










Services on Demand

article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkTemas em Psicologia
Print version ISSN 1413-389X
Temas psicol. vol.20 no.1 Ribeirão Preto June 2012
DOSSIÊ "ACERCA DA INTELIGÊNCIA HUMANA"
The interplay of behavioral dispositions and cognitive abilities: sociosexual orientation, emotional intelligence, executive functions and life history strategy
O interjogo entre disposições comportamentais e habilidades cognitivas: orientação sócio-sexual, inteligência emocional, funções executivas e estratégia de história de vida
La interacción de las disposiciones de comportamiento y las habilidades cognitivas: orientación socio-sexual, inteligencia emocional, funciones ejecutivas e historia de vida
Aurelio José FigueredoI; Alyssa Marie CuthbertsonI; Ilyssa Avery KauffmanI; Elana WeilI; Paul Robert GladdenII
IDepartment of Psychology, University of Arizona - Tucson, Arizona, USA
IIDepartment of Psychology and Sociology, Macon State College - Macon, Georgia, USA
ABSTRACT
Life History (LH) theory predicts that psychosocial traits will be selected to cluster into functionally coherent and coordinated suites of coadapted behavioral tactics. Among these coadapted tactics are behavioral dispositions, or "sociosexual orientations", towards either short-term mating (STM) and long-term mating (LTM), which are associated with fast and slow LH strategies, respectively. We predicted that certain specific mental abilities, executive functions (EF) and emotional intelligence (EI) should be higher in slow LH strategies, and should differentially predict STM and LTM, and partially mediate the relations between LH strategy and these different sociosexual orientations. 527 female undergraduates completed questionnaires measuring their LH strategies, EI, EF, and sociosexual orientations towards STM or LTM. The data were found to be consistent with our theoretically specified structural model. The results suggest that EF partially mediates both the relation between slow LH strategy and STM and the relation between slow LH and EI, and only indirectly influences LTM through its effects on STM and EI. In addition, EI partially mediates the relation between slow LH strategy and LTM. Implications for how LH strategy coordinates behavioral dispositions and the mental abilities required to implement them are discussed.
Keywords: Life history strategy, Long-term mating strategy, Sociosexual orientation, Executive functions, Emotional intelligence.
RESUMO
A teoria da História de Vida (Life History - LH) prevê que traços psicossociais serão selecionados de modo a se agruparem em conjuntos coordenados e coerentes de táticas comportamentais coadaptadas. Entre tais táticas coadaptadas estão disposições comportamentais, ou "orientações sócio-sexuais", dirigidas tanto a uniões de curto prazo (short-term mating - STM) e uniões de longo prazo (long-term mating - LTM), as quais são associadas a estratégias lentas e rápidas de LH, respectivamente. Predizemos que algumas habilidades mentais específicas, funções executivas (executive funcions - EF) e inteligência emocional (emotional intelligente - EI) devem ser altas em estratégias lentas de LH, e devem diferencialmente predizer STM e LTM, e parcialmente mediar as relações entre estratégias de LH e tais diferentes orientações sócio-sexuais. 572 estudantes de graduação do sexo feminino completaram questionários medindo suas estratégias de LH, EI, EF e orientações sócio-sexuais dirigidas a STM ou LTM. Os dados encontrados foram consistentes com nosso modelo estrutural teoricamente especificado. Os resultados sugerem que as EF parcialmente mediam ambas relações entre as estratégias LH lentas e STM e a relação entre LH lento e EI, e somente de maneira indireta influencia a LTM por meio de seus efeitos na STM e EI. Além do que, a EI parcialmente media a relação entre estratégia de LH lenta e LTM. Implicações para como a estratégia de LH coordena disposições comportamentais e habilidades mentais requeridas para implementá-las são discutidas.
Palavras-chave: Estratégia de história de vida, Estratégia de uniões de longo prazo, Orientação sócio-sexual, Funções executivas, Inteligência emocional.
RESUMEN
La teoría de Estrategia de Historia de Vida (EHV) predice que rasgos psicosociales serán seleccionados para agruparse en conjuntos funcionalmente coherentes y coordinados de tácticas conductuales coadaptadas. Entre estas tácticas coadaptadas se encuentran las disposiciones conductuales, u "orientaciones socio-sexuales", hacia el apareamiento a corto plazo (ACP) o el apareamiento a largo plazo (ALP), los cuales están asociados a las estrategias de LH rápidas y lentas, respectivamente. Nosotros predijimos que ciertas habilidades mentales específicas, las funciones ejecutivas (FE) y la inteligencia emocional (IE), deberían ser más elevadas en estrategias con EHV lenta, y deben predecir la ACP y ALP diferencialmente, y parcialmente mediar las relaciones entre la EHV y estas orientaciones socio-sexuales diferentes. 527 estudiantes femeninas completaron cuestionarios midiendo sus niveles de EHV, FE, IE, y orientaciones socio-sexuales hacia el ACP o el ALP. Los datos resultaron ser consistentes con nuestro modelo estructural y teóricamente especificado. Los resultados sugieren que la FE mediatiza parcialmente tanto la relación entre la EHV lenta y el ACP como la relación entre EHV lenta y la IE, y sólo influye el ALP indirectamente por medio de sus efectos sobre el ACP y la IE. Además, la IE mediatiza parcialmente la relación entre la EHV lenta y el ALP. Se abordan las implicaciones de cómo la EHV lenta coordina las disposiciones conductuales con las habilidades mentales necesarias para su ejecución.
Palabras clave: Estrategia de historia de vida, Estrategia de apareamiento a largo plazo, Orientación socio-sexual, Funciones ejecutivas, Inteligencia emocional.
Introduction
The purpose of this paper is to present the results of a structural model that follows the implications of Life History (LH) strategy for the development of certain specific mental abilities, Executive Functions (EF) and Emotional Intelligence (EI) through the role that these mental abilities are expected to have in the psychosocial and sociosexual development of systematic behavioral dispositions towards either Short-Term (ST) or Long-Term (LT) mating strategies.
