concha trazia 0,06 ml de água do reservatório para o interior da caixa. À direita do bebedouro, havia uma barra de resposta (ENV-110M). Acima da barra, encontrava-se uma lâmpada tipo "olho de boi" (ENV-221M). Por meio de comandos de programação, esta lâmpada podia ser acionada de duas formas diferentes, isto é, acesa continuamente ou piscando em alternações de 0,5 s acesa e 0,5 s apagada. Localizada na parte centro-superior da parede oposta ao bebedouro, havia uma lâmpada de luz ambiente (ENV-215M). O piso da caixa era formado por barras metálicas eqüidistantes 1,6 cm entre si (ENV-005). As caixas estavam inseridas dentro de cubículos de isolamento acústico e visual. Um ventilador fazia a circulação de ar dentro do cubículo e emitia um ruído contínuo de fundo.
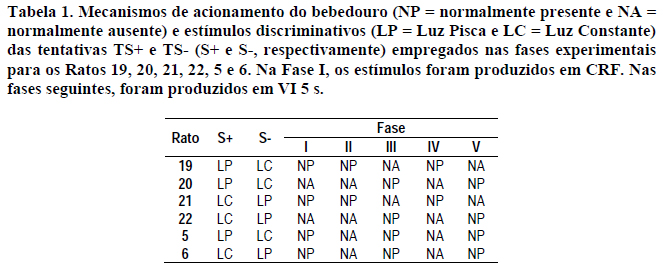
Características de acionamento do bebedouro: o bebedouro utilizado dispunha de dois mecanismos diferentes de acionamento, o Normalmente Ausente (NA) e o Normalmente Presente (NP). O mecanismo NA mantinha a concha d'água normalmente (i.e., em situação de repouso) mergulhada no reservatório localizado do lado externo da caixa. No momento em que era acionado, a concha levantava-se e colocava à disposição do sujeito, por 4 s, 0,06 ml de água. Terminado esse tempo, o bebedouro era desligado e retornava à posição de descanso, longe do alcance do animal. O mecanismo NP, na situação de repouso, mantinha a concha d'água normalmente presente no interior da caixa à disposição do rato. No momento em que era acionado, a concha mergulha no reservatório por 1 s e, a seguir, retornava ao interior da caixa, lá permanecendo até o próximo acionamento.
As sessões experimentais foram controladas por um micro-computador IBM-PC por meio de uma interface (DIG-700P1) e do aplicativo WMPCTM, que comandavam os eventos na caixa experimental (acionamento de estímulos, de mecanismos de bebedouro etc.) e registravam as respostas de pressão à barra. Os arquivos de controle e registro foram desenvolvidos especialmente para a execução desse experimento no programa Turbo Pascal v. 1.5, utilizando a linguagem MedState Notation.
Procedimento
Inicialmente, os seis sujeitos foram submetidos a uma sessão de modelagem da resposta de pressão à barra por aproximações sucessivas e, na seqüência, passaram por três sessões consecutivas de reforçamento contínuo por 200 reforços em cada uma delas. Os Ratos 19, 20, 21 e 22 foram modelados com o mecanismo do bebedouro NA, enquanto os Ratos 5 e 6 foram modelados com o mecanismo do bebedouro NP.
Os sujeitos foram submetidos a sessões diárias formadas por 32 tentativas discretas de 20 s de duração cada. Independentemente de qualquer resposta do sujeito, metade dessas tentativas terminava com a apresentação de 0,06 ml de água (tentativas TS+) e a outra metade terminava sem a apresentação de água (tentativas TS-). As tentativas foram apresentadas em seqüência aleatória determinada por sorteio (p=0,5), não tendo sido permitido que ocorressem mais do que três tentativas do mesmo tipo consecutivamente. Intercalados com as tentativas, havia intervalos entre tentativas (IET) de 10 segundos de duração em que nenhum estímulo era apresentado no interior da caixa experimental. Cada sessão teve início com um IET. O início de cada tentativa era sinalizado pelo acender da luz ambiente localizada na parede oposta ao bebedouro (estímulo discriminativo do esquema misto). Respostas de pressão à barra (respostas de observação), de acordo com o esquema em vigor (ver descrição abaixo), poderiam apagar a luz ambiente e produzir a apresentação de um estímulo discriminativo do tipo de tentativa em vigor, tentativa TS+ ou TS-. Um dos estímulos discriminativos consistiu em acender a luz acima da barra de forma contínua (luz constante). O outro consistiu em acender a luz acima da barra de forma piscante (luz pisca). Para metade dos sujeitos (Ratos 19, 20 e 5), a luz pisca sinalizava as tentativas TS+ e a luz constante sinalizava as tentativas TS-. Para os outros sujeitos (Ratos 21, 22 e 6), esta sinalização era invertida (Tabela 1). Uma vez apresentado um estímulo discriminativo, este permanecia na caixa até o final da tentativa.
