










Services on Demand

article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkTemas em Psicologia
Print version ISSN 1413-389X
Temas psicol. vol.21 no.1 Ribeirão Preto June 2013
https://doi.org/10.9788/TP2013.1-04
ARTIGOS
A percepção do tempo: contributos do procedimento de bissecção
Time perception: contributions of the bisection procedure
La percepción del tiempo: contribuciones del procedimiento de bisección
Ana Catarina Vieira de Castro; Marilia Pinheiro de Carvalho Andréia; Kroger-Costa Armando Machado
Escola de Psicologia da Universidade do Minho, Braga, Portugal
RESUMO
Desde o início do século XX que a psicologia se tem interessado pelo estudo experimental da regulação temporal do comportamento, mas esta área de pesquisa, conhecida como timing, só se consolidou a partir da década de 1960. Desde então, vários procedimentos utilizados com sujeitos de diferentes espécies têm permitido identificar as características mais gerais da regulação temporal aprendida. Um destes procedimentos, talvez o mais amplamente utilizado, é o procedimento de bissecção temporal. Este procedimento tem permitido formalizar algumas das propriedades chave da discriminação temporal de estímulos e possibilitado o teste de diferentes modelos teóricos de timing. No presente trabalho, apresentamos uma revisão da bissecção temporal, justificamos a sua centralidade no estudo do timing, descrevemos as variações no procedimento de base, apresentamos os principais resultados obtidos com ele, e identificamos algumas aplicações recentes a participantes humanos.
Palavras-chave: Bissecção temporal, discriminação temporal, função psicométrica, ponto de bissecção, timing.
ABSTRACT
Since the beginning of the twentieth century psychology has been interested in the experimental investigation of the temporal regulation of behavior but this area of research, known as timing, was consolidated only in the decade of 1960. Since then, several procedures used with subjects of different species have allowed the identification of the most general characteristics of learned temporal regulation. One of those procedures, perhaps the most widely used, is the temporal bisection procedure. This procedure has allowed the formalization of some key properties of the temporal discrimination of stimuli and has enabled the test of distinct theoretical models of timing. In the present work we make a brief review of the temporal bisection, justify its centrality in the study of timing, describe the variations in the basic procedure, present the main results obtained with it and identify some recent applications to human participants.
Keywords: Psychometric function, point of bisection, temporal bisection, temporal discrimination, timing.
RESUMEN
Desde el comienzo del siglo XX, la psicología se ha interesado por la investigación experimental de la regulación temporal de la conducta, pero esta área de investigación, conocida como timing, sólo se consolidó en la década de 1960. Desde entonces, varios procedimientos usados con sujetos de diferentes especies han permitido la identificación de las características más generales de la regulación temporal aprendida. Uno de esos procedimientos, quizás lomás utilizado, es el procedimiento de bisección temporal. Este procedimiento ha permitido la formalización de algunas propiedades fundamentales de la discriminación temporal de los estímulos y ha permitido poner a la prueba distintos modelos teóricos de timing. En el presente trabajo hacemos una breve revisión de la bisección temporal, justificamos su centralidad en el estudio del timing, describimos las variaciones en el procedimiento básico, presentamos los principales resultados obtenidos con ele e identificamos algunas aplicaciones recientes con participantes humanos.
Palabras clave: Bisección temporal, discriminación temporal, función psicométrica, punto de bisección, timing.
Para compreender o comportamento de qualquer organismo é necessário relacionar esse comportamento com o ambiente em que ele ocorre. Poderíamos até dizer, sem qualquer exagero, que no estudo do comportamento nada faz sentido senão à luz do ambiente. Ora, o ambiente pode ser concebido como um conjunto complexo de objetos que se distinguem uns dos outros pelos seus vários atributos (forma, tamanho, cor, movimento, temperatura, número, etc.). A um nível mais analítico, o ambiente é composto pelos estímulos-luzes e sons, cheiros e sabores, por exemplo - aos quais o animal é sensível. Uma parte substancial da pesquisa psicológica consiste pois em caracterizar que sensibilidade um determinado animal tem às propriedades dos estímulos que o rodeiam.
Uma das propriedades elementares de qualquer estímulo é a sua duração. Este fato, em si trivial, era caro aos pioneiros da psicologia experimental pois explicava porque razão todas as sensações, os elementos da vida mental, possuíam o atributo da duração. Para Titchener (1915), por exemplo, o atributo duração era
o acontecer puro, o avançar, mantendo-se igual a si próprio, que pode ser observado em toda e qualquer sensação; reconhece-mo-lo mais facilmente, talvez, quando ouvimos um som ou atendemos ao complexo cinestésico produzido quando lentamente estendemos o braço para além do cotovelo. É o fator tempo elementar presente em todas as nossas percepções do tempo [grifos do autor], - nas percepções de um período, de um intervalo, de uma taxa, de um ritmo, e assim por diante. (pp. 122-123)
Não surpreende, por isso, que o estudo da percepção temporal tenha as suas origens nos primórdios da psicologia experimental.
Por razões inerentes ao método introspetivo, esses estudos iniciais (ver Boring, 1957) estavam forçosamente limitados ao sujeito humano. No entanto, sendo a duração um atributo fundamental do ambiente, seria de esperar que muitos outros animais fossem sensíveis a ela (Gallistel, 1993; Grondin, 2010; Michon, 1993; Richelle & Lejeune, 1980; Roberts, 1998; Shettleworth, 1998). De fato, a partir da segunda metade do século XX, vários estudos revelaram que, tal como os seres humanos, muitas outras espécies animais são sensíveis à duração dos estímulos ambientais, podendo inclusive aprender a regular o seu comportamento com base nesta propriedade. Estes estudos definiram uma nova área de pesquisa, conhecida por timing em inglês,e que estuda os processos e mecanismos de regulação temporal com animais humanos e não-humanos.
A regulação temporal do comportamento divide-se em duas grandes classes, a classe associada aos ritmos biológicos, que explora as periodicidades estáveis do ambiente, como a alternância do dia e da noite (ritmos circadianos), das marés, ou das estações do ano, por exemplo, e a classe associada a intervalos de tempo significativamente mais curtos e de duração arbitrária, da ordem de segundos a minutos. As correspondentes áreas de estudo designam-se, respectivamente, por timing circadiano e timing intervalar (Church, 2002; Richelle & Lejeune, 1980; Roberts, 1998; Shettleworth, 1998). É sobre a segunda área que nos debruçamos no presente artigo. E como a pesquisa com animais não-humanos é menos conhecida do que a pesquisa com humanos, não só em termos de técnicas como de resultados, teorias, e modelos, concentraremos a nossa atenção nos estudos com animais não-humanos. Como veremos, estes estudos têm contribuído para elucidar os processos de regulação temporal não só em não-humanos como também em humanos.
Como referimos, o estudo do timing intervalar em animais não-humanos só se tornou mais sistemático e reconhecido como um domínio próprio de pesquisa a partir da segunda metade do século XX. O grande entrave para um início mais precoce residiu sobretudo em questões técnicas e metodológicas. Do ponto de vista técnico, foi necessário esperar por equipamentos que automatizassem os procedimentos experimentais e, assim, permitissem efetuar sessões diárias prolongadas, frequentemente de uma ou mais horas de duração, e durante vários meses, o tempo necessário para observar o fenômeno de interesse. Foi também preciso esperar pela tecnologia que permitisse manipular e medir com rigor a duração dos estímulos. Sem um módico de automatização de procedimentos e medidas, os estudos simplesmente não eram possíveis. Do ponto de vista metodológico, foi preciso esperar pelo desenvolvimento de procedimentos de treino e de teste que permitissem isolar de algum modo o fenômeno de interesse, a regulação do comportamento em função de um ou mais intervalos de tempo.
Apesar das limitações tecnológicas e metodológicas, as primeiras evidências de regulação temporal do comportamento em animais não-humanos datam do início do século XX. Utilizando procedimentos de condicionamento clássico, Pavlov (1927/1960) e os seus estudantes demonstraram a sensibilidade de cães à duração de intervalos de tempo arbitrários. Num procedimento de condicionamento clássico, um estímulo relativamente arbitrário (p. ex., o som de um metrônomo) é apresentado várias vezes em proximidade temporal com um estímulo biologicamente relevante (p. ex., um pedaço de carne para um cão com fome). A comida é designada por estímulo incondicional porque provoca uma resposta reflexa inata de salivação, designada por resposta incondicional; o som é designado por estímulo neutro porque inicialmente não provoca a resposta de salivação. Após alguns emparelhamentos som/comida, o som passa a provocar uma resposta de salivação, a resposta condicional, semelhante à resposta incondicional produzida pela comida. O som, inicialmente neutro, passou a ser um estímulo condicional capaz de produzir uma resposta de salivação.
Variações no procedimento básico de condicionamento clássico, nomeadamente no grau de contiguidade entre os estímulos condicional e incondicional, revelaram que as varáveis temporais influenciavam fortemente o comportamento dos animais. A tal ponto o tempo era importante, que poderia mesmo substituir o som como estímulo condicional. Assim, a apresentação de comida a intervalos fixos de tempo, digamos a cada 30 minutos, dava origem, após algumas tentativas de treino, a uma resposta condicional de salivação que ocorria próximo do final do intervalo de 30 minutos. Ou seja, neste procedimento de condicionamento temporal, a resposta condicional não se distribuía indiferenciadamente ao longo do intervalo, mas tendia a concentrar-se nos momentos que antecediam a entrega de comida. Para além disso, quando a comida era removida, a resposta de salivação continuava a ocorrer aproximadamente com a mesma periodicidade, de 30 em 30 minutos. Pavlov concluiu que o tempo, ou eventos periódicos, não podiam ser negligenciados no estudo do comportamento e que, pelo contrário, deviam ser reconhecidos como variáveis a ser exploradas.
Ao mesmo tempo que Pavlov desenvolvia a sua pesquisa sobre o condicionamento clássico, alguns estudos isolados sobre percepção temporal, utilizando procedimentos instrumentais, iam também sendo desenvolvidos em laboratórios americanos. Um procedimento instrumental envolve uma situação na qual um sujeito tem de emitir um comportamento ou ação específicos para obter uma recompensa. Anderson (1932) e Sams e Tolman (1925), por exemplo, utilizaram um procedimento no qual ratos colocados num labirinto escolhiam entre diferentes corredores, sendo depois retidos nos corredores por diferentes períodos de tempo. A escolha do corredor com menor intervalo de retenção revelaria percepção do tempo. Em outros experimentos, os sujeitos eram confinados por diferentes períodos de tempo numa caixa de partida e, posteriormente, quando libertados, tinham de escolher um dos braços do labirinto em função da duração passada na caixa de partida (p. ex., Cowles & Finan, 1941; Heron, 1949; Yagi, 1962). Por exemplo, se o rato tivesse ficado confinado na caixa de partida durante 5 s devia escolher o braço esquerdo do labirinto para ter acesso à comida; se o confinamento durasse 25 s o rato devia escolher o braço direito para ter acesso à comida.