Life History Strategy
LH strategy is a latent construct derived from Reproductive LH Theory (MacArthur & Wilson, 1967; Stearns, 1992; Roff, 1992, 2002). LH theory specifies the principles governing the allocation of bioenergetic and material resources among different components of fitness in all plants and animals. The reasoning is that all organisms must allocate some proportion of their total available resources over the life span to individual survival (somatic effort) and other portions to reproduction (reproductive effort). Although individuals may go through developmental stages in which all currently available resources are allocated exclusively to one or the other fitness component, it is ultimately necessary to invest in both somewhere along the line to avoid extinction of the lineage.
To illustrate, the majority of "hooknose" salmon spend most of their lives without engaging in any reproductive effort whatsoever, and concentrating instead on investing sufficient somatic effort on growth and maintenance to achieve the requisite size and strength to compete successfully for mates and breeding territories. Once they have achieved that first objective, they switch dramatically to expending all they have accumulated in a single but massive burst of reproduction, producing and fertilizing (depending on the sex of the individual) from two to ten thousand eggs at once (semelparity). As a result, post-reproductive individuals die shortly thereafter, having reserved nothing for future personal survival. Thus, although at each stage the salmon are investing exclusively in one component of fitness, over the lifespan they need to invest in each to promote the perpetuation of their lineage. Many other species follow less temporally polarized allocation strategies, and reserve some resources for current reproduction and other resources for continued survival as well as future reproduction (iteroparity).
Similarly, in sexually reproducing species, individuals must allocate some portions of their total reproductive effort to obtaining and retaining sexual partners (mating effort), and other portions of their total reproductive effort to producing and possibly rearing the resulting offspring (parental effort). Again, in some species the developmental switch from one pattern of allocation to the other may be extreme, but in other species (especially iteroparous ones), there is an alternation between the expenditure of mating and parental effort over time, and resources ultimately need to be reserved for both components over the life span as a whole.
LH strategies are generally considered to be arrayed along a single dimension, ranging from "fast" to "slow": (1) fast LH strategies allocate resources preferentially to reproductive over somatic effort, and to mating effort over parental effort; (2) slow LH strategies allocate resources preferentially to somatic over reproductive effort, and to parental effort over mating effort (Figueredo, Vásquez, Brumbach, & Schneider, 2007). In other words, fast LH strategists (such as rabbits) engage in heightened sexual activity and produce a large number of offspring, but invest little parental care in each individual offspring. Consequently, morbidity and mortality are high and most of the offspring produced are quite short-lived. In contrast, slow LH strategists (such as elephants) engage in more restricted levels of sexual activity and generally produce a smaller number of offspring, but invest a much larger proportion of parental care in each. Consequently, morbidity and mortality are reduced and many of the offspring produced may be quite long-lived.
Although, as with many other psychological constructs, such as the general factors of intelligence (g) and personality (GFP), there is always controversy over the adequacy of a single general factor in accounting for all the observed variation (and a certain degree of articulation of LH components has been reported, representing some more specific components of variance; see Ellis, Figueredo, Brumbach, and Schlomer, 2009), a single factor (K) has been successful over many replications in explaining a great deal of the variation that we observe in nature (Figueredo, Cabeza de Baca, & Woodley, 2012). This variation in LH strategy is observed both between species and within species, as both group differences and individual differences. As with all within-species group and individual differences, these variations have been shown to be jointly attributable to genetic and environmental influences (Figueredo, Vásquez, Brumbach, & Schneider, 2004; Figueredo & Rushton, 2009).
LH strategy thus has quite far-reaching implications for the development of both mental abilities and behavioral dispositions. We propose that humans require different sets of mental abilities to pursue either ST or LT mating strategies. Because ST and LT mating strategies are associated with faster and slower LH strategies, respectively, we propose that variation in LH strategy should predict the development of the requisite mental abilities to pursue these different sociosexual strategies, as consistent with the variant adaptive problems posed by each. Further, we propose that the development of these specific mental abilities should be differentially associated with the development of different sociosexual strategies, and thus serve as developmental mediators between LH strategy and behavioral dispositions towards either ST and LT mating.
Although the fundamental theoretical framework is developmental in nature, we tested a structural model of these hypothesized pathways on cross-sectional data, to determine whether the natural inter-individual variations observed in these constructs generally support the patterns predicted by theory. This procedure cannot, of course, conclusively demonstrate causation, but it can subject this causal theory to an initial test by subjecting it to risk of empirical falsification.
We start by working our way backwards from Sociosexual Orientation, to Emotional Intelligence, to Executive Functions and finally all the way back to LH strategy.
Sociosexual Orientation
The empirical measurement of mating strategies has long been linked to the development of the construct of sociosexual orientation, or sociosexuality for short. Research on sociosexuality began with the work of Alfred Kinsey, who, while studying population norms for sexual behavior in the late 1940s and early 1950s, noticed a covariance of many different sexual variables. These variables included the number of sexual partners, the onset of sexual behavior, the frequency of sexual intercourse, the incidence of concurrent affairs, and attitudes toward uncommitted sexual relationships (Kinsey, Pomeroy, & Martin, 1948; Kinsey, Pomeroy, Martin, & Gebhard, 1953; Simpson & Gangestad, 1991). Snyder, Simpson, and Gangestad (1986) and Gangestad and Simpson (1990) later suggested that these features may all be aspects of the same individual difference that was later termed sociosexual orientation or sociosexuality.
Sociosexuality is generally conceptualized as a single dimension measuring unrestricted sexual attitudes and behaviors. According to this conceptualization, those who score high on this dimension are unrestricted in their sociosexual orientation, meaning that they feel relatively comfortable engaging in sexual relationships that do not entail commitment or closeness. Conversely, those who score low on this dimension are restricted in their sociosexual orientation, meaning that they require commitment and emotional closeness from an individual before beginning a sexual relationship with that person.