No decorrer das fases experimentais, foram manipulados os mecanismos de funcionamento do bebedouro de forma que todos os sujeitos foram submetidos a ambos mecanismos: Normalmente Ausente (NA) e Normalmente Presente (NP), em diferentes seqüências (Tabela 1). Durante as condições em que vigorava o mecanismo NA, a concha d'água ficava disponível ao sujeito por 4 s contados do final da tentativa até o início do IET. Para igualar a duração das tentativas TS+ (com apresentação do bebedouro) e TS- (sem apresentação do bebedouro), assim como as condições de bebedouro NA e NP, este período de 4 s foi introduzido em ambos os tipos de tentativas (TS+ e TS-) e em ambas as condições de bebedouro (NA e NP). Nas tentativas em que havia produção de estímulo discriminativo, este permanecia presente na caixa durante os 4 s após o término da tentativa. Quando não ocorria produção do estímulo discriminativo, a luz ambiente apagava-se ao término da tentativa.
A apresentação dos estímulos discriminativos foi submetida a dois diferentes esquemas de reforçamento a depender da fase experimental. Na Fase I, foi empregado um esquema de reforçamento contínuo (CRF). Nas fases seguintes, foi usado um esquema de intervalo variável VI 5 s (intervalos 1, 1, 3, 4, 7, 14 s, conforme Fleshler e Hoffman, 1962). Durante uma tentativa, se não houvesse resposta de observação que cumprisse o critério do esquema em vigor, não ocorria a apresentação do estimulo discriminativo correspondente e o estímulo do esquema misto mantinha-se presente até o final da tentativa.
Resultados
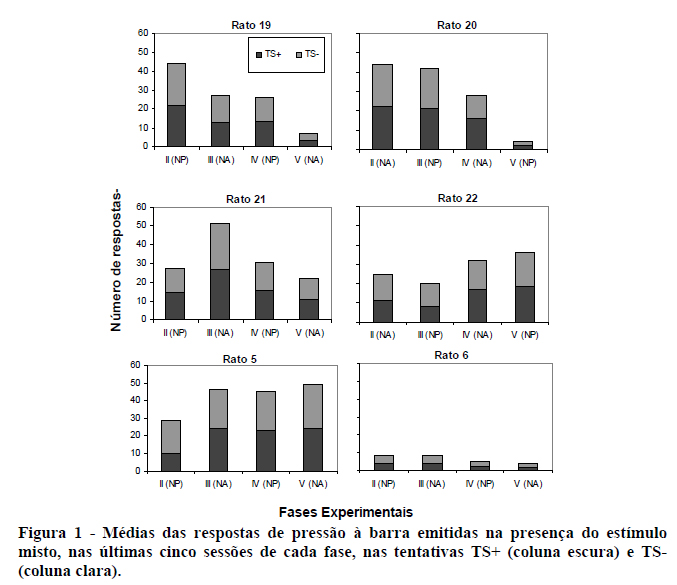
A Figura 1 mostra a freqüência de respostas de observação (respostas na presença do estímulo do esquema misto) como média das cinco últimas sessões de cada uma das fases realizadas no presente estudo, para cada um dos seis sujeitos, nas tentativas TS+ e TS-.
Nesta figura, em relação aos dados do Rato 19, pode-se observar uma diminuição no número de respostas de observação nas Fases III e V (mecanismo NA do bebedouro) em relação às fases respectivamente anteriores II e IV (mecanismo NP).