Os sujeitos tiveram dificuldade em aprender a tarefa nos estudos com labirintos e o desempenho, de um modo geral, era muito irregular. A razão para esta dificuldade estava relacionada com as características dos procedimentos utilizados. Estes requeriam a presença contínua do experimentador e as tentativas eram demasiado curtas para obter uma boa regulação temporal do comportamento. Para além disso, em estudos com labirintos e corredores, variáveis espaciais e temporais podiam confundir-se, na medida em que os sujeitos podiam guiar-se por pistas espaciais para solucionar uma tarefa que os experimentadores programaram com base nas condições temporais.
Uma avaliação de percepção temporal menos contaminada por variáveis intervenientes foi conseguida num estudo de Woodrow (1928) com macacos rhesus. A duração a ser avaliada era o intervalo que decorria entre dois sons. Se o intervalo durasse 4,5 s, era disponibilizada comida numa caixa; se a duração do intervalo fosse de 1,5 s, o macaco não encontrava comida na caixa. Os dois macacos utilizados no experimento aprenderam a abrir a caixa só depois da duração mais longa. Este procedimento, no entanto, exigia ainda muito tempo por parte do experimentador, uma vez que requeria milhares de tentativas - situação que, não sendo incomum neste tipo de treino, torna a automatização dos procedimentos altamente desejável.
Paralelamente ao desenvolvimento tecnológico, o desenvolvimento de metodologia experimental apropriada para pesquisar a capacidade dos animais estimarem o tempo foi central para o fomento da pesquisa em timing. A grande contribuição metodológica surgiu com os procedimentos operantes avançados por Ferster e Skinner (1957) e por Skinner (1938), nos quais eram utilizados programas de reforço baseados em intervalos de tempo.
Num destes programas de reforço, o programa de intervalo fixo (FI, do inglês, fixed interval), um animal recebe um pedaço de comida pela primeira resposta que emitir após a passagem de um intervalo de tempo contado a partir da entrega do pedaço de comida anterior. Imagine, assim, que uma bolinha de comida é dada a um rato e, logo de seguida, é iniciado um cronômetro. No final de 20 s, o rato recebe mais uma bolinha de comida logo que pressione uma barra. Este programa de reforço designa-se por FI20s. Observe que o rato pode pressionar a barra durante todo o intervalo, mas só a resposta emitida após o fim do intervalo de 20 s é reforçada com comida. Passadas algumas sessões de treino, verifica-se que o rato se comporta de maneira diferente no decurso do intervalo de 20 s. No início do intervalo, quando a disponibilidade de comida está ainda distante, o rato não responde ou responde muito raramente, mas a partir de cerca da metade ou dois terços do intervalo, o rato passa a responder a uma velocidade ou taxa mais elevada. Em média, a taxa a que o animal responde aumenta à medida que se aproxima o momento em que a comida fica disponível, sendo máxima nos últimos instantes do FI. O desempenho dos sujeitos em programas de reforço de intervalo fixo é uma evidência forte da sensibilidade do comportamento à passagem do tempo.
Apesar dos programas de reforço incluírem o tempo como variável crítica no estudo do comportamento, o trabalho intensivo de Ferster e Skinner (1957) e Skinner (1938) não tinha como foco principal a percepção temporal. O seu interesse residia sobretudo nas taxas de resposta produzidas por diferentes programas de reforço e nos fatores que afetavam essas taxas de resposta. Só mais tarde na história da psicologia, o programa de intervalo fixo e suas variantes começaram a ser utilizados para o estudo direto de questões relativas à percepção temporal (ver Lejeune, Richelle, & Wearden, 2006).
Em 1968 é publicado aquele que é considerado o trabalho inaugural no estudo do timing intervalar, assim considerado por ter tratado da regulação temporal do comportamento de forma central e independente da discussão dos programas de reforço. Stubbs (1968) conduziu uma série de três estudos nos quais pesquisou a psicofísica da percepção temporal em pombos. O procedimento consistia em ensinar os sujeitos a bicar uma tecla vermelha após a apresentação de qualquer um dos estímulos pertencentes ao conjunto Curto (i.e., estímulos com durações de 1, 2, 3, 4 ou 5 s), e a bicar uma tecla verde após a apresentação de qualquer um dos estímulos pertencentes ao conjunto Longo (i.e., estímulos com durações de 6, 7, 8, 9 e 10 s). A escolha da tecla vermelha ou da tecla verde indicava a percepção que os sujeitos tinham de uma dada duração.
Stubbs (1968) analisou os resultados em termos de percentagem de respostas na tecla verde (i.e., percentagem de respostas 'longo') em função de cada uma das durações treinadas. A partir do treino, seria de se esperar que os sujeitos não respondessem à tecla verde após qualquer um dos estímulos do conjunto Curto (0% de respostas 'longo'), e que respondessem sempre à tecla verde após a apresentação de qualquer um dos estímulos do conjunto Longo (100% de respostas 'longo'). No entanto, os resultados revelaram que o comportamento dos sujeitos não era categórico; a percentagem de respostas 'longo' aumentou gradualmente de 0 a 100% com o aumento da duração do estímulo apresentado. As durações mais curtas (1 s e 2 s) produziram muito poucas (cerca de 0%) respostas 'longo', as durações mais longas (9 s e 10 s) produziram muitas (cerca de 100%) respostas 'longo', e as durações intermediárias produziram percentagens intermediárias de respostas 'longo'. Os resultados revelaram também que as durações mais próximas do limite que distinguia os dois conjuntos (5 s e 6 s) produziram cerca de 50% de respostas 'longo'. O mesmo padrão de resultados obtido com o leque de durações de 1 s a 10 s (Stubbs, 1968, Experimento 1) foi obtido com leques de durações que variavam entre 1 s e 40 s (Experimento 2).
A partir do trabalho de Stubbs (1968), a pesquisa na área do timing intervalar recebeu cada vez mais atenção dos pesquisadores e é hoje uma das áreas de pesquisa mais prolíficas da psicologia experimental. Os procedimentos operantes utilizados nesta área são divididos em duas categorias principais: regulação temporal e discriminação temporal. Nos procedimentos de regulação temporal, a sensibilidade temporal do comportamento é revelada pelo modo como as respostas dos sujeitos se distribuem ao longo de um intervalo de tempo. O procedimento de intervalo fixo (FI) é um exemplo de um procedimento de regulação temporal. Nos procedimentos de discriminação temporal, a sensibilidade ao tempo é revelada pela aprendizagem de diferentes respostas em função de durações de tempo distintas. O procedimento desenvolvido por Stubbs (1968) é um exemplo de um procedimento de discriminação temporal.
Um dos procedimentos de discriminação temporal mais utilizado nos últimos 30 anos, tanto com não-humanos quanto com humanos, é o procedimento de bissecção temporal, usado por Catania (1970) com macacos rhesus e desenvolvido com mais profundidade por Church e Deluty (1977) com ratos. Este último estudo é, hoje em dia, considerado um clássico da literatura porque formalizou algumas das propriedades psicofísicas fundamentais do timing, propriedades essas apenas esboçadas nos trabalhos anteriores (p. ex., Stubbs, 1968). Inúmeros estudos efetuados com o procedimento de bissecção temporal têm produzido resultados semelhantes com diferentes espécies animais, permitindo assim enunciar algumas das propriedades mais gerais da percepção temporal.
No presente artigo resumimos a história recente da bissecção temporal. Mais concretamente, descrevemos as características da tarefa, incluindo as variações usadas com diferentes espécies. Em seguida, descrevemos as técnicas mais comuns de análise de dados e os principais resultados (sob a forma de regularidades psicofísicas) que essas técnicas evidenciaram. Damos ainda exemplos da aplicação do procedimento de bissecção temporal a situações socialmente relevantes. Por fim, resumimos dois modelos teó-ricos que procuram integrar os resultados empíricos encontrados até o momento e explicar os processos subjacentes à discriminação temporal.
O Procedimento de Bissecção
Imagine que pretendemos saber se pombos são capazes de discriminar entre dois intervalos de tempo distintos. Para tal, construímos uma tarefa na qual os pombos têm de aprender a emitir uma resposta se um estímulo previamente apresentado for de curta duração (1 s, por exemplo) e a emitir uma outra resposta se o mesmo estímulo for de longa duração (4 s, por exemplo). A Figura 1 descreve a ordem dos acontecimentos neste procedimento, quando este é realizado numa caixa de condicionamento operante para pombos. Inicialmente, a tecla central (i.e., um disco de plástico translúcido com cerca de 2,5 cm de diâmetro) é iluminada por trás com luz branca durante 1 s ou 4 s (I). A iluminação desta tecla designa-se por estímulo modelo. Quando o intervalo de tempo de 1 s ou 4 s acaba, a luz da tecla central é apagada e as duas teclas laterais são iluminadas, uma com luz vermelha e a outra com luz verde (II). As teclas laterais designam-se por estímulos de comparação. Se a duração da iluminação da tecla central tiver sido de 1 s, a resposta correta é bicar na tecla vermelha; se tiver sido de 4 s, a resposta correta é bicar na tecla verde. Ou seja, a duração do estímulo modelo indica que estímulo comparação é correto. Após uma resposta correta, as teclas laterais apagam-se e é ativado o dispensador de comida durante um breve intervalo de tempo, geralmente de 3 s. Após uma resposta incorreta, todas as luzes da caixa se apagam durante um outro intervalo de tempo, de cerca de 3 a 5 s, mas o dispensador de comida não é ativado (III). Cada sequência de eventos, desde a apresentação do estímulo modelo na tecla central até à recompensa ou punição (timeout, assinalado com um 'X' na Figura 1), que se segue à resposta do pombo nas teclas laterais, constitui uma tentativa experimental. Um intervalo de cerca de 20 s separa tentativas sucessivas (ITI, do inglês, inter-trial interval). Uma sessão diária pode incluir 100 tentativas, das quais, por exemplo, 50 seriam com o estímulo modelos de 1 s e 50 com o estímulo modelo de 4 s; em cada um destes grupos de 50 tentativas, 25 teriam a tecla vermelha à esquerda e a tecla verde à direita, e os outros 25 teriam a disposição contrária.
Após uma série de sessões experimentais, os pombos aprendem a responder 'vermelho' após durações de 1 s e a responder 'verde' após durações de 4 s. Designamos as duas durações de treino por Curta e Longa e as respectivas respostas corretas por 'curta' e 'longa'.