The first externally validated construct of sociosexuality was developed in the early 1990s by Simpson and Gangestad (1991). The construct, known as the Sociosexual Orientation Inventory (SOI) is composed of different behavioral and attitudinal items that demonstrate sociosexuality's uniqueness from differences in sexual drive, sexual anxiety, sexual guilt, and sexual satisfaction (Simpson & Gangestad, 1991). Around the same time, the Sexual Strategies Theory (SST) of human mating was developed by Buss and Schmitt (1993); this is where the concept of mating strategies began. According to SST, the human selection of sexual mates is strategic, meaning that mate selection is based upon evolved solutions to adaptive problems with regards to reproducing as successfully as possible. It also contends that these mating strategies can be divided into two categories based upon the desired length of mating relationship, as adaptive mating problems differ with respect to the length of the mating relationship: (1) long-term mating strategy for long-term, committed relationships, and (2) short-term mating strategy for relationships of a shorter, less-committed or non-committed nature (Buss &Schmitt, 1993). As was quickly realized, the long-term mating strategy and the short-term mating strategy constructs of SST coincided with sociosexuality's restricted and unrestricted sociosexual orientations, respectively, and the SOI was subsequently used as an empirical measure of mating strategy.
However, it was soon discovered that the SOI is a less-than-ideal measure of mating strategy, primarily due to its one-dimensionality. The SOI treats the restricted sociosexual orientation and the unrestricted sociosexual orientation as opposite ends of a single continuum; conversely, long-term and short-term mating strategies represent separate constructs entirely. An individual can utilize both strategies at once with different mating partners, for instance, but an individual cannot be simultaneously restricted and unrestricted. Jackson and Kirkpatrick (2007) found that the SOI, in fact, contains three separate factors: Short-Term (ST) mating strategy, Long-Term (LT) mating strategy, and previous Sexual Behavior (SB).
In the present model, we use only the ST and LT mating strategy factors as our ultimate criterion variables, because the SB factor may depend as much upon mate value (the general attractiveness of individuals as social and sexual partners) as upon sociosexual orientation. Because LT mating is presumed to be an adaptation for long-term biparental care of offspring in humans (see Cabeza de Baca, Figueredo, & Ellis, in press), we hypothesize that it is preferentially associated with slow LH strategy. We therefore hypothesize that the specific mental abilities needed to develop and maintain LT sexual and romantic relationships should be preferentially associated with slow LH, and should therefore mediate this relation. Two major classes of mental ability that have been identified as serving this role are Emotional Intelligence and Executive Functions. We first by describing what is meant by Trait Emotional Intelligence.
Emotional Intelligence (EI)
Salovey and Mayer (1990) defined emotional intelligence as "the ability to monitor one's own and others' feelings and emotions, to discriminate among them, and to use this information to guide one's thinking and actions." Petrides (2010) considered this definition to be good conceptually, but sought to better operationalize the concept of emotional intelligence so that it could be scientifically tested. To do so, a distinction was created between the cognitive abilities required by emotional intelligence (as described above) and the wider domain of trait emotional intelligence (Trait EI), as more broadly described in Table 1. Trait EI facets are personality traits, and acknowledge the subjective nature of emotional experience. Furthermore, Trait EI is defined as a constellation of emotional self-perceptions located at the lower levels of personality hierarchies (Petrides, 2010). Higher Trait EI has been shown to be related to increased life satisfaction and coping, and decreased rumination, depression, dysfunctional attitudes, supporting the criterion and incremental validity of this construct. In addition, lower Trait EI can cause one to be susceptible to several mental abnormalities, including personality disorders such as paranoid, schizoid, schizotypal, borderline, dependent, and avoidant personality disorders (Petrides, Perez-Conzalez, & Furnham, 2007). Trait EI is not to be confused with general cognitive ability (g), as many studies have found zero correlations between measures of Trait EI and IQ tests (Mavroveli, Petrides, Sangareau, & Furnham, 2009).
Trait EI has been operationalized by means of multiple convergent indicators as consisting of the following domains (Petrides, 2010): (1) Adaptability, in which high scorers report being flexible and willing to adapt to new conditions; (2) Assertiveness, in which high scorers report being forthright, frank, and willing to stand up for their rights; (3) Emotional Expression, in which high scorers report being capable of communicating their feelings to others; (4) Emotion Management of the emotions of others, in which high scorers report being capable of influencing other people's feelings; (5) Emotional Perception of those of both self and others, in which high scorers report being clear about their own and other people's feelings; (6) Emotional Regulation, in which high scorers report being capable of controlling their emotions; (7) Low Impulsiveness, in which high scorers report being reflective and less likely to give in to their urges; (8) Relationships, in which high scorers report being capable of maintaining fulfilling personal relationships; (9) Self-Esteem, in which high scorers report being successful and self-confident; (10) Self-Motivation, in which high scorers report being driven and unlikely to give up in the face of adversity; (11) Social Awareness, in which high scorers report being accomplished networkers with superior social skills; (12) Stress Management, in which high scorers report being capable of withstanding pressure and regulating stress; (13) Trait Empathy, in which high scorers report being capable of taking someone else's perspective; (14) Trait Happiness, in which high scorers report being cheerful and satisfied with their lives; (14) Trait Optimism, in which high scorers report being confident and likely to "look on the bright side" of life.
Assessments of cognitive abilities and trait emotional intelligences (also known as emotional self-efficacies) differ not only in what they measure, but also in the methods use to measure them. Trait EI is measured via self-report questionnaires, whereas cognitive ability emotional intelligences are measured through tests of maximal performance. The most prominent measures of cognitive ability emotional intelligence are the Multifactor Emotional Intelligence Scale and the Mayer-Salovey-Caruso Emotional Intelligence Test. The most significant limitation of these measures, and others which attempt to asses cognitive ability emotional intelligence, is the subjectivity of emotional experience, making these tests very difficult to score. Prominent measures of Trait EI include the Bar-On Emotional Quotient Inventory and the Trait Emotional Intelligence Questionnaire (Perez, Petrides, & Furnham, 2005). The Bar-On Emotional Quotient Inventory was found to be slightly incomplete, as it does not include emotion expression, emotion regulation, and self-motivation, which are important parts of the domain of Trait EI. Other measures that cover the domain of Trait EI more extensively would be favorable (Petrides & Furnham, 2001).
