










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.6 n.1 São Paulo jun. 2004
ARTIGOS
Amamentação da prole e amamentação cooperativa em catetos (Tayassu tajacu): uma análise exploratória*
Offspring and cooperative nursing in collared peccaries (Tayassu tajacu): an exploratory analysis
Cibele Biondo; Vera Silvia Raad Bussab**
Universidade de São Paulo
RESUMO
Sendo a amamentação uma atividade custosa, esperar-se-ia que as fêmeas de mamíferos apenas cedessem seu leite aos próprios filhotes. O comportamento de amamentar filhotes alheios foi, contudo, observado em muitas espécies de mamíferos. Neste estudo, investigou-se a amamentação em catetos com especial atenção aos padrões de amamentação cooperativa em contraste com os de amamentação da própria prole. Foram observados quatro fêmeas e seis filhotes de um grupo de 26 catetos mantidos em cativeiro, utilizando os métodos de animal focal e varredura instantânea. Todas as fêmeas amamentaram filhotes alheios, porém a freqüência de amamentação cooperativa foi menor do que a esperada para catetos. A amamentação cooperativa e a dos próprios filhotes foram caracterizadas em termos da aceitação da fêmea, do número de filhotes mamando, da posição dos filhotes, e do modo de finalização das mamadas. A maioria dessas características foi semelhante nos dois tipos de amamentação.
Descritores: Amamentação cooperativa, Comportamento materno, Catetos, Tayassu tajacu.
ABSTRACT
Being a high-cost activity, nursing should be restricted to own young. The behavior of nursing alien offspring has however been observed in many mammal species. In this study, nursing was observed in collared peccary, with special attention to the patterns of cooperative nursing in contrast to offspring nursing. Four females and six young of a group of 26 collared peccaries were observed in captivity, throughout focal animal and scan sampling methods. All of the females nursed alien young, but the frequency of cooperative nursing was lower than that expected for collared peccaries. Offspring and cooperative nursing events were assessed in terms of female acceptance, number of young sucking, young postures, and finalization patterns of nursing bouts. Most of such features were similar in offspring and cooperative nursing.
Index terms: Cooperative nursing, Maternal behavior, Collared peccaries, Tayassu tajacu
Sabe-se que a lactação é custosa em termos metabólicos e que pode aumentar significativamente as taxas de mortalidade das fêmeas de mamíferos (Clutton-Brock, Albon, & Guiness, 1989). Um recurso do valor do leite deve, a rigor, ser regulado em termos de custos e benefícios de acordo com a lógica evolucionária. A partir deste raciocínio, seria de se esperar que fêmeas fornecessem o leite exclusivamente para sua própria prole. Contudo, a amamentação de proles alheias tem sido descrita na maioria dos grandes grupos taxonômicos de mamíferos, como Artiodactyla, Carnivora, Cetacea, Chiroptera, Perissodactyla, Pinnipedia, Primata (incluindo humanos, por exemplo, Tronick, Morelli, & Winn, 1987), Proboscidea e Rodentia (Packer, Lewis, & Pusey, 1992).
A terminologia para designar este tipo de amamentação é variada, em virtude das especificidades do fenômeno em cada espécie. Pode-se citar alguns termos: amamentação de não-prole (non-offspring nursing, Packer et al., 1992); amamentação comunitária (communal nursing, Hoogland, Tamarin, & Levy, 1989); alo-amamentação (allonursing, Roulin, 2002). Neste trabalho, será utilizado o termo amamentação cooperativa (cooperative nursing, Byers & Bekoff, 1981), que foi o termo empregado na literatura para a espécie em questão, Tayassu tajacu. Para simplificar, será utilizada a sigla AC para designar a amamentação cooperativa e a sigla APP para a amamentação de própria prole.
Packer et al. (1992) verificaram que a AC parece ser mais freqüente em estudos com animais cativos do que com animais selvagens, sendo até considerada um artefato da perturbação humana ou do próprio confinamento por alguns autores (por exemplo, Fogden, 1971). Características do cativeiro, como a quantidade ilimitada de alimento, poderiam diminuir os custos da doação de leite para filhotes alheios, favorecendo a ocorrência desse tipo de amamentação.