A grande contribuição deste procedimento para o estudo do timing está nas possibilidades que ele oferece a partir do momento em que uma discriminação temporal é aprendida. Imagine, por exemplo, que depois de o pombo aprender a discriminação entre 1 s e 4 s acima descrita, realizamos um teste, no qual apresentamos estímulos modelo de duração intermediária às durações treino (p. ex., 1,5; 2,0; 2,5; 3,0 e 3,5 s). O que fará o pombo nestes testes? Será que, apesar de não ter sido exposto a estas durações, o pombo generalizará o que aprendeu, respondendo'curto' após durações próximas de 1 s e 'longo' após durações próximas de 4 s? Neste caso, qual será a duração que o pombo classificará o mesmo número de vezes como 'curto' e 'longo' (i.e., 50% de respostas em cada estímulo comparação). Por outras palavras, que duração será julgada equidistante das duas durações de treino e, por isso, bissecta o contínuo temporal entre 1 s e 4 s?1 E que implicações decorrem do valor do ponto de bissecção quanto à natureza da escala subjetiva do tempo?
Outras questões importantes dizem respeito à generalidade dos resultados. Será que o desempenho se mantém quando variamos os valores absolutos das duas durações de treino, o número, a amplitude e o espaçamento das durações usadas no teste, a modalidade sensorial do estímulo modelo ou a topografia da resposta perante os estímulos de comparação? E será que os resultados são os mesmos para diferentes espécies animais, incluindo seres humanos? Como veremos a seguir, ao procurar responder a estas perguntas, os investigadores descobriram ou confirmaram algumas das propriedades mais importantes do timing.
Caracterização do Procedimento
Sujeitos
O procedimento de bissecção temporal já foi utilizado com várias espécies animais, incluindo estorninhos (Hulse & Kline, 1993), pombos (Fetterman & Killeen, 1991), ratos (Church & Deluty, 1977) e macacos (Catania, 1970). No entanto, as espécies mais comumente utilizadas são pombos e ratos. Com participantes humanos o procedimento já foi usado com crianças (Droit-Volet & Wearden, 2001), adultos (Wearden & Ferrara, 1995) e idosos (Droit-Volet & Wearden, 2001; McCormack, Brown, Maylor, Darby, & Grenn, 1999). Outros estudos examinaram a percepção temporal em populações clínicas, como doentes de Alzheimer (Caselli, Iaboli, & Nichelli, 2009), Parkinson (Merchant, Luciana, Hooper, Ma-jestic, & Tuite, 2008) e esquizofrenia (Carroll, Boggs, O'Donnell, Shekhar, & Hetrick, 2008).
O número de sujeitos experimentais varia de estudo para estudo, tanto na investigação não-humana quanto na investigação humana, e é geralmente definido com base nos requisitos das condições experimentais planejadas e dos testes estatísticos a usar na análise dos dados. Nos casos em que há perda de sujeitos (p. ex., por desistência, doença, morte, ou dificuldade de aprendizagem da tarefa, por exemplo), há que se considerar a necessidade de introduzir sujeitos adicionais aos inicialmente planejados.
A pesquisa com sujeitos não-humanos costuma ser mais prolongada do que com participantes humanos. Um número relativamente elevado de sessões diárias de treino (entre 1520 sessões, em média) é necessário até que os animais aprendam a tarefa, ou seja, até que respondam corretamente às duas durações de treino em mais de 85% das tentativas. Os estudos com humanos são, usualmente, muito mais breves, não sendo raro, por exemplo, que os dados sejam todos coletados numa única sessão de 1 hora. Como o número de sujeitos tende a ser muito maior nos estudos com humanos do que nos estudos com não-humanos, obtém-se assim um equilíbrio interessante entre os dois tipos de estudos: O que os estudos com humanos ganham em número de sujeitos, perdem em tempo de estudo de cada sujeito.
Materiais
Os materiais utilizados variam de acordo com os sujeitos experimentais. No caso de sujeitos não-humanos, a coleta de dados é geralmente realizada em caixas operantes adaptadas para a espécie em estudo. As caixas para ratos (Figura 2, painel da esquerda) contêm lâmpadas e fontes sonoras que podem ser ligadas por diferentes períodos de tempo para representarem os estímulos temporais, e barras que funcionam como dispositivos de resposta. Estas barras podem ser fixas ou retráteis e, no segundo caso, geralmente só são apresentadas ao sujeito no momento da emissão da resposta. As caixas para pombos (Figura 2, painel da direita) contêm teclas de plástico translúcido, que têm duas funções. Primeiro, como podem ser iluminadas por trás com luzes de diferentes cores e intensidades, servem para apresentar os estímulos. Por outro lado, como têm acoplado por trás um microswitch, servem ainda para receber e registrar a resposta de bicar. Adicionalmente, ambas as caixas incluem um dispositivo dispensador de comida ou água, utilizado para o reforço das respostas corretas.
Com participantes humanos, a apresentação dos estímulos ocorre geralmente na tela de um computador, quando se trabalha com estímulos visuais (que são, usualmente, formas geométricas coloridas), e alto-falantes ou fones de ouvido, quando são utilizados estímulos sonoros (geralmente sons com frequências entre 500 e 1000Hz). As respostas são dadas utilizando o mouse, o teclado de computador ou equipamento adaptado (por exemplo, um joystick ligado ao computador). Tanto em experimentos com participantes humanos como com sujeitos não-humanos, o controle dos acontecimentos da sessão experimental e o registro dos dados é efetuado por um computador programado para o efeito.
Procedimento: Descrição Técnica
Os procedimentos de bissecção temporal utilizados com animais não se alteraram significativamente desde o estudo de Church e Deluty (1977). Neste estudo, numa caixa operante para ratos, uma lâmpada de iluminação geral acendia-se durante 30 s, o ITI. No final deste período, a luz era apagada durante 1 s ou 4 s (os autores utilizaram diferentes pares de durações para além de 1 s e 4 s - p. ex., 2 s e 8 s, 3 s e 12 s, e 4 s e 16 s, mas para maior clareza descreveremos apenas o treino da discriminação entre 1 s e 4 s). A duração do período de escuridão na caixa representava o estímulo modelo. Decorrido este intervalo de escuridão, as duas barras eram introduzidas na caixa, uma à esquerda e outra à direita. Nas tentativas em que as barras eram introduzidas após 1 s de escuridão, a resposta correta consistia em pressionar a barra da esquerda. Nas tentativas em que as barras eram introduzidas após 4 s de escuridão, a resposta correta era pressionar a barra da direita. A discriminação temporal aprendida pelos ratos pode ser esquematizada da seguinte forma: '1 s→Esquerda, 4 s→Direita'.
Respostas corretas eram reforçadas com uma bolinha de comida e, em seguida, iniciava-se um novo ITI e, posteriormente, uma nova tentativa começava. Respostas incorretas eram seguidas imediatamente pelo ITI e, depois, pela repetição da tentativa, procedimento que se designa por procedimento de correção2. Numa sessão experimental, em metade das tentativas era apresentada a duração de 1 s e, na outra metade, era apresentada a duração de 4 s. As sessões experimentais decorriam diariamente, durante 15 dias.
Posteriormente, Church e Deluty (1977) realizaram uma fase de teste. Cada sessão de teste incluía ¼ de tentativas de treino e ¾ de tentativas de teste. As tentativas de treino eram iguais às tentativas da fase anterior e foram incluídas durante a fase de teste para garantir que os sujeitos mantinham a discriminação inicial. Nas tentativas de teste foram apresentadas durações novas entre 1 s e 4 s (concretamente, 1,2; 1,6; 2,0; 2,5 e 3,1 s). Após o estímulo modelo, o sujeito continuava a ter que escolher entre as barras da esquerda e da direita, mas as suas respostas nestas tentativas não eram reforçadas. A fase de teste envolveu dez sessões experimentais diárias.
Sistematizando, uma tarefa de bissecção temporal com sujeitos não-humanos envolve uma fase de treino e uma fase de teste. Durante a fase de treino, em cada tentativa, é apresentada uma de duas durações modelo. Após a apresentação do modelo, o sujeiro tem de emitir uma de duas respostas; uma delas é correta após a apresentação de uma das durações e a outra é correta após a apresentação da outra duração. Respostas corretas são seguidas pela entrega de um reforço. Quando ocorre uma resposta incorreta, para além de não haver reforço, pode ser utilizado um procedimento de correção. Neste caso, a tentativa é repetida até a resposta correta ser emitida (Church & Deluty, 1977), ou até se atingir um número limite de repetições (p. ex., três); depois de atingido esse limite, apenas a resposta correta é disponibilizada (p. ex., Machado & Arantes, 2006). Após uma resposta incorreta pode também ser introduzido um timeout, que adia o início de uma nova tentativa (p. ex., Stubbs, 1968). Durante a fase de teste são apresentadas durações intermediárias às durações treinadas e as mesmas opções de resposta da fase de treino estão disponíveis. Geralmente, as respostas nas tentativas de teste não são seguidas pela entrega de reforço.
Nos estudos com participantes humanos, o procedimento sofre algumas mudanças. Em primeiro lugar, pode envolver uma fase de treino e uma fase de teste (p. ex., Wearden, 1991), tal como nos procedimentos com não-humanos, ou envolver apenas uma fase de teste (p. ex., Allan & Gibbon, 1991). Quando a fase de treino é implementada, o ensino da discriminação temporal pode ser realizado de duas formas, a primeira semelhante à fase de treino com não-humanos e a segunda recorrendo a instruções verbais que, obviamente, não são possíveis com não-humanos. Assim, na primeira forma, as duas durações de treino são apresentadas em tentativas sucessivas e, após cada apresentação, o participante emite uma de duas respostas possíveis (p. ex., pressiona uma de duas teclas do teclado do computador). As respostas corretas podem ser seguidas de reforço (tipicamente, o ganho de pontos) ou de um feedback indicando a duração apresentada na tentativa (por exemplo, a palavra 'curto' ou a palavra 'longo', consoante o estímulo modelo apresentado na tentativa). Na segunda forma de ensino da discriminação, o experimentador indica aos participantes a resposta correta a emitir após cada duração (Wearden, 1991). Por exemplo, depois de apresentar a duração Curta, o experimentar exibe na tela do computador a mensagem 'Este é o estímulo curto'; o mesmo acontece depois de apresentar a duração Longa.
Nos estudos com fase de treino, a fase de teste inclui durações intermediárias às durações de treino e é pedido ao participante que diga se o estímulo apresentado é mais semelhante à duração Curta ('neste caso, pressione a tecla 'curto'') ou à duração Longa ('neste caso, pressione a tecla 'longo''). As sessões de teste envolvem apresentações das durações de teste misturadas com apresentações das durações de treino mantendo-se, quando utilizado, o feedback para estas últimas. As condições para a passagem da fase de treino para a fase de teste serão descritas mais à frente, na secção sobre as variáveis dependentes.
Quando o procedimento envolve apenas uma fase de teste (p. ex., Allan & Gibbon, 1991), várias durações são apresentadas ao participante e a sua tarefa consiste em classificar essas durações em dois grupos, 'curto' ou 'longo'. O experimentador pode ainda dar feedback após as durações mais extremas.