The Trait Emotional Intelligence Questionnaire (TEIQue) is the premier operationalized measure of Trait EI. Petrides et al. (2010) discovered that the TEIQue overlaps with the Big Five dimensions of personality cross-culturally. The TEIQue has been validated cross-culturally and cross-developmentally, measuring Trait EI in both children and adults: (1) in one study, the TEIQue correlated positively with peer-related social competence and adaptive coping styles, but negatively to depression, somatic complaints, and maladaptive coping styles (Mavroveli, Petrides, Rieffe, & Bakker, 2007), and (2) in another study, the TEIQue correlated positively with emotion-perception and peer-rated social competence in children between the ages of 8 and 12 (Mavroveli et al., 2009). Such findings suggest that the validity of the TEIQue in measuring Trait EI can be generalized to diverse age and cultural groups.
Research has found that Trait EI, defined as the ability to monitor one's own and others feelings and emotions, to discriminate among them, and to use this information to guide one's thinking and action, is perceived, by the general public, as a primarily feminine attribute. Petrides, Furnham, and Martin (2004) had both male and female participants estimate there own and their parents' Trait EI. Estimates for one's parents were taken into account to determine whether sex differences are limited to the self or whether they generalize to include others. Data for self and parental estimates of Trait EI have shown that women estimated themselves to have more Trait EI than men, and both sexes rated their mothers as having more Trait EI than their fathers. Such findings provide evidence of yet another type of subjectivity when it comes to Trait EI. Because people tend to distinguish Trait EI as a feminine quality, men may rate themselves lower on constructs of Trait EI while women may rate themselves higher.
Trait EI has been found to be associated with various aspects of human experience. One such aspect is decision-related affect. Sevdalis, Petrides, and Harvey (2007) discovered that trait emotional intelligence was related to mood deterioration following the recall of a poor decision, over-predictions of post-decisional emotions, and a more positive emotional state five days after receiving the outcome of a poor decision. Such results show that Trait EI does not merely describe personality constructs. It is also capable of predicting behavior in various situations. Trait EI also predicts behaviors in the workplace. Petrides and Furnham (2006) found that high Trait EI was related to lower stress and higher levels of perceived control, satisfaction, and commitment to one's occupation; however, these relations may be indirect. For example, Trait EI may be linked to occupational commitment via perceived job control.
Studies have also been done to discover whether Trait EI domains are heritable, as are other personality traits. Vernon, Petrides, Bratko, and Schermer (2008) conducted two studies which sought to examine the heritability of Trait EI scores, as measured by the TEIQue. The first was a family study in which high-school students and their parents completed the TEIQue. It was found that about 1/3 of the Trait EI variance between individuals is attributable to genetic variation. It is necessary to keep in mind, however, that when using family data, the environmental influences must also be taken into account. The second study is a twin study, which is more informative than the first in terms of distinguishing genetic and environmental influences on Trait EI differences among individuals. Several adult monozygotic (MZ) and dizygotic (DZ) twins completed the TEIQue and a zygosity questionnaire that asked about the twins' physical similarity and the frequency with which they are mistaken for one another by other family members and friends. This study yielded similar results to the first, in that it concluded that differences in all of the dimensions measured by the TEIQue have shown degrees of genetic influence comparable in magnitude to the published heritabilities of other personality traits.
We therefore expect that Trait EI will be required primarily for the implementation of LT mating strategies, and does not contribute significantly to those of ST mating strategies, and thus will be preferentially developed by slow LH strategists. The relevant adaptive functions of Trait EI in this context are presumed to be in the development, maintenance, and nurturance of long-term pair bonds among sexual/romantic partners that are conducive to the provision of long-term biparental care of offspring (Cabeza de Baca, Figueredo, & Ellis, in press).
Executive Functions
Executive Functions (EF) are a set of cognitive processes that support future oriented, goal-directed behaviors (Morgan & Lilienfeld, 2000; Alvarez & Emory, 2006). These abilities include inhibiting prepotent responses, shifting from one mental task to another, and updating working memory (as with new goals or rules of a task; see Friedman et al., 2006). EF permits the cognitive flexibility required for planning, attention selection, decision-making, and behavioral regulation in everyday life; EF serves as the "conductor" that directs and coordinates the activities of lower-level cognitive and emotional systems (Miller & Cohen, 2001). Conversely, when EF is disrupted, the activity of the lower-level cognitive and emotional systems will predominate, leading to "impulsive" behaviors that may be inconsistent with future-oriented goals. This description is consistent with evidence suggesting that antisocial behaviors are increased among individuals scoring lower on neuropsychological tests of EF (for a review, see Morgan & Lilienfeld, 2000) because antisocial behaviors can also be seen as a class of impulsive behaviors (Gottfredson & Hirshi, 1990).
As mentioned above, one important EF is the ability to inhibit prepotent responses including impulsive and emotionally-laden ones in the service of future-oriented goals. For example, sexual desire or "lust" might be one such emotionally-laden response, but failing to inhibit these impulsive temptations is often strategically inconsistent with other goals (as when the potential mate is a friend's romantic partner, when one is attempting to maintain a committed relationship with someone else, or when the object of immediate desire is unwilling to copulate). Thus, EF might act to inhibit systems that produce desire for short-term sexual gratification. Similarly, another critical EF - the ability to shift attention from one mental set to another - might also disrupt the temptation for immediate sexual gratification by enabling attentional focus to shift away from the attractive object. In short, EF might serve to inhibit ST sexual behaviors where they are functionally inconsistent with LT mating strategies.
We therefore expect that higher levels of EF serve as a necessary prerequisite for the development of Trait EI, which we consider necessary for the implementation of LT mating strategies, but that EF will not contribute directly to the development or implementation of LT mating strategies, but indirectly through Trait EI, as a necessary set of cognitive abilities to implement and support emotionally intelligent behavior. Instead, aside from positively contributing to the development of Trait EI, EF should also function to inhibit the execution of impulsive ST sexual behaviors, which might otherwise produce strategic interference with LT mating strategies in slow LH strategists. In the service of these two adaptive functions, higher levels of EF should therefore be preferentially developed by slow LH strategists, as opposed to fast LH strategists, who hardly require EF for their ST sexual strategies.