Em relação ao Rato 20, nota-se que o número de respostas ao final das Fases II (NA) e III (NP) permanece em torno de, em média, 43 respostas. Nas Fases seguintes, Fases IV (NA) e V (NP), há uma diminuição sistemática no número de respostas. Podemos notar que a primeira queda mais acentuada no número de respostas acontece na mudança do mecanismo de bebedouro de NP (vigente na Fase III) para NA (em vigor na Fase IV). A segunda ocorre na mudança de NA (Fase IV) para NP (Fase V).
Nos dados do Rato 21, pode-se observar um maior número de respostas de observação na Fase III (NA, em torno de 52 respostas) em relação às Fases II e IV (NP, em torno de 30 respostas). Para este sujeito, a segunda mudança do mecanismo do bebedouro de NP para NA (Fase V), diferentemente da primeira mudança, é acompanhada por uma diminuição no número de respostas de observação. As duas mudanças para NA (Fases III e V) são acompanhadas por alteração, ainda que em sentido contrário, na freqüência de respostas de observação. Entre as Fases II (NP) e III (NA), há aumento; entre as Fases IV (NP) e V (NA), há diminuição.
Os dados do Rato 22 mostram uma leve redução no número médio de respostas de observação da Fase II para a Fase III, seguido de um aumento gradual até a Fase V.
Para o Rato 5, há um aumento no número de respostas de observação quando ocorre a primeira mudança do mecanismo do bebedouro de NP para NA (Fases II e III). Uma leve queda acompanha o retorno do mecanismo para NP (Fase IV) e o número de respostas volta a aumentar na Fase V, com a segunda mudança do mecanismo para NA.
Os dados do Rato 6 mostram, de modo geral, um baixo número de respostas de observação (menos de dez respostas, em média, por sessão), em todas as quatro fases apresentadas. Ao longo do experimento, pode-se observar uma tendência de diminuição no número de respostas de observação independentemente do mecanismo de bebedouro em vigor, NA ou NP.
Comparando-se a proporção de respostas de observação nas tentativas TS+ e TS-, podemos observar que esta, para todos os sujeitos, em todas as fases, é semelhante. Este dado permite-nos concluir sobre o controle experimental de nossa programação. Conforme as contingências experimentais haviam sido programadas, a produção de ambos os estímulos, S+ e S-, encontrava-se em esquema VI 5 s. Tal semelhança de desempenho mostra, portanto, que o procedimento utilizado não contou com estímulos discriminativos adventícios; exceto pelos estímulos discriminativos S+ e S-, não havia, no procedimento, outra fonte de controle discriminativo correlacionada com o tipo de tentativa em vigor.
De modo geral, analisando-se o conjunto de dados mostrados na Figura 1, podemos constatar uma grande variabilidade nos dados entre sujeitos. Há instâncias em que o número de respostas de observação, quando vigora o mecanismo NA do bebedouro, é maior do que aquele registrado na fase imediatamente anterior em que vigorava o mecanismo NP. Este dado pode ser constatado em relação ao Rato 21, na Fase III; ao Rato 22, na Fase IV; e ao Rato 5, na Fase III. Ao contrário, há também instâncias, tais como aquelas reveladas pelos dados do Rato 19, que mostram que o número de respostas de observação, quando o mecanismo do bebedouro é NA, é menor do que o número de respostas emitidas na fase imediatamente anterior em que vigorava o mecanismo NP. Para ampliar a variabilidade, há ainda dados, tais como aqueles dos Ratos 20 e 6, que mostram a ausência de qualquer predominância quanto a um maior ou menor número de respostas de observação aparentemente relacionada com o mecanismo do bebedouro. Em função de tal variabilidade, parece correto afirmar que, nas condições propostas pelo presente estudo, os efeitos gerados pelas manipulações nos mecanismos do bebedouro sobre as respostas de observação não teriam sido preponderantes.
A Figura 2 mostra o tempo médio decorrido desde o início de cada tentativa até o momento em que os estímulos discriminativos foram produzidos. A figura é dividida em dois painéis que contêm os dados de todos os sujeitos, nas Fases II a V do presente experimento. Os dados dos Ratos 19, 20 e 21 encontram-se no Painel A, e os dados dos Ratos 22, 5 e 6 no Painel B. Em cada gráfico podemos encontrar, em destaque, as divisões das fases e a indicação do mecanismo do bebedouro utilizado em cada uma delas.