Variáveis Independentes
Duração do Estímulo. Dado que o interesse central dos estudos de discriminação temporal é verificar se e como os sujeitos distribuem as suas respostas em função da duração de intervalos de tempo, a variável independente nestes estudos é a duração de um estímulo. A ordem de grandeza das durações é de segundos a poucos minutos nos estudos com não-humanos e de décimos de segundo a poucos segundos nos estudos com humanos. Esta redução das durações modelo nos estudos com humanos deve-se à necessidade de evitar que os participantes contem (silenciosamente, que seja) para estimar a passagem do tempo. Nos estudos com humanos que utilizam durações mais longas, a contagem encoberta pode ser evitada, pedindo ao participante para repetir números ou letras desordenadas durante o estímulo modelo ou, simplesmente, repetir em voz alta "blá-blá-blá...".
Modalidade do Estímulo. Alguns estudos têm procurado avaliar de que forma a modalidade do estímulo afeta a percepção da sua duração. Os estímulos utilizados podem ser visuais ou auditivos e podem também ser classificados como 'preenchidos' ou 'vazios'. No caso de intervalos preenchidos, o intervalo de tempo é marcado pela apresentação contínua de um estímulo (p. ex., uma luz que permanece acesa durante toda a duração do intervalo). No caso de intervalos vazios, o início do intervalo de tempo é sinalizado por um estímulo (p. ex., um clique) e o seu fim por outro estímulo (outro clique); o intervalo entre os estímulos não é preenchido com qualquer outro sinal.
As Variáveis Dependentes
Percentagem (ou Proporção) de Acertos. Retomemos o nosso exemplo. O animal aprendeu a responder 'curto' após estímulos de 1 s e a responder 'longo' após estímulos de 4 s. No final de cada sessão de treino, podemos calcular a percentagem (ou proporção) de tentativas em que o sujeito emitiu a resposta correta em função das durações apresentadas, 'curto' quando foi apresentado o estímulo de 1 s e 'longo' quando foi apresentado o estímulo de 4 s. O número de acertos aumenta gradualmente com o decorrer do treino e, em sujeitos não-humanos como ratos e pombos, atingem normalmente níveis superiores a 80% para cada uma das durações. Esta medida é utilizada para acompanhar o progresso da aprendizagem durante o treino da discriminação e, nos estudos com não-humanos, é com base nela que se define o critério para passar à fase de teste. Como critério de aprendizagem da discriminação utiliza-se, geralmente, um desempenho com pelo menos 80% de respostas corretas após cada uma das duas durações durante três (ou cinco) sessões consecutivas.
Em alguns estudos com humanos, a proporção de respostas corretas também é utilizada como critério para o avanço para a fase de teste (p. ex., Caselli et al., 2009; Spínola, Machado, Carvalho, & Tonneau, in press). No entanto, como a aprendizagem da discriminação por participantes humanos é bastante rápida, a passagem do treino para o teste é mais comumente definida pelo número de tentativas (p. ex., Wearden, 1991). Isto é, após a conclusão de um número pré-determinado de tentativas (p. ex., 100 tentativas, 50 com cada uma das duas durações modelo), é feito o teste, sem a adoção de um critério de aprendizagem. Quando o treino é realizado através de instruções verbais, a passagem do treino para o teste ocorre, geralmente, após cinco apresentações de cada duração modelo (p. ex., Wearden, 1991).
Proporção de Respostas. Um procedimento de bissecção temporal envolve, em cada tentativa, a emissão de uma de duas respostas, 'curto' ou 'longo'. Sendo assim, é possível analisar o desempenho do sujeito em termos da proporção de uma das respostas, por exemplo, 'longo', em função das durações do estímulo modelo. Obviamente, a proporção de respostas 'curto' para cada duração é complementar à proporção de respostas 'longo'.
Análise de Dados
Os resultados obtidos na fase de teste de um procedimento de bissecção temporal são, geralmente, apresentados sob a forma de uma função psicofísica ou função psicométrica. Esta função relaciona a proporção de uma das respostas (digamos, 'longo') com a duração do estímulo modelo. A função psicométrica pode ser concebida como um "relato" do modo como as durações do estímulo modelo são percebidas e classificadas pelo participante.
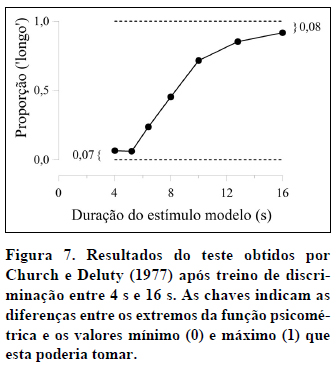
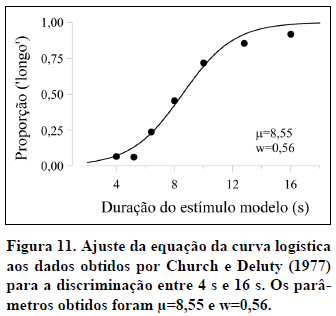
A título de exemplo, a Figura 3 apresenta os dados médios obtidos por Church e Deluty (1977) no teste após o treino da discriminação entre 4 s e 16 s. A função representada na figura é típica dos dados obtidos em tarefas de bissecção temporal.
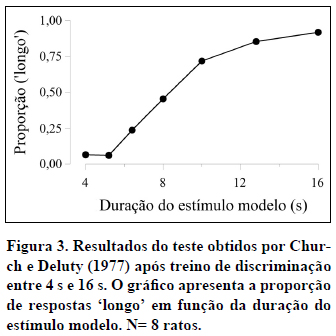
A função psicométrica tem quatro características principais. Em primeiro lugar, a sua forma é sigmóide, com a proporção de respostas 'longo' a aumentar de 0 para a duração Curta até 1 para a duração Longa. Em estudos com sujeitos não-humanos, os valores extremos de 0 e 1 raramente são observados, o que indica que os animais cometem alguns erros nas durações de treino.
Em segundo lugar, o ponto da função com ordenada igual a 0,5 identifica a duração que o sujeito classifica 50% das vezes como 'curto' e 50% das vezes como 'longo'. Essa duração, assinalada com uma seta na Figura 4, designa-se por ponto de igualdade subjetiva (PIS) ou ponto de bissecção, porque identifica a duração que subjetivamente divide ao meio o intervalo entre as durações Curta e Longa. Trata-se da duração julgada pelo sujeito como equidistante das duas durações de treino. Na medida em que essa duração ocasiona tantas respostas 'curto' quanto 'longo', também se designa por ponto de indiferença. O PIS pode ser estimado a partir dos dados por interpolação linear. Na interpolação linear calculamos a equação da reta que passa por dois pontos conhecidos (p. ex., os pontos [8; 0,45] e [10; 0,72] na Figura 4) e, em seguida, utilizamo-la para estimar a abcissa do ponto com ordenada 0,5. Na Figura 4, o PIS toma o valor de 8,35 s.
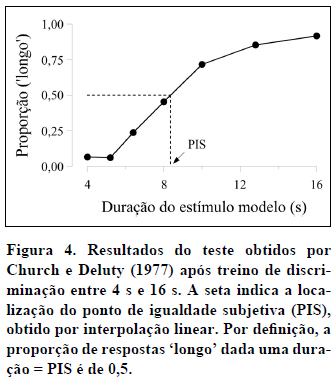
Para sujeitos não-humanos, o ponto de bissecção está geralmente próximo, não da média aritmética, mas da média geométrica das duas durações de treino (Catania, 1970; Church & Deluty, 1977; Fetterman & Killeen, 1991; Platt & Davis, 1983; Stubbs, 1968). A média geométrica de dois valores é igual à raiz quadrada do seu produto. No exemplo da Figura 4, a média geométrica é igual a 8=√(4*16), enquanto que a média aritmética é igual a 10=(4+16)/2. Com participantes humanos, a localização do ponto de bissecção não é tão clara. Alguns estudos revelaram pontos de bissecção na média geométrica (p. ex., Allan & Gibbon, 1991), mas outros revelaram pontos de bissecção na média aritmética das durações de treino (p. ex., Wearden, 1991).
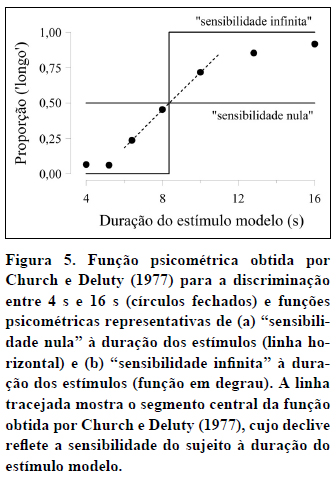
Em terceiro lugar, a "inclinação" do segmento da função próximo do PIS (ver linha tracejada na Figura 5) dá-nos uma medida da sensibilidade do sujeito à variação da duração do estímulo modelo. Para compreendermos melhor este ponto, notemos que um sujeito totalmente insensível à duração do estímulo modelo produziria uma função psicométrica horizontal, de declive 0 (ver Figura 5). Por outro lado, um sujeito que discriminasse perfeitamente todas as durações produziria uma função psicométrica em degrau, com todos os estímulos de duração inferior ao PIS a ocasionarem nenhuma resposta 'longo' e todos os estímulos de duração superior ao PIS a ocasionarem respostas 'longo'. Na duração correspondente ao PIS, a função daria um "salto" e teria, assim, um "declive infinito" (ver Figura 5). As funções reais estão entre estes dois extremos, com segmentos centrais mais próximos da horizontal nos sujeitos pouco sensíveis à duração e mais próximos da vertical nos sujeitos mais sensíveis à duração. Quanto maior a sensibilidade à dimensão temporal, maior o declive da função psicométrica no ponto de bissecção.
Para estimar o declive de uma função psicométrica específica, precisamos de conhecer as coordenadas de dois pontos. É frequente utilizar os pontos de coordenadas (t1,0,25) e (t2,0,75). Como as ordenadas desses pontos são dadas, basta-nos calcular as respetivas abcissas, as durações t1 e t2 assinaladas na Figura 6, o que pode ser feito por interpolação linear. No presente caso, t1=6,5 s e t2=10,7 s. O declive é estimado dividindo a diferença das ordenadas (0,75-0,25) pela diferença das abcissas (10,7-6,5), o que dá 0,12. Ou seja, neste caso, junto do PIS, a proporção de respostas 'longo' aumenta 0,12 pontos por cada segundo a mais do estímulo modelo.