Aspectos da organização social das espécies têm sido associados com a AC, constatando-se interação entre os fatores tamanho da ninhada e tamanho do grupo. Espécies uníparas apresentam menos AC do que espécies plurípa-ras. Prováveis explicações aventadas são: nas espécies uníparas, a amamentação simultânea dos próprios filhotes e dos alheios requer razões de consumo de comida consideravelmente maiores; a amamentação continuada de filhotes alheios, não associada à do próprio filhote, por emancipação ou morte deste, prorrogaria a lactação e poderia adiar reprodução subseqüente (por exemplo, Altmann, Altmann, & Hausfater, 1978, para primatas). Por outro lado, a AC poderia implicar em custos mais baixos para as espécies pluríparas, em especial nos casos de ninhadas próprias menores do que o limite da espécie.
Exceções entre uníparas ocorrem nas espécies que vivem em grupos grandes. Um raciocínio explicativo possível está relacionado ao fato de que, nesses grupos, o aumento de exposição a filhotes alheios possibilite a ocorrência de furtos de leite (Packer et al., 1992), pela dificuldade que as mães teriam de reconhecer e localizar sua própria prole ou de afastar filhotes alheios. Exemplos disso são dados nas espécies que formam grandes colônias reprodutivas. As fêmeas de morcegos mexicanos deixam seus filhotes em cavernas contendo milhões de infantes e voltam para amamentar duas vezes por dia. Encontram muitos filhotes alheios ávidos para mamar e, embora rejeitem muitas dessas tentativas, acabam perdendo uma certa quantidade de leite para eles (McCracken, 1984). Com fêmeas de elefantes marinhos, acontece algo parecido (Reiter, Stinson, & Le Boeuf, 1978). Entretanto, este raciocínio não se aplica ao caso das espécies pluríparas, nas quais a ocorrência deste comportamento é mais freqüente naquelas que se organizam em grupos pequenos. Neste último caso, a seleção de parentesco seria um dos fatores mais importantes na ocorrência de AC, já que se espera um maior grau de parentesco entre as fêmeas nos grupos pequenos (Packer et al., 1992).
De acordo com essas hipóteses, os catetos reúnem as características associadas à ocorrência do fenômeno, pois vivem geralmente em grupos pequenos de indivíduos aparentados e que podem ter ninhadas de mais de um filhote. Além disso, são animais bastante sociais e cooperativos, sendo que os adultos são bem tolerantes com os juvenis (Byers & Bekoff, 1981). A AC foi mencionada em catetos por Byers e Bekoff (1981), porém nenhum estudo sistemático foi realizado para este comportamento nesta espécie.
Este estudo visou explorar a AC e a APP em catetos, buscando descrever e comparar seus padrões com base em características como a freqüência e duração dos eventos, aceitação das fêmeas às solicitações dos filhotes para mamar, número de filhotes mamando por fêmea, posição dos filhotes para mamar, e modo de finalização dos eventos. Acredita-se que ao investigar o efeito de fatores determinantes proximais/causais seja possível obter informações que possam ajudar a esclarecer os processos distais/funcionais de adaptação desta espécie e na compreensão dos aparentes paradoxos relacionados à AC.
Método
Local de estudo e animais
O estudo foi conduzido no criadouro comercial da Fazenda Vale Verde, uma propriedade particular no Município de São Miguel Arcanjo (SP), nos meses de janeiro e fevereiro de 2001. Os animais observados estavam confinados em um recinto de área arborizada de 2.600 m2, com um cocho coberto e um tanque de água. No total eram 26 indivíduos: 15 adultos (três machos e 12 fêmeas), cinco juvenis (cinco fêmeas) e seis filhotes (três machos e três fêmeas). A alimentação era fornecida no início da manhã e final da tarde, e compunha-se de ração de fabricação própria da Fazenda, à base de milho, soja, trigo e minerais.
Procedimento
Os animais, previamente marcados com tosa no pêlo, foram observados do centro do recinto com auxílio de um gravador portátil em períodos contínuos de duas horas. As observações foram feitas durante 22 dias consecutivos.
O método utilizado para obtenção das freqüências de amamentação cooperativa foi o de grupo focal (Setz, 1991), composto por uma fêmea determinada e pelos filhotes que ela estava amamentando. Foram observadas quatro fêmeas focais e seus filhotes. Cada observação focal durava dez minutos. Para determinar a ordem de observação dos focais, as fêmeas eram sorteadas aleatoriamente. Paralelamente, ao fim de cada focal, era feita uma varredura ("scan", Setz, 1991) de dois minutos para registrar as atividades de todos os filhotes observados.