We are not suggesting that the development of LT mating strategies represent the only adaptive functions served by EF in the context of a slow LH strategy, because long-term social relationships with same-sex peers as well as long-term sexual relationships can be facilitated by high levels of EF and Trait EI, as indicated by the occupational performance literature in industrial and organizational psychology briefly summarized above. However, we are predicting that the theoretically-specified effects of these two mental abilities should be clearly noticeable in the development of sociosexual orientations and have their evolutionary and developmental roots in different LH strategies.
We therefore offer a preliminary empirical test of these predictions.
Methods
Participants
Five-hundred and twenty-seven female undergraduate students, enrolled in an Introductory Psychology course participated. Participation was limited to female respondents because these data were originally collected in the context of a series of studies on Disordered Eating Behaviors (e.g., Salmon, Figueredo, & Woodburn, 2009). Of these 527 respondents, 94 cases were eventually excluded from the multivariate analyses due to excessive, non-recoverable missing data.
Procedure
Participants completed a series of self-report questionnaires on personal computers that assessed their Life History Strategies, Executive Functions, Trait Emotional Intelligence, and both Short-Term Mating and Long-Term Mating Sociosexual Orientations. Participants signed up for the study, provided informed consent, and completed the questionnaires over the internet through a secured research website maintained by the Ethology and Evolutionary Psychology Program of the University of Arizona.
Measures
The Arizona Life History Battery (ALHB; Figueredo, 2007; Gladden, Figueredo, & Jacobs, 2008) is a battery of cognitive and behavioral indicators of life history strategy compiled and adapted from various original sources. These self-report psychometric indicators measure graded individual differences along various complementary facets of a coherent and coordinated life history strategy, as specified by Life History Theory, and converge upon a single multivariate latent construct. They are scored directionally to indicate a "slow" (K-selected) life history strategy on the "fast-slow" (r-K) continuum. The Inter-Item Consistency Reliability (α) and Convergent Validity (λ) coefficients for these subscales on the general K factor were respectively as follows: α=.81 and λ=.83 for the Mini-K Short Form; α=.91 and λ=.63 for Insight, Planning, and Control; α=.93 and λ=.59 for Mother and Father Relationship Quality; α=.89 and λ=.68 for Family Social Contact and Support; α=.89 and λ=.58 for Friends Social Contact and Support; α=.92 for and λ=.46 Secure Romantic Partner Attachment; α=.91 for and λ=.63 General Altruism; and α=.96 and λ=.45 for Religiosity.
Behavioral Regulation Scales of the Behavior Rating Inventory of Executive Function - Adult version (BRIEF-A; Gioia, Isquith, Retzlaff, & Espy, 2002) was used to assess Executive Functions, including behavioral inhibition, cognitive inhibition, and appropriate regulation of behavior. This portion of the BRIEF-A is a 30-item self-report instrument of adult self-regulation in everyday environments that assess Emotional Control (e.g., "I overreact emotionally"), Inhibition (e.g., "I tap my fingers or bounce my legs"), Self-Monitoring (e.g., "I don't think about consequences before doing something"), and Set Shifting (e.g., "I have trouble changing from one activity or task to another"). The BRIEF-A scale used ranges from 0 (Never) to 6 (Almost Always) and asks participants to report the frequency of each problematic behavior over the past month. For present purposes, all items were reverse-scored to indicate better, rather than worse, Behavioral Regulation. The Inter-Item Consistency Reliability (α) and Convergent Validity (λ) coefficients for these subscales on the general EF factor were respectively as follows: α=.93 and λ=.82 for Emotional Control; α=.80 and λ=.82 for Inhibition; α=.86 and λ=.88 for Self-Monitoring; and α=.83 and λ=.87 for Set Shifting.
The Trait Emotional Intelligence Questionnaire (TEIQue; Petrides & Furnham, 2001; Petrides, 2009) is a self-report inventory that comprehensively covers the sampling domain of Trait EI, containing 153 items, measuring 15 distinct facets, which can be aggregated into 4 specific factors (Well-Being, Self-Control, Emotionality, and Sociability), and into a general factor score for global Trait EI. Sample items on the TEIQue include: "Understanding the needs and desires of others is not a problem for me" and "I always find ways to express my affection to others when I want to". The Inter-Item Consistency Reliability (α) and Convergent Validity (λ) coefficients for these subscales on the general Trait EI factor were respectively as follows: α=.68 and λ=.59 for the Adaptability; α=.74 and =.58 for Assertiveness; α=.76 and λ=.77 for Emotion Perception (self and others); α=.91 and λ=.69 for Emotion Expression; α=.71 and λ=.61 for Emotion Management (others); α=.73 and λ=.54 for Emotion Regulation; α=.76 and λ=.57 for Deliberation (low Impulsiveness); α=.78 and λ=.78 for Relationships; α=.86 and λ=.82 for the Self-Esteem; α=.74 and λ=.68 for Self-Motivation; α=.81 and λ=.82 for Social Awareness; α=.76 and λ=.69 for Stress Management; α=.75 and λ=.67 for Trait Empathy; α=.92 and λ=.83 for Trait Happiness; and α=.85 and λ=.84 for Trait Optimism.
The Multidimensional Sociosexual Orientation Inventory Short-Term Mating Scale (MSOI-STM; Jackson & Kirkpatrick, 2007) contains 10-items and measures personal preference for short-term mating (STM) patterns in their sexual/romantic relationships. The STM scale used ranges from -3 (Strongly Disagree) to +3 (Strongly Agree) and included items such as: "Sex without love is ok" and "Sometimes I'd rather have sex with someone I didn't care about." The Inter-Item Consistency Reliability (α) for the STM scale was α=.92; no general sociosexual orientation factor was constructed, so no Convergent Validity (λ) coefficient was available.