A Figura 2, por meio de uma análise mais detalhada de dados, mostra os possíveis efeitos dos diferentes mecanismos de bebedouro sobre o momento, na tentativa, em que os estímulos discriminativos foram produzidos. A hipótese levantada seria a de que, em vigor o mecanismo Normalmente ausente, as respostas de observação ocorreriam mais inicialmente nas tentativas caso estas interagissem com a emissão de respostas preparatórias no sentido de maximizar o tempo de contato com o bebedouro.
Para o Rato 19, a média do momento de produção dos estímulos se mantém em torno de 10 s nas Fases II, III e IV. Nas sessões finais da Fase IV, em vigor o mecanismo NP, há uma tendência de queda. A mudança do mecanismo para NA, na Fase V, é seguida por uma queda acentuada no tempo médio de produção dos estímulos, atingindo valores médios de 2 s, o que indica que estes estavam sendo produzidos próximo do início das tentativas.
O tempo médio de produção dos estímulos, para o Rato 20, na Fase II, com o mecanismo do bebedouro NA, mostra uma relativa variabilidade no início da fase. A partir da 12a sessão, há uma maior estabilidade em torno de 9 s. Essa estabilidade mantém-se por toda a Fase III, na qual foi introduzida a primeira mudança do mecanismo do bebedouro para NP. Quando o mecanismo de acionamento do bebedouro é alterado para NA (Fase IV), a variabilidade dos dados aumenta. Essa variabilidade permanece ao longo da Fase V, com o mecanismo do bebedouro NP. Durante essa fase, a partir da 10a sessão, os estímulos passam a ser produzidos mais inicialmente nas tentativas.
Os valores do tempo médio de produção dos estímulos discriminativos nas tentativas, para os Ratos 21, 22 e 5 mantêm-se praticamente constantes e estáveis por todas as fases do experimento. Para o Rato 21, a média do momento de produção dos estímulos situa-se em torno de 10 s. Os dados do Rato 22 apresentam essa média em torno de 11 s por todo o experimento.
Os dados do Rato 6 são os que apresentam a maior variabilidade no tempo de produção dos estímulos discriminativos no decorrer das sessões de todas as fases do experimento. Assim como foi observado nos dados dos outros sujeitos, não parece haver correlação entre o tipo de mecanismo do bebedouro utilizado e o momento de apresentação dos estímulos.
De um modo geral, observando a Figura 2, pode-se verificar que o tempo médio de produção dos estímulos discriminativos nas tentativas não apresenta variações sistemáticas correlacionadas com o mecanismo do bebedouro. Para todos os sujeitos, o tempo médio de produção dos estímulos na tentativa mantém-se praticamente constante por todo o experimento, exceto em alguns pontos isolados.
Respostas de pressão à barra emitidas na presença de S+ e S-, em cada uma das sessões, em todas as fases do experimento, para todos os seis sujeitos. A linha contínua refere-se às respostas emitidas na presença do S+ e a linha tracejada indica as respostas emitidas na presença do S-. Acima de cada gráfico, estão indicados o número da fase e o mecanismo do bebedouro utilizado durante a mesma, NA ou NP . A figura compõe-se por três painéis ( A, B e C) que se referem, respectivamente, aos sujeitos 19 e 20; 21 e 22; 5 e 6.
A Figura 3 mostra o número de respostas na presença dos estímulos discriminativos. Em relação aos dados do Rato 19, pode-se verificar que, em todas as sessões, em todas as fases apresentadas, há predominância de um maior número de respostas na presença de S- (em relação a S+). Nas sessões iniciais da Fase II (NP), esta diferença não ocorre; no entanto, a partir da 8a sessão, já pode ser observada. Em grande número de sessões das Fases III (NA) e IV (NP), o maior número de respostas em S- é bastante acentuado (por exemplo, na 2a sessão da Fase III, em que ocorrem 6 respostas em S+ e 18 em S-; e na 13a sessão da Fase IV, em que ocorrem 6 respostas em S+ e 17 em S-). Durante a Fase V, em muitas sessões, não ocorrem respostas na presença do S+. Nestas mesmas sessões, ocorrem respostas em S- (por exemplo, a 3a sessão, em que não ocorre resposta em S+, enquanto ocorrem 5 respostas em S-).