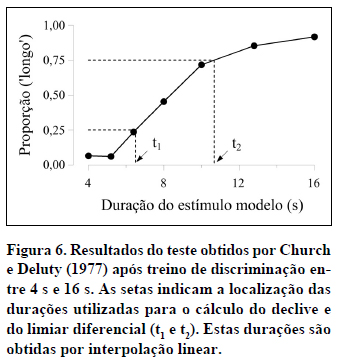
A sensibilidade é diretamente proporcional ao declive. Mas, como a diferença das ordenadas usadas para estimar o declive é constante e igual a 0,5, a sensibilidade depende unicamente da diferença entre t1 e t2. Quanto maior for essa diferença, menor é a "inclinação" da função entre esses dois pontos e, por isso, menor é a sensibilidade. Na psicofísica clássica, designa-se por limiar diferencial a diferença t2-t1 dividida por 2. Concluímos assim que as três quantidades, a sensibilidade do sujeito à duração do estímulo modelo, o declive da função no PIS, e o limiar diferencial, estão intimamente relacionadas, sendo a sensibilidade diretamente proporcional ao declive e inversamente proporcional ao limiar diferencial.
Em quarto lugar, os extremos da função psicométrica dão-nos informação sobre o nível de atenção do sujeito (i.e., o nível de controle de estímulo) durante a tarefa (Blough, 1996; Heinemann, Avin, Sullivan, & Chase, 1969). No caso que temos vindo a estudar, a proporção de respostas 'longo' atinge o seu valor mínimo, aproximadamente 0,07, para a duração Curta e o seu valor máximo, aproximadamente 0,92, para a duração Longa - ver Figura 7. A diferença entre o valor mínimo atingido, 0,07, e o menor valor possível da função, 0, é de 0,07, e a diferença entre o valor máximo atingido, 0,92, e o maior valor possível da função, 1, é de a 0,08. Ou seja, no total, 15% das respostas (0,07+0,08=0,15), aparentemente, não foram influenciadas pela duração dos estímulos. Desses 15%, 7% foram "palpites" de que o estímulo apresentado foi o 'longo' e 8% "palpites" de que o estímulo apresentado foi o 'curto' (Blough, 1996). Por outro lado, 85% das respostas (0,85=0,92-0,07) foram controladas pela duração dos estímulos.
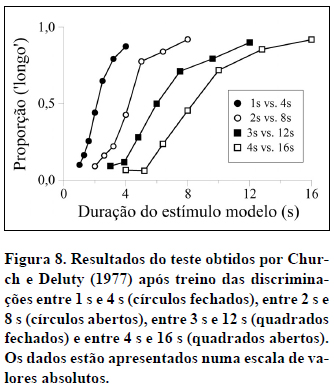
A Propriedade Escalar. Outras propriedades das funções psicométricas são reveladas quando comparamos os resultados obtidos com diferentes pares de durações de treino. Uma delas é a propriedade escalar: Sujeitos não-humanos ou participantes humanos treinados em procedimentos de bissecção temporal com durações de treino na mesma razão (p. ex., 1 s vs. 4 s e 4 s vs. 16 s - neste caso, durações na razão de 1 para 4), produzem funções psicométricas que se sobrepõem quando expressas numa escala relativa. Vejamos um exemplo a partir dos dados obtidos por Church e Deluty (1977). A Figura 8 mostra os dados de teste produzidos pelos mesmos 8 ratos após treinos de discriminação com 1 s e 4 s, 2 s e 8 s, 3 s e 12 s e 4 s e 16 s. Os dados estão representados numa escala de durações absolutas. Quando apresentadas desta forma, as curvas revelam diferentes declives e pontos de igualdade subjetiva.
Estes dados, no entanto, podem também ser representados numa escala relativa. Esta última pode ser obtida de várias maneiras, por exemplo, dividindo as durações usadas em cada teste pela média geométrica das duas durações usadas no treino correspondente, ou, se a razão das durações de treino se mantiver constante nos diferentes treinos, como no caso presente, dividindo as durações usadas em cada teste pela duração mais curta usada no treino correspondente. A Figura 9 mostra este último caso: Todas as durações de teste entre 1 s e 4 s foram divididas por 1 s, todas as durações entre 2 s e 8 s divididas por 2 s, todas as durações entre 3 s e 12 s divididas por 3 s e todas as durações entre 4 s e 16 s divididas por 4 s. Assim, as quatro funções psicométricas podem ser expressas numa escala comum entre 1 e 4. Nesta escala relativa, as quatro funções sobrepõem-se.
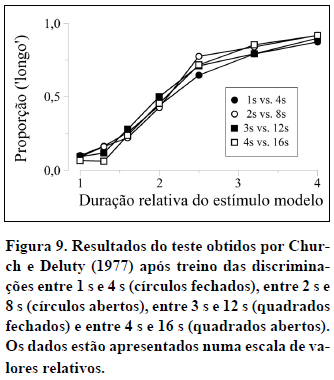
A sobreposição das funções psicométricas em escala relativa revela a propriedade escalar da percepção do tempo, talvez a propriedade mais importante do ponto de vista teórico. O que ela significa é que a discriminação de dois estímulos temporais depende da sua proporção e não dos seus valores absolutos (Church, 2003; Gibbon, 1977, 1991; Lejeune & Wearden, 2006).
A sobreposição das funções psicométricas é a expressão, no domínio temporal, da famosa lei de Weber. Tradicionalmente, a lei de Weber é apresentada da seguinte forma: O limiar diferencial ou JND (Just Noticeable Difference) associado a um estímulo padrão de magnitude t é proporcional a t, ou seja,

em que k>0 é a constante de proporcionalidade designada por fração de Weber. Assim, quando a lei de Weber é verdadeira para um dado contínuo perceptivo, a razão JND(t)/t é independente de t e característica do domínio.
Recorramos, como exemplo, à experiência original de Weber. Os participantes tinham de levantar, com uma mão, um objeto com um determinado peso fixo (o estímulo padrão) e depois levantar objetos de pesos variáveis. Posteriormente tinham de relatar se os pesos destes últimos eram maiores, menores ou iguais ao peso padrão. O objetivo da experiência era descobrir qual a diferença mínima necessária entre os pesos de dois objetos para estes serem julgados como diferentes. Weber verificou que, quando o peso padrão era de 40 g, um segundo objeto tinha de pesar em média 41 g para ser julgado mais pesado ou 39 g para ser julgado mais leve. Neste caso, com o peso padrão de 40 g, o limiar diferencial era de 1 g. Quando, por outro lado, o peso padrão era de 160 g, o peso do segundo objeto tinha de ser em média 164 g ou 156 g para este ser julgado mais pesado ou mais leve, respectivamente, do que o padrão. Ou seja, com o peso padrão de 160 g, o limiar diferencial era de 4 g. Weber descobriu, assim, que quando a magnitude do estímulo padrão aumentava (p. ex., de 40 g para 160 g), o limiar diferencial aumentava na mesma proporção (de 1 g para 4 g). A razão JND(t)/t mantinha-se constante - neste caso, 0,025 = 1/40 = 4/160.
Num procedimento de bissecção temporal, o PIS é o equivalente do estímulo padrão. Como o PIS reflete a duração julgada equidistante das duas durações de treino, os participantes escolhem 'curto' quando a duração do estímulo modelo é percebida como mais curta do que o PIS e escolhem 'longo' quando a duração do estímulo modelo é percebida como mais longa do que o PIS. Imaginemos agora que, numa tarefa de bissecção temporal com durações de treino de 1 s e 4 s, um sujeito escolhe 'longo' em 75% das vezes em que a duração do estímulo modelo é de 2,2 s e em 25% das vezes em que a duração do estímulo modelo é de 1,8 s. Se o PIS estiver na média geométrica das durações de treino, 2 s, o limiar diferencial seria, para este caso, de 0,2 s e a fração de Weber seria de k=0,1 (0,2/2,0). Se a fração de Weber se mantém constante para o domínio temporal3, numa discriminação entre 4 s e 16 s, o mesmo sujeito escolherá 'longo' em 75% das vezes em que a duração do estímulo modelo for de 8,8 s e em 25% das vezes em que a duração do estímulo modelo for de 7,2 s. Como o mesmo raciocínio se aplica ao adotarmos quaisquer outros índices de discriminabilidade para além de 75% e 25%, a lei de Weber para o tempo implica a sobreposição das funções psicométricas quando expressas em escala relativa.
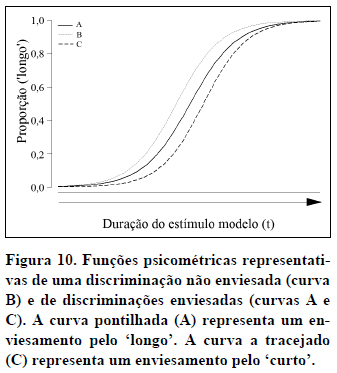
Por último, a função psicométrica também pode ser usada para detectar efeitos de certas variáveis na percepção temporal. Imagine, por exemplo, que uma droga como a metaanfetamina acelera a percepção do tempo (Meck, 1983). O efeito da droga revelar-se-á pelo deslocamento da função psicométrica para a esquerda (ver Figura 10) já que, sob efeito da droga, uma dada duração será percebida como mais longa do que sem esse efeito. O PIS será inferior aos PIS da condição controle (comparar curvas A e B na Figura 10). Drogas que tenham o efeito contrário sobre a percepção do tempo deslocarão a função para a direita e o PIS aumentará em relação à condição controle (comparar curvas B e C na Figura 10).
Resumindo, a análise de uma função psicométrica permite-nos obter informações sobre: (a) o grau de atenção do sujeito à dimensão ou propriedade do estímulo manipulada durante o procedimento experimental (no caso de uma bissecção temporal, à duração dos estímulos), (b) a sensibilidade do sujeito a variações nessa dimensão e (c) possíveis enviesamentos nas respostas (Blough, 1996; Heinemann et al., 1969).
Ajustamento Logístico. Os dados obtidos em tarefas de bissecção temporal são, regra geral, bem descritos por funções matemáticas e, por essa razão, a análise de dados costuma envolver o ajuste de funções aos dados brutos. A função logística e a função normal ou Gaussiana são frequentemente utilizadas para o efeito. O exemplo que se segue usa a função logística porque esta é mais fácil de tratar matematicamente. No caso mais simples, a função logística tem equação
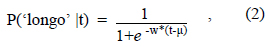
em que P('longo'|t) é a proporção de respostas 'longo' após um estímulo modelo de duração t, e µ e w são dois parâmetros livres, ambos positivos, estimados a partir dos dados.
A Figura 11 apresenta o ajuste da função logística aos dados de Church e Deluty (1977) para a discriminação entre 4 s e 16 s. A qualidade do ajuste é aferida pelo coeficiente de determinação (r2) - uma medida que varia entre 0 e 1, dependendo da qualidade do ajuste. Neste caso, r2 é igual a 0,98 o que legitima a intrepretação dos seus dois parâmetros. O parâmetro µ corresponde ao PIS. De fato, de acordo com a Equação (2), quando o estímulo modelo tem a duração t=µ s, então P('longo'|µ)=0,5. Na Figura 11, µ=8,55 .O parâmetro w é diretamente proporcional ao declive da função no PIS4 (e inversamente proporcional ao limiar diferencial5). Por isso, w mede a sensibilidade do sujeito à duração. Na Figura 11, w=0,56.