The Multidimensional Sociosexual Orientation Inventory Long-Term Mating Scale (MSOI-LTM; Jackson & Kirkpatrick, 2007) contains 10-items and measures preference for long-term mating (LTM) patterns in their sexual/romantic relationships. The LTM scale used also ranges from -3 (Strongly Disagree) to +3 (Strongly Agree) and includes items such as: "Finding a long-term romantic partner is not important to me" (reversed scored item) and "I can see myself settling down romantically with one special person." The Inter-Item Consistency Reliability (α) for the LTM scale was α=.86; no general sociosexual orientation factor was constructed, so no Convergent Validity (λ) coefficient was available.
Statistical Analyses
All univariate and multivariate analyses were performed using SAS 9.3. Because it was not feasible to analyze all of the individual subscales within a single multivariate model simultaneously without compromising our efficiency of parameter estimation and statistical power, a hierarchical analytical strategy was employed. Unit-weighted common factor scores (Gorsuch, 1983) were estimated, using SAS PROC STANDARD and DATA, as the means of the standardized scores for all non-missing subscales on each factor (Figueredo, McKnight, McKnight, & Sidani, 2000; McKnight, McKnight, Sidani, & Figueredo, 2007). The internal consistency reliability coefficients (Cronbach's alphas), the factor structure loadings (subscale-factor correlations) of the unit-weighted factors, and the covariance matrices of the subscales were computed using SAS PROC CORR.
All the unit-weighted factor scales were entered as manifest variables for multivariate causal analysis within a single structural equation model. Structural equation modeling (SEM) was performed by EQS. Structural equation modeling between these constructs then provided a multivariate causal analysis of the structural relations between them. SEMs were evaluated by use of Chi-Squared, the Bentler-Bonett Normed Fit Index (NFI), the Bentler-Bonnett Comparative Fit Index (CFI), and the Root Mean Squared Error of Approximation (RMSEA). Index values of the NFI and CFI greater than 0.90 are considered satisfactory levels of practical goodness-of-fit (Bentler & Bonnett, 1980; Bentler, 1995), whereas RMSEA values of 0.05 or less are considered indications of good fit, values between 0.08 and 0.10 are considered indications of a mediocre fit, and values greater than 0.10 are considered indications of a poor fit (Steiger & Lind, 1980; Browne & Cudeck, 1993). The CFI was selected because it is adjusted for model parsimony and performs well across a large range of sample sizes, especially with Maximum Likelihood estimation (Bentler, 1990; Hu & Bentler, 1995).
Results
Structural Equation Model
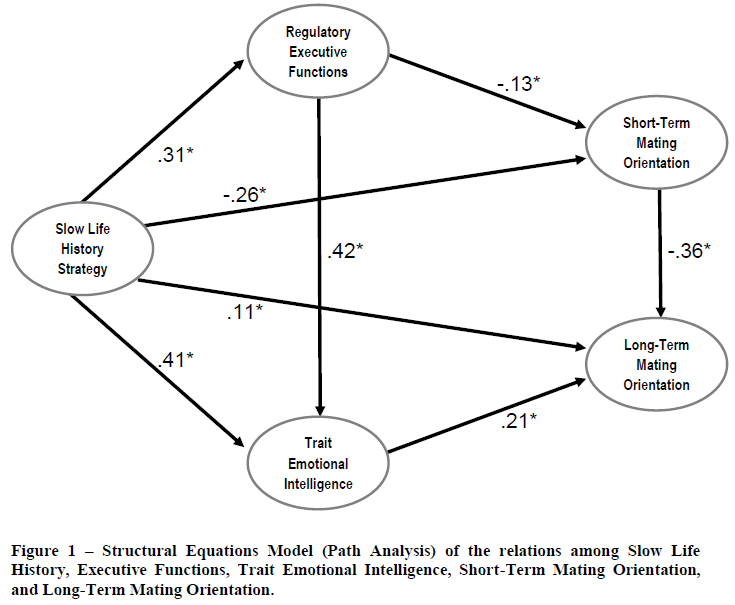
We hypothesized that the Slow Life History (SLH) factor would directly influence EF, EI, STM, and LTM, with the effect on STM being negative and all others positive in direction, based on the well-documented behavioral dispositions of slow LH strategists (see Figueredo, Cabeza de Baca, & Woodley, 2012). We further hypothesized that EF would directly influence, EI and STM, with the effect on EI being positive (facilitative) the effect on
STM being negative (inhibitory), but that EF would not directly influence LTM, based on the theoretical arguments detailed above. Finally, we hypothesized that EI would directly and positively influence LTM, but not directly influence STM, and that STM would directly and negatively influence LTM, due to the expectation of strategic interference among functionally inconsistent mating strategies.
The Structural Equations Model (SEM) specifying these hypothesized relations fit almost perfectly according to all statistical and practical indices of fit considered (2(2) = 3.911, p = .8224, NFI = .999, CFI = 1.000 , RMSEA = 0.000, with a 90% confidence interval ranging from .0000 to .0569). The results of these multivariate causal analyses are displayed in Figure 1. All path coefficients (effect sizes) tested were significantly different (p<.05) from zero, as are indicated by the asterisks (*). Standardized regression coefficients (β-weights) for the structural pathways are reported. All model parameters were estimated by Maximum Likelihood (ML).
The magnitudes and directions of the standardized path coefficients estimated by the SEM all very closely matched what was expected: (1) the SLH factor significantly predicted the EF factor (β = .31, p < 0.05), the EI factor (β = .41, p < .05), the STM scale (β = -0.26, p < .05), and the LTM scale (β = .11, p < .05); (2) the EF factor significantly predicted both the EI factor (β = .42, p < .05) and the STM scale (β = -.13, p < .05); (3) the EI factor significantly predicted the LTM scale (β = .21, p < .05); and (4) the STM scale also significantly predicted the LTM scale (β = -.36, p < .05).
Discussion
We have constructed and tested a path model that that follows the implications of Life History (LH) strategy for the development of Executive Functions (EF) and Emotional Intelligence (EI), through the roles that EF and EI are expected to have in the psychosocial and sociosexual development of systematic behavioral dispositions towards either Short-Term (ST) or Long-Term (LT) mating orientations. This path model has shown a very good fit to the data on a large number of female undergraduate students.