Os dados do Rato 20 revelam que a predominância de respostas na presença de S- se dá logo no inicio da Fase II (NA) e permanece em seu decorrer. Nas Fases III (NP), IV (NA) e V (NP), observa-se uma atenuação nesta predominância.
Quanto ao Rato 21, pode-se observar que, na Fase II (NP), a predominância de sessões em que ocorre maior número de respostas em S- também ocorre, porém é discreta. Na Fase III (NA), esta predominância torna-se evidente e a diferença claramente acentuada. Na Fase IV (NP), o desempenho é semelhante à Fase II (NP), ou seja, o número de respostas na presença do estímulo S- é bastante próximo ao número de respostas ocorridas na presença do S+. A diferença volta a aumentar quando o mecanismo do bebedouro muda pela segunda vez para NA, na Fase V. Novamente torna-se evidente o maior número de respostas emitidas na presença do S-.
Os dados do Rato 22 mostram, à semelhança dos outros sujeitos, a predominância, em todas as fases, de sessões em que ocorrem mais respostas em S-. Uma maior variabilidade no número de respostas pode ser observada nas Fases II e IV quando vigora o mecanismo NA.
Para o Rato 5, nas sessões da Fase II (NP), não se verifica qualquer predominância no número de respostas em S+ ou S-. Com a primeira inversão do mecanismo do bebedouro (de NP, Fase II, para NA, Fase III), percebe-se claramente que o número de respostas na presença do S- torna-se maior do que na presença de S+. Em todas as sessões dessa fase, o número de respostas durante a apresentação do S- foi superior à de S+. Este dado não se repete na fase seguinte (Fase IV), quando o mecanismo do bebedouro volta a ser NP. Na Fase V, no entanto, com a reinstalação do mecanismo NA, a predominância de respostas na presença de S- torna-se novamente evidente. Nesta fase, a diferença não é tão acentuada quanto na Fase III, mas é significativa e parece seguir uma tendência de aumento nas últimas duas sessões.
Os dados do Rato 6 mostram que, até a 10ª sessão da Fase II (NP), ocorre um número baixo de respostas (em torno de 1 resposta), tanto na presença do S+ quanto na de S-. A partir da 14a sessão, verifica-se uma predominância de respostas na presença do S-. Nas primeiras sessões da Fase III (NA), observam-se freqüências de respostas próximas de zero. A partir da 12ª sessão, há um aumento nas respostas emitidas na presença de ambos estímulos, porém predominantemente na presença do S-. Quando o mecanismo do bebedouro retorna a NP, na Fase IV, o número de respostas diminui rapidamente e permanece em torno de zero até o final, com exceção de um pico na 13ª sessão. Esse número continua próximo a zero também por toda a Fase V, cujo mecanismo do bebedouro utilizado foi NA.
Na Figura 3, observando-se o conjunto de dados de todos sujeitos, verifica-se que, de um modo geral, há predominância de sessões em que ocorre um número maior de respostas na presença de S- do que na presença de S+, sistematicamente para os seis sujeitos, exceto o Rato 5 na Fase II. Há sessões em que o número de respostas na presença de S+ é superior ao número de respostas na presença de S-, porém estas são ocorrências menos freqüentes.
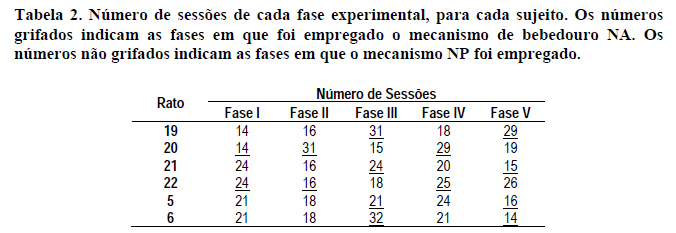
A Tabela 2 mostra o número de sessões transcorridas para cada rato em todas as cinco fases da presente investigação. Os números sublinhados indicam as fases em que foi empregado o mecanismo de bebedouro NA. Os números não sublinhados indicam as fases em que foi empregado o mecanismo NP.