Em cada evento de amamentação foram registrados: a ocorrência de solicitação do filhote para mamar e a reação da fêmea; o número e a identidade dos filhotes em amamentação postura de mamada do filhote (em pé ou deitado) e posição em relação à fêmea (por entre as patas traseiras dela, perpendicular a ela ou embaixo dela); postura da fêmea (em pé ou com as patas traseiras arqueadas); responsável pelo término do episódio ( fêmea ou filhote); e a duração em segundos.
Os dados amostrados foram analisados usando-se o pacote estatístico SPSS (Statistical Package for Social Sciences).
Resultados
Amostraram-se 671 eventos de amamentação efetiva, dos quais, 650 foram de APP (96,9%) e 22 de AC (3,1%); com predominância significativa da APP (X2 (1, N = 672) = 586,88; p < 0,001).
Todas as fêmeas observadas amamentaram cooperativamente, sendo as fêmeas D e A, as que amamentaram com maior freqüência e maior diversidade de filhotes (Tabela 1). A fêmea D amamentou cooperativamente os filhotes a2, b1 e c; e a fêmea A, os filhotes b1, b2 e c. A fêmea C só amamentou, em cooperação, fêmeas juvenis que não faziam parte da amostra original. A fêmea B amamentou os filhotes a1 e a2. O filhote b1 foi amamentado cooperativamente com maior freqüência. O filhote d não foi amamentado por fêmeas alheias.
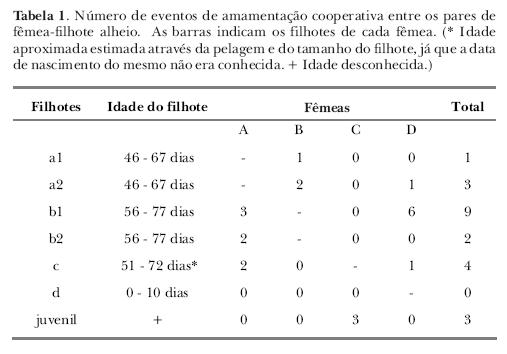
Em cerca de 41% dos eventos de AC, os filhotes alheios mamaram acompanhados do filhote da fêmea em questão; em 14%, houve parceria entre filhotes alheios. Em um destes casos, o filhote a2 mamou com o filhote b1 na fêmea D, em ocasião na qual o filhote dela, d, não estava mamando, estando apenas próximo da fêmea. Em 45%, os filhotes mamaram sozinhos nas fêmeas alheias. Embora as ACs tenham se dividido quase igualmente entre solitárias e acompanhadas do filhote da fêmea, a duração foi significativamente maior quando na presença de parceiro (m = 101,86 segundos e dp = 90,16, com parceiro; m = 10,67 e dp = 9,69, quando sozinho; t (11) = 2,45; p = 0,032).
Quando um filhote mama em sua própria mãe acompanhado de outros, a parceria pode ser entre irmãos, no caso dos filhotes das fêmeas A e B, ou com filhotes alheios, tanto no caso de ninhadas pluríparas como no de ninhadas uníparas. Nos episódios de parceria entre irmãos, a média da duração das mamadas foi de 103,68 segundos (dp = 115,28); nas mamadas de um filhote pluríparo com um alheio, a média foi de 121,50 segundos (dp = 95,01); e, nas mamadas de filhote uníparo com alheio, de 80,00 segundos (dp = 96,45). Apesar das diferenças entre esses dados não serem significativas, e o número de episódios entre pluríparos e uníparos com alheios ser pequeno, essas médias podem ser indicativas de que o tamanho da ninhada da fêmea pode ser uma influência importante na duração das mamadas com filhotes alheios. No geral, assim como na AC, a duração dos eventos é significativamente maior quando o filhote mama na presença de um parceiro do que quando sozinho (m = 103,41 segundos e dp = 113,94, com parceiro e m = 26,57 e dp = 55,87, quando sozinho; t (478) = 9,91; p < 0,001).
Quanto à posição dos filhotes em relação às fêmeas nos eventos de amamentação, observou-se que os filhotes são amamentados cooperativamente com maior freqüência por entre as patas traseiras das fêmeas (68,2% dos eventos), o que também ocorreu nos eventos de APP (88,6%). A preferência por mamar em posição perpendicular à fêmea ficou em 27,3% na AC e 9,7% na APP. A porcentagem de eventos nos quais os filhotes mamaram embaixo da fêmea foi de 4,5% na AC e de 1,7% na APP.