Although testing a structural model on cross-sectional data does not provide a conclusive test of causality, this evidence must nonetheless be seen as supportive of our theoretical position because the specific predictions we derived from LH theory have survived a critical risk of empirical falsification. Had our theory been grossly inadequate, the model would no doubt have been rejected by both statistical and practical criteria of goodness-of-fit in relation to the correlational data, especially given the relatively large effective sample size (N = 433) involved, which was aggregated across multiple studies (originally on Disordered Eating Behaviors) sharing these measures.
These and other convergent results lead us to consider whether a slow LH strategy entails a set of behavioral predispositions or a set of mental abilities or both (see Figueredo & Jacobs, 2010). For example, personality is generally deemed to be a set of behavioral predispositions, whereas intelligence is generally deemed to be a set of mental abilities. Could a slow LH entail both? An evolutionary scientist might reasonably ask how systematic behavioral predispositions could possibly evolve without the concurrent coevolutionary selection of the supporting mental abilities that are required to implement them.
We have argued here and elsewhere (e.g., Figueredo & Jacobs, 2010) that certain mental abilities, such as EF and EI, underpinning the behavioral self-regulatory control and the social competencies required for the maintenance of harmonious (mutualistic) social relations are essential to the implementation of a slow LH strategy in humans. Specifically, in spite of mostly verbal arguments to the contrary (Jackson & Kirkpatrick, 2007), we predict some degree of partial incompatibility between ST and LT mating strategies associated with fast and slow LH strategies, respectively, due to mutual strategic interference. This condition is reflected by the observation that although these two separate factors may be extracted from the MSOI by exploratory factor-analytic methods, they routinely show a significant correlation on the order of -.30, indicating a partially inverse relationship among them. The moderate magnitude of that relation does not preclude the occasional existence of the "mixed strategies" predicted by Jackson and Kirkpatrick (2007), but falls far short of the "independence" claimed between ST and LT mating strategies. In this view, a combination of a behavioral predispositions and a set of supporting mental abilities (associated with behavioral self-regulatory control and social competence or lack thereof) adaptively modulate the development and implementation of fast or slow LH strategies and their associated sociosexual orientations.
References
Alvarez, J. A., & Emory, E. (2006). Executive Function and the Frontal Lobes: A Meta-Analytic Review. Neuropsychological Review, 16,17-42. [ Links ]
Bentler, P. M. (1990). Fit indices, Lagrange Multipliers, constraint changes, and incomplete data in structural models. Multivariate Behavioral Research, 25,163-172. [ Links ]
Bentler, P. M. (1995). EQS structural equations program manual. Encino, CA: Multivariate Software. [ Links ]
Bentler, P. M., & Bonnett, D. G. (1980). Significance tests and goodness of fit in the analysis of covariance structures. Psychological Bulletin, 88,588-606. [ Links ]
Browne, M., & Cudeck, R. (1993). Alternative ways of assessing model fit. Sociological Methods and Research, 21,230-258. [ Links ]
Buss, D. M., & Schmitt, D. P. (1993). Sexual strategies theory: An evolutionary perspective on human mating. Psychological Review, 100,204-232. [ Links ]
Cabeza De Baca, T., Figueredo, A. J., & Ellis, B. J. (in press). An evolutionary analysis of variation in parental effort: Determinants and assessment. Parenting: Science and Practice. [ Links ]
Ellis, B. J., Figueredo, A. J., Brumbach, B. H., & Schlomer, G. L. (2009). Mechanisms of environmental risk: The impact of harsh versus unpredictable environments on the evolution and development of life history strategies. Human Nature, 20,204-268. [ Links ]
Figueredo, A. J. (2007). The Arizona Life History Battery. Retrieved from http://www.u.arizona.edu/~ajf/alhb.html [ Links ]
Figueredo, A. J., Cabeza de Baca, T., & Woodley, M. A. (in press). The measurement of human life history strategy. Personality and Individual Difference. [ Links ]
Figueredo, A. J., & Jacobs, W. J. (2010). Aggression, risk-taking, and alternative life history strategies: The behavioral ecology of social deviance. In M. Frias-Armenta & V. Corral-Verdugo (Eds.), Bio-Psycho-Social Perspectives on Interpersonal Violence (pp. 3-28). Hauppauge, NY: Nova Science Publishers. [ Links ]
Figueredo, A.J., McKnight, P.E., McKnight, K.M., & Sidani, S. (2000). Multivariate modeling of missing data within and across assessment waves. Addiction, 95(Supplement 3),S361-S380. [ Links ]
Figueredo, A. J., & Rushton, J. P. (2009). Evidence for shared genetic dominance between the general factor of personality, mental and physical health, and life history traits. Twin Research and Human Genetics, 12(6),555-563. [ Links ]
Figueredo, A. J., Vásquez, G., Brumbach, B. H., & Schneider, S. M. R. (2004). The heritability of life history strategy: The K-factor, covitality, and personality. Social Biology, 51,121-143. [ Links ]
Figueredo, A. J., Vásquez, G., Brumbach, B. H., & Schneider, S. M. R. (2007). The K-factor, covitality, and personality: A psychometric test of life history theory. Human Nature, 18(1),47-73. [ Links ]
Friedman, N. P., Miyake, A., Corley, R. P., Young, S. E., DeFries, J. C., & Hewitt, J. K. (2006). Not all executive functions are related to intelligence. Psychological Science, 17,172-179. [ Links ]
Gangestad, S. W., & Simpson, J. A. (1990). Toward an evolutionary history of female sociosexual variation. Journal of Personality, 58,69-96. [ Links ]