Por meio da Tabela 2, pode-se observar que, de modo geral, quando vigora o mecanismo NA do bebedouro, o número de sessões na fase (23 sessões, em média) é maior do que quando vigora o mecanismo NP (20 sessões, em média). Para que uma fase fosse encerrada, era necessário que (1) se realizassem, no mínimo, quinze sessões, e que (2) houvesse estabilidade, verificada por inspeção visual, na freqüência de produção dos estímulos discriminativos em três sessões consecutivas. Dada a necessidade de estabilidade para o encerramento da fase, o número maior de sessões nas fases com mecanismo de bebedouro NA pode indicar que, nessas, a variabilidade do desempenho dos ratos era maior.
Discussão
O objetivo da presente investigação foi analisar o papel exercido pelas respostas preparatórias na manutenção das respostas de observação em ratos. Para isso, foram manipulados sistematicamente dois diferentes modos de atuação do mecanismo do bebedouro, um que permanecia constantemente disponível ao sujeito na caixa experimental, exceto quando era mergulhado na cuba d'água (Normalmente Presente, NP), e outro mecanismo em que o bebedouro ficava disponível ao sujeito somente por 4 s durante os períodos de reforçamento (Normalmente Ausente, NA). Supostamente, quando a disponibilidade de acesso ao reforço é restrita (mecanismo NA), estar-se-ia favorecendo a ocorrência de respostas de observação que produziriam estímulos discriminativos para as respostas preparatórias.
Em síntese, os dados obtidos no presente estudo mostram a manutenção de respostas de observação, em ratos, em um procedimento em que o reforçamento primário é independente de resposta. São poucos os estudos que analisam as respostas de observação em ratos (Preston, 1985; Carvalho & Machado, 1992; Roper & Baldwin, 2004; Tomanari & Machado, 1996). Entre esses, existem divergências quanto ao valor reforçador dos estímulos discriminativos na manutenção das respostas de observação em função, aparentemente, do fato da liberação do reforçador primário ser dependente ou independente de respostas. Preston (1985) e Carvalho e Machado (1992) relataram respostas de observação em esquema dependente de respostas. Em contrapartida, Tomanari e Machado (1996), empregando esquema independente de resposta, não chegaram a resultados semelhantes. A não demonstração de respostas de observação no estudo de Tomanari e Machado (1996) indicou a importância de se explorar as variáveis que afetam as respostas de observação em ratos como estratégia de se compreendê-las melhor. Entre outros fatores, características específicas da espécie, parâmetros de procedimento, a utilização de esquema independente de respostas, poderiam ter contribuído para os resultados obtidos por Tomanari e Machado (1996). Os resultados da presente investigação, nesse sentido, contribuem com uma descrição de um procedimento que permite estudar, em ratos, respostas de observação quando o reforçador primário é independente de respostas, e com a identificação de parâmetros de procedimento que poderiam ter gerado a ausência de respostas de observação no estudo de Tomanari e Machado (1996).
Quanto ao objetivo central da investigação, que foi o de verificar os efeitos de se empregar os mecanismos NA e NP do bebedouro sobre a emissão de respostas de observação, o que se observou nos dados foi a inexistência de sistematicidade inter e intra-sujeitos na freqüência de respostas de observação quando os mecanismos do bebedouro foram modificados. Tal variabilidade nos dados parece indicar que, nas condições propostas pelo presente estudo, os efeitos gerados pelas manipulações nos mecanismos do bebedouro sobre as respostas de observação não foram preponderantes. Portanto, os resultados do presente estudo não esclarecem a ausência de respostas de observação em Tomanari e Machado (1996) como uma decorrência da interação entre respostas de observação e respostas preparatórias.
Por outro lado, como uma forma de verificar se os efeitos dos diferentes mecanismos do bebedouro teriam atuação sobre uma variável dependente que não a freqüência de respostas de observação, o tempo médio de produção dos estímulos na tentativa foi analisado. A hipótese levantada seria a de que, em vigor o mecanismo Normalmente Ausente, as respostas de observação ocorreriam mais inicialmente nas tentativas caso estas interagissem com a emissão de respostas preparatórias, posto que poderiam possivelmente maximizar o tempo de contato do animal com o bebedouro. De um modo geral, os resultados desta análise não confirmam essa possibilidade, pois não foram verificadas regularidades entre o momento de produção dos estímulos discriminativos e o mecanismo de bebedouro em vigor. Para todos os sujeitos, o tempo médio de produção dos estímulos na tentativa manteve-se praticamente constante por todo o experimento, exceto por alguns pontos isolados.