Para que as duas funções psicométricas se sobreponham, a sua equação deve respeitar a seguinte condição,
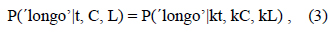
ou seja, a proporção de respostas 'longo' dado um estímulo de duração t e após um treino discriminativo com os estímulos C (curto) e L (longo) é igual à proporção de respostas'longo' dado um estímulo de duração k×t e após um treino discriminativo com os estímulos k×C (curto) e k×L (longo), com k>1. No exemplo que temos vindo a analisar, C=1 s, L=4 s e k=4, de modo que, neste caso, a condição acima exige que P('longo'|t, 1, 4) = P('longo'|4×t, 4, 16). Após o treino com 1 s e 4 s, a proporção de respostas 'longo' dado um estímulo de teste de t=2,5 s, por exemplo, deve ser igual à proporção de respostas 'longo' dado um estímulo de teste de t=10,0 s, após o treino com 4 s e 16 s. Formalmente, para que tal aconteça com a função logística, é necessário que os dois pares de parâmetros usados para ajustar as duas funções, um par referente às durações C e L (m1 e w1) e o outro par referente às durações k'C e k'L (m2 e w2), estejam relacionados da seguinte forma: m2=k×m1 e w2=w1/k. Quando C=1, L=4 e k=4, como no exemplo, então m2=4×m1 (com durações de treino 4 vezes maiores, o PIS é quatro vezes maior) e w2= w1/4 (com durações de treino 4 vezes maiores, o parâmetro de sensibilidade é quatro vezes menor).
Variações no Procedimento
O procedimento de bissecção temporal é utilizado para estudar de que forma sujeitos não-humanos e participantes humanos se comportam em função da duração dos estímulos. Dentro deste tema central, alguns pesquisadores exploraram a influência que diferentes características dos estímulos podem ter na percepção da sua duração. Os resultados têm mostrado que, para participantes humanos, estímulos auditivos tendem a ser percebidos como mais longos do que estímulos visuais (Wearden et al., 1998) e que intervalos preenchidos também parecem ser mais longos do que intervalos vazios (efeito de ilusão da duração preenchida; Wearden, Norton, Martin, & Montford-Bebb, 2007). Em estudos que procuram investigar o efeito das emoções na percepção temporal, os resultados mostram que a duração de estímulos que apresentam faces humanas com expressões de raiva é percebida como mais longa do que a duração de estímulos que apresentam faces com expressões neutras, ou com formas geométricas (p. ex., Gil & Droit-Volet, 2011).
Relações mais complexas entre estímulos temporais também têm sido estudadas utilizando o procedimento de bissecção temporal. Alguns pesquisadores têm-se questionado sobre se, nesta tarefa, não-humanos e humanos aprendem algo sobre a relação entre os dois estímulos. Uma tarefa de bissecção temporal envolve sempre uma relação de 'curto/longo' (ou 'menor/maior'). Neste sentido, alguns estudos têm mostrado que, depois de aprendida uma discriminação temporal entre dois estímulos, as durações Curta e Longa de um novo par de estímulos ocasionam as mesmas respostas que foram associadas à duração Curta e à duração Longa na primeira discriminação (Molet & Zentall, 2008; Zentall, Weaver, & Clement, 2004).
A mesma lógica foi também utilizada para pesquisar a discriminação da relação 'rápido/ lento' entre estímulos temporais formados por repetições de pulsos (Hulse & Kline, 1993) e os resultados também mostraram que pulsos temporais podem ser discriminados com base na sua rapidez relativa. No entanto, como outros estudos não encontraram evidência de representações relativas na tarefa de bissecção (Carvalho & Machado, 2012; Maia & Machado, 2009; Spínola et al., in press), a questão continua por ser esclarecida.
Algumas Aplicações do Procedimento
Para além das contribuições que tem dado para a compreensão dos processos básicos de percepção temporal em não-humanos e humanos, o procedimento de bissecção também tem sido aplicado em áreas como a psicofarmacologia e as neurociências. A utilização do procedimento nestas áreas tem permitido avaliar como é que a discriminação temporal é afetada por fármacos (p. ex., Meck, 1983; Odum, Lieving, & Schaal, 2002) e como se altera em doenças neurológicas como a doença de Huntington (p. ex., Brown et al., 2011), de Alzheimer (p. ex., Caselli et al., 2009), de Parkinson (p. ex., Merchant et al., 2008) e esquizofrenia (p. ex., Carroll et al., 2008).
Por exemplo, Merchant e colaboradores (2008) compararam o desempenho de doentes de Parkinson com adultos saudáveis (grupo controle) numa tarefa de bissecção temporal. Os dois grupos mostraram funções psicométricas com formas e declives semelhantes, mas a curva dos doentes de Parkinson estava deslocada ligeiramente para a direita quando comparada com curva do grupo controle. Tal resultado parece indicar que pacientes com a doença de Parkinson não apresentam uma sensibilidade temporal inferior àquela apresentada por adultos saudáveis, mas o deslocamento da curva indica que os pacientes perceberam o tempo como mais curto em comparação com adultos saudáveis.
Num estudo recente, Brown e colaboradores (2011) acompanharam a evolução da percepção temporal em ratos transgênicos, usados como modelo animal da doença de Huntington. Outros estudos (p. ex., Höhn et al., 2011) tinham já mostrado comprometimento da percepção temporal por volta dos quatro meses de vida neste modelo animal. Como as estruturas cerebrais que se vão degenerando na doença de Huntington são estruturas que se acredita estarem envolvidas no processamento da informação temporal, neste estudo os pesquisadores acompanharam o desempenho de ratos transgênicos numa tarefa de bissecção temporal dos 4 aos 8 meses de vida. Depois de um treino inicial numa discriminação entre 2 s e 8 s, foram conduzidos vários testes com durações intermediárias ao longo destes meses. Verificou-se que, aos quatros meses de idade, os ratos transgênicos apresentavam uma menor sensibilidade temporal do que ratos num grupo controle. Com a repetição dos testes, a sensibilidade temporal aumentou para os dois grupos de ratos (o declive das funções psicométricas aumentou), mas os ratos transgênicos pararam de responder mais cedo às durações intermediárias, revelando um aumento maior na sensibilidade temporal. Os pesquisadores sugeriram que outros fatores, não a sensibilidade temporal, contribuíram para o desempenho dos ratos na tarefa de bissecção temporal, uma vez que uma menor sensibilidade temporal aos 4 meses de idade seria preditiva de uma aprendizagem temporal mais lenta, contrariamente ao que foi observado. Mais especificamente, os pesquisadores sugeriram que um aumento da sensibilidade à extinção (ausência de reforço) nos ratos transgênicos poderia explicar porque razão eles pararam de responder mais depressa do que os ratos do grupo controle.
Explicações Teóricas (SET e LeT)
Com o objetivo de explicar os mecanismos que estão na base da percepção temporal em não-humanos e humanos e, de uma forma mais direta, integraros resultados obtidos em distintas tarefas temporais, diversos modelos teóricos foram desenvolvidos. Aqui resumimos o Scalar Expectancy Theory (SET), desenvolvido por John Gibbon, Russell Church e colaboradores (Gibbon, 1977, 1991; Gibbon, Church, & Meck, 1984) e o Learning-to-Time (LeT), desenvolvido por Armando Machado (Machado, 1997; ver Machado, Malheiro, & Erlhagen, 2009, para uma nova versão do modelo), este último com base nos trabalhos anteriores de Killeen e Fetterman (1988).
Scalar Expectancy Theory. O modelo SET (Figura 12, painel esquerdo) é um modelo cognitivo que postula que a informação temporal é processada por um relógio interno composto por três subunidades: (a) um pacemaker-acumulador, (b) uma ou mais memórias e (c) um mecanismo de tomada de decisão. O pacemaker gera pulsos continuamente a uma taxa variável e, quando se inicia a apresentação do estímulo temporal, estes pulsos começam a ser enviados para o acumulador. Quando termina a apresentação do estímulo modelo, o número de pulsos registrado no acumulador é armazenado numa memória. O modelo assume também que é formada uma memória diferente para cada duração aprendida. Assim, de acordo com o SET, numa tarefa de bissecção temporal são formadas duas memórias, uma para a duração Curta e outra para a duração Longa. No final da apresentação de cada duração, para decidir que resposta emitir, o SET propõe um mecanismo de tomada de decisão que toma em consideração o número de pulsos que está no acumulador no final do estímulo modelo e dois outros números, um número extraído aleatoriamente da memória de durações curtas e um número extraído aleatoriamente da memória de durações longas. Representemos por X o valor no acumulador, por C o valor extraído da memória para 'curto' e por L o valor extraído da memória para 'longo'. O SET propõe que a decisão se baseia nas razões X/C e L/X. Assim, se X/C <L/X, então o valor no acumulador está mais próximo do valor extraído da memória 'curto' e o sujeito responde 'curto'; se X/C> L/X, então o valor no acumulador está mais próximo do valor extraído da memória 'longo' e o sujeito responde 'longo'. Daqui se segue que o ponto de bissecção ocorre quando X/C = L/X, ou seja, quando X corresponde à média geométrica de C e L (X2=C×L, o que implica que X=√(C×L). O SET prevê ainda a sobreposição de funções psicométricas obtidas para pares de durações na mesma razão.
Learning-to-Time. O LeT (Figura 12, painel direito) é um modelo de cariz comportamental que postula três componentes principais: (a) uma organização em série de estados comportamentais, (b) um conjunto de elos associativos que ligam os estados comportamentais às respostas, e (c) as próprias respostas. Quando o estímulo temporal é iniciado, somente o primeiro estado comportamental está ativo mas, com a passagem do tempo, os estados subsequentes vão sendo ativados em série. Para além disso, todos os estados comportamentais estão ligados às respostas operantes por elos associativos. No início do treino, todos estes elos têm a mesma força associativa mas, durante o treino, a força associativa altera-se. Quando um determinado estado comportamental está ativo e o sujeito emite uma resposta que é reforçada, o elo que liga esse estado a essa resposta é fortalecido e os elos que ligam esse estado a outras respostas são enfraquecidos. Por outro lado, quando o sujeito emite uma resposta que não é reforçada, o elo que liga o estado comportamental ativo no momento da resposta a essa resposta é enfraquecido. Assim sendo, a escolha do sujeito depende, a cada momento, do estado comportamental mais ativo (o que, por sua vez, depende da duração do estímulo modelo) e das forças relativas dos elos que ligam esse estado às respostas (o que, por sua vez, depende de processos de aprendizagem que envolvem reforçamento e extinção). A resposta com elos mais fortes tem maior probabilidade de ser emitida.