Gioia, G., Isquith, P., Retzkaff, P. D., & Espy, K. A. (2002). Confirmatory factor analysis of the Behavioral Rating Inventory of Executive Function [BRIEF] in a clinical sample. Child Neuropsychology, 8,294-257. [ Links ]
Gladden, P. R., Figueredo, A. J., & Jacobs, W. J. (2008). Life history strategy, psychopathic attitudes, personality, and general intelligence. Personality and Individual Differences, 46,270-275. [ Links ]
Gorsuch, R. L. (1983). Factor Analysis. Hillsdale, N.J.: Erlbaum. [ Links ]
Gottfredson, M. R., & Hirschi, T. (1990). A general theory of crime. Stanford, CA: Stanford University Press. [ Links ]
Hu, L. T., & Bentler, P. M. (1995). Evaluating model fit. In R. H. Hoyle (Ed.), Structural equation modeling: Concepts, issues, and applications (pp. 76-99). Thousand Oaks, CA: Sage. [ Links ]
Jackson, J. J., & Kirkpatrick, L. A. (2007). The structure and measurement of human mating strategies: toward a multidimensional model of sociosexuality. Evolution and Human Behavior, 28(6),382-391. [ Links ]
Kinsey, A., Pomeroy, W., & Martin, C. (1948). Sexual behavior in the human male. Philadelphia: Saunders. [ Links ]
Kinsey, A., Pomeroy, W., Martin, G., & Gebhard, P. (1953). Sexual behavior in the human female. Philadelphia: Saunders [ Links ]
MacArthur, R. H., & Wilson, E. O. (1967). The theory of island biogeography: Monographs in population biology. Princton, NJ: Princeton University Press. [ Links ]
McKnight, P. E., McKnight, K. M., Sidani, S., & Figueredo, A. J. (2007). Missing Data: A Gentle Introduction. New York, NY: Guilford. [ Links ]
Mavroveli, S., Petrides, K. V., Rieffe, C., & Bakker, F. (2007). Trait emotional intelligence, psychological well-being and peer-rated social competence in adolescence. British Journal of Developmental Psychology, 25,263-275. [ Links ]
Mavroveli, S., Petrides, K. V., Sangareau, Y., & Furnham, A. (2009). Exploring the relationships between trait emotional intelligence and objective socio-emotional outcomes in children. British Journal of Educational Psychology, 79,259-272. [ Links ]
Miller, E., & Cohen, J. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24,167-202. [ Links ]
Morgan, A. B. & Lilienfeld, S. O. (2000). A meta-analytic review of the relation between antisocial behavior and neuropsychological measures of executive function. Clinical Psychology Review, 20,113-136. [ Links ]
Perez, J. C., Petrides, K. V., & Furnham, A. (2005). Measuring trait emotional intelligence. Trait Emotional Intelligence, 123-143. [ Links ]
Petrides, K. V. (2009). Psychometric properties of the Trait Emotional Intelligence Questionnaire. In C. Stough, D. H. Saklofske, and J. D. Parker, Advances in the assessment of emotional intelligence. New York: Springer. [ Links ]
Petrides, K. V. (2010). Trait emotional intelligence theory. Industrial and Organizational Psychology, 2,136-139. [ Links ]
Petrides, K. V., & Furnham, A. (2001). Trait emotional intelligence: Psychometric investigation with reference to established trait taxonomies. European Journal of Personality, 15,425-448. [ Links ]
Petrides, K. V., & Furnham, A. (2006). The role of trait emotional intelligence in a gender- specific model of organizational variables. Journal of Applied Social Psychology, 36(2),552-569. [ Links ]
Petrides, K. V., Furnham, A., & Martin, G. N. (2004). Estimates of emotional and psychometric intelligence: Evidence for gender-based stereotypes. The Journal of Social Psychology, 144(2),149-162. [ Links ]
Petrides, K. V., Perez-Conzalez, J. C., & Furnham, A. (2007). On the criterion and incremental validity of trait emotional intelligence. Cognition and Emotion, 21(1),26-55). [ Links ]
Petrides, K. V., Vernon, P. A., Schermer, J. A., Ligthart, L., Boomsma, D. I., & Veselka, L. (2010). Relationships between trait emotional intelligence and the Big Five in the Netherlands. Personality and Individual differences, 48,906-910. [ Links ]
Roff, D. (1992). The evolution of life histories: Theory and analysis. New York: Chapman and Hall. [ Links ]
Roff, D. (2002). Life history evolution. Sunderland, MA: Sinauer Associates, Inc. [ Links ]
Salmon, C., Figueredo, A. J., & Woodburn, L. (2009). Life history strategy and disordered eating behavior. Evolutionary Psychology, 7(4),585-600. [ Links ]
Salovey, P., & Mayer, J. D. (1990). Emotional intelligence. Imagination, Cognition and Personality, 9,185-211. [ Links ]
Sevdalis, N., Petrides, K. V., & Harvey, N. (2007). Trait emotional intelligence and decision-related emotions. Personality and Individual Differences, 42,1347-1358. [ Links ]
Simpson, J. A., & Gangestad, S. W. (1991). Individual differences in sociosexuality: Evidence for convergent and discriminant validity. Journal of Personality and Social Psychology, 60(6),870-883. [ Links ]
Snyder, M., Simpson, J. A., & Gangestad, S. (1986). Personality and sexual relations. Journal of Personality and Social Psychology, 51,181-190. [ Links ]
Stearns, S. G. (1992). The evolution of life histories. Oxford, UK: Oxford University Press. [ Links ]
Steiger, J. H., & Lind, J. C. (1980, May). Statistically-based tests for the number of common factors. Paper presented at the annual Spring Meeting of the Psychometric Society in Iowa City. [ Links ]
Vernon, P. A., Petrides, K. V., Bratko, D., & Schermer, J. A. (2008). A behavioral genetic study of trait emotional intelligence. Emotion, 8(5),635-642. [ Links ]
 Address for correspondence:
Address for correspondence:
Aurelio José Figueredo
Director, Graduate Program in Ethology and Evolutionary
Psychology, Department of Psychology, 1503 East University Boulevard, School of Mind, Brain, and Behavior, College of Science, University of Arizona
Tucson, Arizona 85721-0068 USA; Voice: 520-621-7444
Fax: 520-621-9306
Email: ajf@u.arizona.edu
Web: http://www.u.arizona.edu/~ajf/
Received February 13th, 2012
Accepted March 31th, 2012
Published June 30th, 2012

{kind=link}