Apesar da ausência de efeitos sistemáticos dos mecanismos de bebedouro sobre a emissão de respostas de observação, um dado bastante sistemático que se verificou na presente investigação refere-se ao número de respostas à barra emitidas na presença de S+ e de S- (esquema múltiplo). A este respeito, é predominante o número de sessões em que ocorre um maior número de respostas na presença de S-. Este dado parece correlacionar-se com os mecanismos do bebedouro na medida em que a discrepância entre o número de respostas em S+ e S- parece maior quando vigora o mecanismo NA.
Um outro dado também parece demonstrar pequenos efeitos produzidos pelos diferentes mecanismos do bebedouro. Trata-se do número de sessões realizadas em cada fase que, ao que os dados indicam, foi predominantemente maior quando esteve em vigor o mecanismo NA. Porque um dos critérios para o encerramento da fase era a estabilidade na freqüência de respostas de observação, este dado reflete a maior variabilidade produzida por este mecanismo do bebedouro em relação ao mecanismo NP.
Conclusivamente, portanto, futuras análises, manipulando-se as contingências envolvidas no contexto que produziram estes dados, poderão contribuir para a compreensão das interações mútuas entre respostas preparatórias e de observação, as quais configuram um sistema complexo de determinantes do comportamento.
Referências
Allen, K. D., & Lattal, K. A. (1989). On conditional reinforcing effects of negative discriminative stimuli. Journal of the Experimental Analysis of Behavior, 52, 335-339. [ Links ]
Benvenuti, M. F. L., & Tomanari, G. Y. (no prelo). Respostas de observação mantidas por estímulos compostos em pombos. Acta Comportamentalia. [ Links ]
Blanchard, R. (1975). The effect of S- on observing behavior. Learning and Motivation, 9, 1-10. [ Links ]
Carvalho, S. G., & Machado, L. M. C. M. (1992). Esquemas mistos e múltiplos concorrentes: uma reavaliação da resposta de observação. Acta Comportamentalia, 0, 109-144. [ Links ]
Case, D. A., & Fantino, E. (1981). The delay-reduction hypothesis of conditioned reinforcement and punishment: Observing behavior. Journal of the Experimental Analysis of Behavior, 35, 93-108 [ Links ]
Case, D. A., & Fantino, E. (1989). Instructions and reinforcement of adults and children. Learning and Motivation, 20, 373-412. [ Links ]
Case, D. A., Fantino, E., & Wixted, J. (1985). Human observing: maintained by negative informative stimuli only if correlated with improvement in response efficiency. Journal of the Experimental Analysis of Behavior, 43, 289-300. [ Links ]
Case, D. A., Ploog, B. O., & Fantino, E. (1990). Observing behavior in a computer game. Journal of the Experimental Analysis of Behavior, 54, 185-199. [ Links ]
Dinsmoor, J. A. (1983). Observing and conditioned reinforcement. Behavioral and Brain Sciences, 6, 693-728. [ Links ]
Dinsmoor J. A. (1995a). Stimulus control: Part I. The Behavior Analyst, 18, 51-68. [ Links ]
Dinsmoor J. A. (1995b). Stimulus control: Part II. The Behavior Analyst, 18, 253-269. [ Links ]
Dinsmoor, J. A., Bowe, C. A., Green, L., & Hanson, J. (1988). Information on response requirements compared with information on food density as a reinforcer of observing in pigeons. Journal of the Experimental Analysis of Behavior, 49. 229-237. [ Links ]
Dinsmoor, J. A., Browne, M. P., & Lawrence, C. E. (1972). A test of the negative discriminative stimulus as a reinforcer of observing. Journal of the Experimental Analysis of Behavior, 18, 79-85. [ Links ]
Dinsmoor, J. A., Mulvaney, D. E., & Jwaideh, A. R. (1981). Conditioned reinforcement as a function of duration of stimulus. Journal of the Experimental Analysis of Behavior, 36, 41-49. [ Links ]
Fantino, E., & Case, D. A. (1983). Human observing: maintained by stimuli correlated with reinforcement but not extinction. Journal of the Experimental Analysis of Behavior, 40, 193-210. [ Links ]