Numa bissecção temporal, no final do treino, os estados comportamentais iniciais, mais ativos após a duração Curta, vão estar fortemente ligados à resposta 'curto' e fracamente ligados à resposta 'longo' e os estados comportamentais posteriores, mais ativos após a duração Longa, vão estar fortemente ligados à resposta 'longo' e fracamente ligados à resposta 'curto'. Estas forças associativas decorrem do reforçamento e da extinção das respostas corretas e incorretas, respectivamente. Deste modo, o animal responde 'curto' após durações curtas e responde 'longo' após durações longas. Durante o teste, após a apresentação de estímulos intermediários mais próximos da duração Curta, os estados comportamentais mais ativos vão ser os iniciais e por isso o sujeito tem maior probabilidade de responder 'curto'. Após a apresentação de estímulos mais próximos da duração Longa, os estados comportamentais mais ativos vão ser os finais e por isso o sujeito tem maior probabilidade de responder 'longo'. Assim, o LeT prevê, tal como o SET, que a proporção de respostas 'longo' aumente com a duração do estímulo modelo. O LeT também prevê que o ponto de bissecção ocorra próximo da média geométrica das durações de treino e que as funções psicométricas se sobreponham (ver Machado, 1997; e Machado et al., 2009).
O SET e o LeT são dois dos modelos de timing mais estudados recentemente. Mas uma diversidade de modelos6 foi avançada nas últimas décadas por diferentes grupos de pesquisadores, como por exemplo, o Behavioral Theory of Timing (BeT; Killeen & Fetterman, 1988) e o Behavioral Economic Model (BEM; Jozefowiez, Staddon, & Cerutti, 2009). Cada modelo sugere processos distintos de regulação temporal e sublinha diferentes conjuntos de dados empíricos (Lejeune et al., 2006; Machado et al., 2009).
Descortinamos assim no estudo do timing três fases distintas, a primeira dominada pela exploração do plano empírico, dos procedimentos e técnicas de estudo; a segunda dominada pela exploração do plano teórico, dos modelos quantitativos e suas implicações; e a terceira, ainda na sua infância, dominada pelo teste empírico dos diferentes modelos, do confronto entre eles e os dados da experimentação (p. ex., Arantes, 2008; Arantes & Machado, 2008; Carvalho & Machado, 2012; Machado & Arantes, 2006; Machado & Keen, 1999; Machado & Oliveira, 2009; Machado & Pata, 2005; Pinto & Machado, 2011; Vieira de Castro & Machado, 2012). A exploração do domínio empírico, ilustrada neste artigo com o procedimento de bissecção, interage com a exploração do domínio teórico, ilustrada neste artigo com dois modelos quantitativos, e sobre ela repousa o progresso científico.
Referências
Allan, L. G., & Gibbon, J. (1991). Human bisection at the geometric mean. Learning and Motivation, 22,39-58. doi: 10.1016/0023-9690(91)90016-2 [ Links ]
Anderson, A. C. (1932). Time discrimination in the white rat. Journal of Comparative Psychology, 13,25-55. doi: 10.1037/h0074562 [ Links ]
Arantes, J. (2008). Comparison of Scalar Expectancy Theory (SET) and the Learning to-Time (LeT) model in a successive temporal bisection task. Behavioural Processes, 78,269-278. doi: 10.1016/j.beproc.2007.12.008 [ Links ]
Arantes, J., & Machado, A. (2008). Context effects in a temporal discrimination task: Further tests of the Scalar Expectancy Theory and Learning-to-Time models. Journal of the Experimental Analysis of Behavior, 90,33-51. [ Links ]
Blough, D. S. (1996). Error factors in pigeon discrimination and delayed matching. Journal of Experimental Psychology: Animal Behavior Processes, 22,118-131. doi: 10.1037/0097-7403.22.1.118 [ Links ]
Boring, E. G. (1957). A history of experimental psychology (2nd ed.). New York: Prentice Hall. [ Links ]
Brown, B. L., Höhn, S., Faure, A., von Hörsten, S., LeBlanc, P., Desvignes, N., ...Doyère, V. (2011). Temporal sensitivity changes with extended training in a bisection task in a transgenic rat model. Frontiers in Integrative Neurosciences, 5,1-7. doi: 10.3389/fnint.2011.00044 [ Links ]
Carroll, C. A., Boggs, J., O'Donnell, B. F., Shekhar, A., & Hetrick, W. P. (2008). Temporal processing dysfunction in schizophrenia. Brain and Cognitions, 67,150-161. doi: 10.1016/j. bandc.2007.12.005 [ Links ]
Carvalho, M. P., & Machado, A. (2012). Relative versus absolute stimulus control in the temporal bissection task. Journal of the Experimental Analysis of Behavior, 98,23-44. [ Links ]
Caselli, L., Iaboli, L., & Nichelli, P. (2009). Time estimation in mild Alzheimer's disease patients. Behavioral and Brain Functions, 5. Retrieved from http://www.behavioralandbrainfunctions.com/content/pdf/1744-9081-5-32.pdf [ Links ]
Catania, A. C. (1970). Reinforcement schedules and the psychophysical judgments: A study of some temporal properties of behavior. In W. N. Schoenfeld (Ed.), The theory of reinforcement schedules (pp. 1-42). New York: Appleton-Century-Crofts. [ Links ]
Church, R. M. (2002). Temporal Learning. In H. Pashler & C. R. Gallistel (Eds.), Stevens' handbook of experimental psychology: Vol. 3. Learning, Motivation, and Emotion (pp. 365-393). New York: Wiley. [ Links ]
Church, R. M. (2003). A concise introduction to scalar timing theory. In H. W. Meck (Ed.), Functional and neural mechanisms of interval timing (pp. 3-22). Boca Raton, FL: CRC Press. [ Links ]
Church, R. M., & Deluty, M. Z. (1977). Bisection of temporal intervals. Journal of Experimental Psychology: Animal Behavior Processes, 3,216-228. doi: 10.1037/0097-7403.9.2.160 [ Links ]
Church, R. M., Getty, D. J., & Lerner, N. D. (1976). Duration discrimination by rats. Journal of Experimental Psychology: Animal Behavior Processes, 2,303-312. doi: 10.1037/00977403.2.4.303 [ Links ]
Cowles, J. T., & Finan, J. L. (1941). An improved method for establishing temporal discrimination in white rats. Journal of Psychology: Interdisciplinary and Applied, 11,335-342. doi: 10.1080/ 00223980.1941.991.9917040 [ Links ]
Crystal, J. D. (2002). Timing inter-reward intervals. Learning and Motivation, 33,311-326. doi: 10.1016/S0023-9690(02)00005-X [ Links ]
Droit-Volet, S., & Wearden, J. H. (2001). Temporal bisection in children. Journal of Experimental Child Psychology, 80,142-159. doi: 10.1006/ jecp.2001.2631 [ Links ]
Ferrara, A., Lejeune, H., & Wearden, J. H. (1997). Changing sensitivity to duration in human scalar timing: An experiment, a review and some possible explanations. Quarterly Journal of Experimental Psychology, 50,217-237. doi: 10.1080/713932654 [ Links ]
Ferster, C. B., & Skinner, B. F. (1957). Schedules of Reinforcement. New York: Appleton-Century-Crofts. [ Links ]
Fetterman, J. G., & Killeen, P. R. (1991). Adjusting the pacemaker. Learning and Motivation, 22,226-252. doi: 10.1016/0023-9690(91)90024-3 [ Links ]
Gallistel, C. R. (1993). The organization of learning. Cambridge, MA: Massachusetts Institute of Technology Press. [ Links ]
Gibbon, J. (1977). Scalar expectancy theory and Weber's law in animal timing. Psychological Review, 84,279-325. doi: 10.1037/0033295X.84.3.279 [ Links ]
Gibbon, J. (1991). Origins of scalar timing theory. Learning and Motivation, 22,3-38. doi: 10.1016/0023-9690(91)90015-Z [ Links ]
Gibbon, J., Church, R. M., & Meck, W. H. (1984). Scalar timing in memory. In L. Allan & J. Gibbon (Eds.), Timing and time perception (pp. 5277). New York: Annals of the New York Academy of Sciences. [ Links ]
Gil, S., & Droit-Volet, S. (2011). "Time flies in the presence of angry faces"... depending on the temporal task used! Acta Psychologica, 136,354-362. doi: 10.1016/j.actpsy.2010.12.010 [ Links ]
Grondin, S. (2010). Timing and time perception: A review of recent behavioral and neuroscience findings and theoretical directions. Attention, Perception, & Psychophysics, 72,561-582. doi: 10.3758/APP.72.3.56 [ Links ]
Heinemann, E. G., Avin, E., Sullivan, M. A., & Chase, S. (1969). Analysis of stimulus generalization with a psychophysical method. Journal of Experimental Psychology, 80,215-224. doi: 10.1037/h0027309 [ Links ]
Heron, W. T. (1949). Time discrimination in the rat. Journal of Comparative and Physiological Psychology, 42,27-31. doi: 10.1037/h0054217 [ Links ]
Höhn, S., Dallérac, G., Faure, A., Urbach, Y., Nguyen, H. P., Riess, O., ...Doyère, V. (2011). Behavioral and in vivo electrophysiological evidence for presymptomatic alteration of prefronto-striatal processing in the transgenic rat model for Huntington disease. Journal of Neuroscience, 31,8986-8997. doi: 10.1523/JNEUROSC.1238-11.2011 [ Links ]
Hulse, S. H., & Kline, C. L. (1993).The perception of time relations in auditory tempo discrimination. Animal Learning and Behavior, 21,281-288. doi: 10.3758/BF03197992 [ Links ]