Fantino, E., Case, D. A., & Altus, D. (1983). Observing reward-informative and -uninformative stimuli by normal children of different ages. Journal of the Experimental Child Psychology, 36, 437-452. [ Links ]
Fleshler, M., & Hoffman, H. S. (1962). A progression for generating variable-interval schedules. Journal of the Experimental Analysis of Behavior, 5, 529-530. [ Links ]
Jenkins, H. M., & Boakes, R. A. (1973). Observing stimulus sources that signal food or no food. Journal of the Experimental Analysis of Behavior, 20, 197-207. [ Links ]
Lieberman, D. A. (1972). Secondary reinforcement and information as determinants of observing behavior in monkeys (Macaca mulatta). Learning and Motivation, 3, 341-358. [ Links ]
Lieberman, D. A., Cathro, S. S., Nichol, K., & Watson, E. (1997). The rule of S- in human observing behavior: Bad news is sometimes better than no news. Learning and Motivation, 28, 20-42. [ Links ]
Mueller, K. L., & Dinsmoor, J. A. (1984). Testing the reinforcement properties of S-: a replication of Lieberman's procedure. Journal of the Experimental Analysis of Behavior, 41, 17-25. [ Links ]
Mueller, K. L., & Dinsmoor, J. A. (1986). The effect of negative stimulus presentations on observing-response rates. Journal of the Experimental Analysis of Behavior, 46, 281-291. [ Links ]
Perone, M., & Baron, A. (1980). Reinforcement of human observing behavior by stimulus correlated with extinction or increased effort. Journal of the Experimental Analysis of Behavior, 34, 239-261. [ Links ]
Preston, G. C. (1985). Observing responses in rats: Support for the secondary reinforcement hypothesis. The Quarterly Journal of Experimental Psychology B: Comparative and Physiological Psychology, 37B, 23-31. [ Links ]
Roper, K. L., & Baldwin, E. R. (2004). The two-alternative observing response procedure in rats: Preference for non-discriminative stimuli and the effect of delay. Learning and Motivation, 35, 275-302. [ Links ]
Schrier, A. M., Thompson, C. R., & Spector (1980). Observing behavior in monkeys (Macaca arctoides): Support for the information hypothesis. Learning and Motivation, 11, 355-365. [ Links ]
Sidman, M. (1994). Equivalence relations and behavior: a research story. Authors Cooperative, Inc., Boston, MA. [ Links ]
Tomanari, G. Y. (no prelo). Resposta de observação: Uma reavaliação. Acta Comportamentalia. [ Links ]
Tomanari, G. Y. (2001). Respostas de observação controladas por estímulos sinalizadores de reforçamento e extinção. Acta Comportamentalia, 2, 119-143. [ Links ]
Tomanari, G. Y. (2004). Human observing behavior maintained by S+ and S-: Preliminary data. International Journal of Psychology and Psychological Therapy, 4, 155-204. [ Links ]
Tomanari, G. Y., & Machado, L. M. C. M. (1996). Observing responses in rats: effects of different schedules of reinforcement. Encontro Anual da Association for Behavior Analysis. Maio, São Francisco. [ Links ]
Tomanari, G. Y., Machado, L. M. C. M., & Dube, W. V. (1998). Pigeons' observing behavior and response-independent food presentations. Learning and Motivation, 29, 249-260. [ Links ]
Wyckoff, L. B. Jr. (1952). The role of observing responses in discrimination learning - Part I. Psychological Review, 59, 431-442. [ Links ]
Wyckoff, L. B. Jr. (1969). The role of observing responses in discrimination learning. In D. P. Hendry (Ed.). Conditioned Reinforcement. Homewood, IL: Dorsey Press. [ Links ]
 Endereço para correspondência:
Endereço para correspondência:
Gerson Yukio Tomanari
Av. Professor Mello Moraes, 1721
São Paulo, SP, CEP: 05508-030
E-mail: tomanari@usp.br
Enviado em Novembro de 2008
Revisado em Novembro de 2009
Aceite final Dezembro de 2009
Publicado em Maio de 2010
Esta pesquisa contou com a colaboração de José Luiz Balestrini Júnior, bolsista de iniciação científica da Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) entre 1999 e 2002. Agradecemos o apoio do CNPq do concedido na forma de auxílios à pesquisa e bolsa de Produtividade em Pesquisa.

{kind=link}
{kind=link}
{kind=link}
{kind=link}