Jozefowiez, J., Staddon, J. E. R., & Cerutti, D. T. (2009). The behavioral economics of choice and interval timing. Psychological Review, 116,519-539. doi: 10.1037/a0016171 [ Links ]
Killeen, P. R., & Fetterman, J. G. (1988). A behavioral theory of timing. Psychological Review, 95,274-295. doi: 10.1037/0033-295X.95.2.274 [ Links ]
Lejeune, H., & Wearden, J. H. (2006). Scalar properties in animal timing: Conformity and violations. The Quarterly Journal of Experimental Psychology, 59,1875-1908. [ Links ]
Lejeune, H., Richelle, M., & Wearden, J. H. (2006). About Skinner and time: Behavior-analytic contributions to research on animal timing. Journal of the Experimental Analysis of Behavior, 85,125-142. doi: 10.1901/jeab.2006.85.04 [ Links ]
Machado, A. (1997). Learning the temporal dynamics of behavior. Psychological Review, 104,241-265. doi: 10.1037/0033-295X.104.2.241 [ Links ]
Machado, A., & Arantes, J. (2006). Further tests of the scalar expectancy theory (SET) and the learning-to-time (Let) model in a temporal bisection task. Behavioural Processes, 72,195-206. doi: 10.1016/j.beproc.2006.2006.03.001 [ Links ]
Machado, A., & Keen, R. (1999). Learning to Time (LET) or Scalar Expectancy Theory (SET)? A critical test of two models of timing. Psychological Science, 10,285-290. doi: 10.1111/14679280.00152 [ Links ]
Machado, A., & Oliveira, L. (2009). Dupla bissecção temporal: Testes críticos de dois modelos de timing. Acta Comportamentalia, 17,25-60. [ Links ]
Machado, A., & Pata, P. (2005). Testing the scalar expectancy theory (SET) and the learning-to-time model (LeT) in a double bisection task. Learning and Behaviour, 33,111-122. doi: 10.3758/ BF03196055 [ Links ]
Machado, A., Malheiro, M. T., & Erlhagen, W. (2009). Learning to time: A perspective. Journal of the Experimental Analysis of Behavior, 92,423-458. doi: 10.1901/jeab.2009.92-423 [ Links ]
Maia, S., & Machado, A. (2009). Representation of time intervals in a double bisection task: Relative or absolute? Behavioural Processes, 81,280-285. doi: 10.1016/j.beproc.2008.10.012 [ Links ]
McCormack, T., Brown, G. D. A., Maylor, E. A., Darby, R. J., & Green, D. (1999). Developmental changes in time estimation: Comparing childhood and old ages. Developmental Psychology, 35,1143-1155. doi: 10.1037/00121649.35.4.1143 [ Links ]
Meck, W. H. (1983). Selective adjustment of the speed of internal clock and memory processes. Journal of Experimental Psychology: Animal Behavior Processes, 9,171-201. doi: 10.1037/0097-7403.9.2.171 [ Links ]
Merchant, H., Luciana, M., Hooper, C., Majestic, S., & Tuite, P. (2008). Interval timing and Parkinson's disease: Heterogeneity in temporal performance. Experimental Brain Research, 184,233-248. doi: 10.1007/s00221-007-1097-7 [ Links ]
Michon, J. A. (1993). Concerning the time sense: The seven pillars of time psychology. Psychologica Belgica, 33,329-345. [ Links ]
Molet, M., & Zentall, T. R. (2008). Relative judgments affect assessments of stimulus duration. Psychonomic Bulletin & Review, 15,431-436. doi: 10.3758/PBR.15.2.431 [ Links ]
Odum, A. L., Lieving, L. M., & Schaal, D. W. (2002). Effects of d-amphetamine in a temporal discrimination procedure: Selective changes in timing or rate dependency? Journal of the Experimental Analysis of Behavior, 78,195-214. doi: 10.1901/jeab.2002.78-195 [ Links ]
Pavlov, I. P. (1960). Conditioned refl exes: An Investigation of the physiological activity of the cerebral cortex (G. V. Anrep, Trans.). New York: Dover. (Original work published 1927) [ Links ]
Pinto, C., & Machado, A. (2011). Short-term memory for temporal intervals: Contrasting explanations of the choose-short effect in pigeons. Learning and Motivation, 42,13-25. doi: 10.1016/j. lmot.2010.05.001 [ Links ]
Platt, J. R., & Davis, E. R. (1983). Bisection of temporal intervals by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 9,160-170. doi: 10.1037/0097-7403.9.2.160 [ Links ]
Richelle, M., & Lejeune, H. (1980). Time in Animal Behavior. Oxford, UK: Pergamon Press. [ Links ]
Roberts, W. A. (1998). Principles of animal cognition. New York: McGraw-Hill. [ Links ]
Sams, C. F., & Tolman, E. C. (1925). Time discrimination in white rats. Journal of Comparative and Physiological Psychology, 5,255-263. doi: 10.1037/h0074886 [ Links ]
Shettleworth, S. J. (1998). Cognition, evolution, and behavior. New York: Oxford University Press. [ Links ]
Skinner, B. F. (1938). The behavior of organisms: An experimental analysis. Oxford, England: Appleton-Century. [ Links ]
Spínola, I., Machado, A., Carvalho, M. P., & Tonneau, F. (in press). What do humans learn in a double, temporal bisection task: Absolute or relative stimulus durations? Behavioural Processes. [ Links ]
Stubbs, D. A. (1968). The discrimination of stimulus duration by pigeons. Journal of the Experimental Analysis of Behavior, 11,223-238. doi: 10.1901/jeab.1968.11-223 [ Links ]
Titchener, E. B. (1915). A beginner's psychology. New York: Macmillan. [ Links ]
Vieira de Castro, A. C., & Machado, A. (2012). The interaction of temporal generalization gradients predicts the context effect. Journal of the Experimental Analysis of Behavior, 97,263-279. [ Links ]
Wearden, J. H. (1991). Human performance on an analogue of the interval bisection task. Quarterly Journal of Experimental Psychology: Comparative and Physiological Psychology, 43,59-81. doi: 10.1080/14640749108401259 [ Links ]
Wearden, J. H., & Ferrara, (1995). Stimulus spacing effects in temporal bisection by humans. Quarterly Journal of Experimental Psychology, 48,289-310. doi: 10.1080/14640749508401454 [ Links ]
Wearden, J. H., Edwards, H., Fakhri, M., & Percival, A. (1998). Why "sounds are judged longer than lights": Application of a model of the internal clock in humans. Quarterly Journal of Experimental Psychology, 51,97-120. doi: 10.1080/713932672 [ Links ]
Wearden, J. H., Norton, R., Martin, S., & Montford-Bebb, O. (2007). Internal clock processes and the filled duration illusion. Journal of Experimental Psychology: Human Perception and Performance, 33,716-729. doi: 10.1037/00961523.33.3.716 [ Links ]
Woodrow, H. (1928). Temporal discrimination in the monkey. Journal of Comparative Psychology, 8,395-427. doi: 10.1037/h0073047 [ Links ]
Yagi, B. (1962). The effect of motivating conditions on "the time estimation" in rats. Japanese Journal of Psychology, 33,8-24. [ Links ]
Zeiler, M. D., & Powell, D. G. (1994). Temporal control in fixed-interval schedules. Journal of the Experimental Analysis of Behavior, 61,1-9. doi: 10.1901/jeab.1994.61-1 [ Links ]
Zentall, T. R., Weaver, J. E., & Clement, T. S. (2004). Pigeons group time intervals according to their relative duration. Psychonomic Bulletin & Review, 11,113-117. doi: 10.3758/BF03206469 [ Links ]
 Endereço para correspondência:
Endereço para correspondência:
Escola de Psicologia, Universidade do Minho
Braga, Portugal 4710
E-mail: castro.anacatarina@gmail.com
Recebido: 18/07/2012
1ª revisão: 29/11/2012
Aceite final: 06/12/2012
Todos os autores foram apoiados pela Fundação para a Ciência e Tecnologia (FCT).
1 A designação de bissecção temporal (temporal bisection em inglês) surge no estudo de Church e Deluty (1977).
2 A repetição da tentativa em caso de erro assegura que, no fim de cada sessão, o número de reforços recebidos por responder na barra esquerda é igual ao número de reforços recebidos por responder na barra direita, o que contribui para reduzir o enviesamento por uma das respostas.
3 Apesar de a grande maioria dos resultados experimentais favorecerem a lei de Weber (p. ex., Allan & Gibbon, 1991; Wearden, 1991, com participantes humanos; Church & Deluty, 1977, com ratos; Platt & Davis, 1983; Stubbs, 1968, com pombos), a constância da fração de Weber não é uma regra absoluta para discriminações temporais (p. ex., Ferrara, Lejeune, & Wearden, 1997; Wearden, Edwards, Fakhri, & Percival, 1998; com participantes humanos; Church, Getty, & Lerner, 1976; Crystal, 2002; Zeiler & Powell, 1994, com não-humanos).
4 A derivada da função P('longo'|t) avaliada no ponto t=µ é igual a w/4.
5 Quando o limiar diferencial é calculado como acima referido, ou seja, a partir das durações correspondentes às proporções de 0,75 e 0,25, então o limiar diferencial é igual a ln(3)/w em que ln(3) é o logaritmo neperiano de 3.
6 A lista de modelos de timing aqui apresentada não é exaustiva. Para uma lista mais completa, ver Machado et al., 2009.

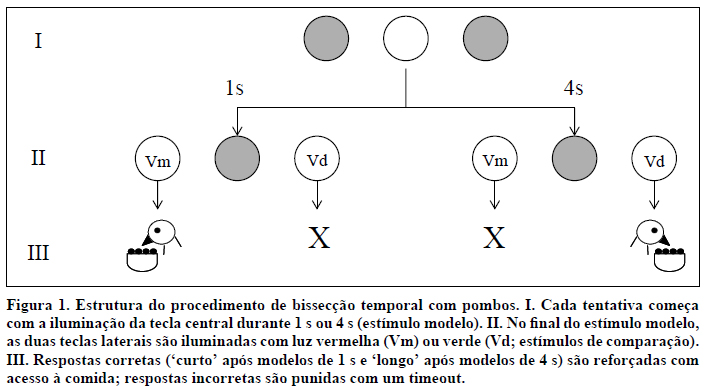{kind=link}
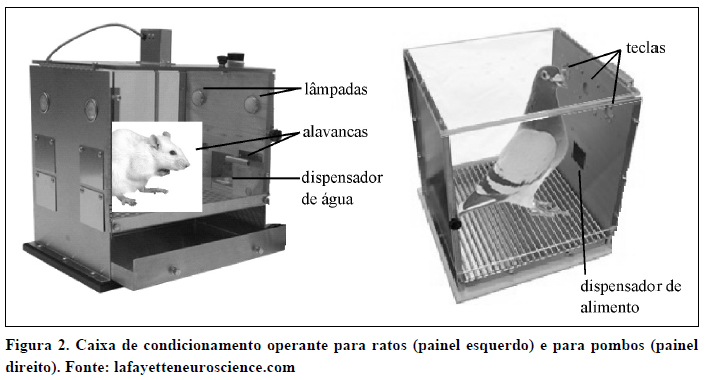{kind=link}
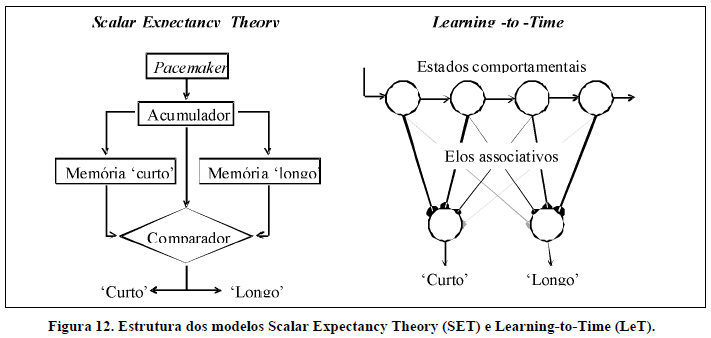{kind=link}