Verificou-se também, que na maioria dos eventos de AC o filhote alheio está deitado (54,5% versus 45,5% na posição em pé). No caso da APP, observou-se o contrário: na maioria dos eventos os filhotes mamam em pé (56,2% versus 43,8% na posição deitada). Porém, não houve diferença significativa com relação à posição dos filhotes nos eventos de AC e de APP (X2 de Pearson (1, N = 672) = 0,98; p = 0,320).
Observou-se que, nos dois tipos de amamentação, geralmente o filhote mama deitado quando está acompanhado de um parceiro de mamada (75% dos eventos de AC e 67,1%, de APP). Quando está sozinho, ele mama em pé na maioria das vezes (70% na AC e 70,9%, na APP). A duração dos eventos é maior quando o filhote está deitado (F (1, 488) = 84,62; p < 0,001) ou quando está na presença de um parceiro de mamada (F (1, 488) = 39,14; p < 0,001) (Tabela 2).

A maioria dos eventos de amamentação foi finalizada pelas fêmeas. Na AC, em 81,8% dos eventos, elas andam deixando o filhote para trás ou mexem as patas traseiras até que ele pare de mamar. Os filhotes tiveram a iniciativa de parar em apenas dois eventos, e em outros dois, a díade fêmea-filhote foi interrompida por outros indivíduos do bando. Nos eventos de APP também é a fêmea que interrompe o evento na maioria das vezes (94,5% versus 5,5% de eventos nos quais o filhote é que pára de mamar).
Em contraste com os 671 eventos de amamentação observados, foram amostrados 79 eventos nos quais as fêmeas rejeitaram a solicitação dos filhotes para mamar. Analisando-se esses eventos, observou-se que apenas seis foram solicitações de filhotes alheios. Desses casos, cinco rejeições foram de caráter agonístico (quatro eventos nos quais a fêmea D rejeitou os filhotes a2, b1, c e um juvenil; e um evento no qual a fêmea C rejeitou o filhote a1) e uma única rejeição por afastamento, em que a fêmea andou e deixou o filhote para trás (a fêmea B e o filhote a1). Os outros 73 eventos de rejeição corresponderam a solicitações de filhotes próprios das fêmeas. Dentre esses, 69 foram de rejeições não agonísticas; em dois, as fêmeas mexeram as patas traseiras para tentar impedir que os filhotes mamassem; e em outros dois houve reação agonística (envolvendo a fêmea A e seus filhotes a1 e a2). Nos eventos nos quais os filhotes foram bem sucedidos em mamar cooperativamente, as fêmeas aceitaram as solicitações dos filhotes alheios na primeira tentativa, com exceção de um evento envolvendo a fêmea D e o filhote b1. A fêmea mexia as patas traseiras para evitar que o filhote mamasse nos primeiros instantes, mas o filhote continuou tentando e a fêmea acabou aceitando que ele mamasse.
Discussão
A baixa freqüência de AC
Influência das características do grupo e da ninhada. Seguindo as hipóteses de Packer et al. (1992) com relação ao tamanho do grupo e tamanho de ninhada, esperava-se que a freqüência de AC fosse alta em catetos, já que esses animais vivem geralmente em bandos pequenos e aparentados e, as fêmeas podem dar à luz a mais de um filhote. De acordo com esta lógica, em capivaras, por exemplo, que também têm ninhadas de mais de um filhote e vivem em bandos aparentados, a freqüência de AC foi tal, que não se observou diferença significativa com a amamentação da própria ninhada (Nogueira, 1997). Embora a hipótese seja corroborada por evidências em algumas espécies, têm sido obtidos resultados diversos, mostrando que a lógica subjacente ao fenômeno é mais complexa. Em búfalos, por exemplo, também se encontrou uma certa divergência das hipóteses de Packer et al. (1992) só que de maneira contrária ao que ocorreu neste trabalho. Como búfalos são uníparos e vivem em grupos grandes, era esperada uma baixa ocorrência de AC, mas foi observada uma freqüência alta (Paranhos da Costa, Cromberg, & Andriolo, 1996).
Influência do cativeiro. Segundo Packer et al. (1992), a AC é mais comum em estudos de cativeiro do que nos estudos de campo. Muitos sugerem que o comportamento pode estar sendo aumentado nestas condições pelo acesso ilimitado ao alimento, que poderia estar diminuindo os custos da amamentação e possibilitando a amamentação de filhotes alheios. Na presente pesquisa não se confirmou a previsão desta influência do cativeiro. É possível que outras variáveis associadas ao cativeiro possam influenciar a freqüência do comportamento em sentido contrário. A formação de grupos de animais ao acaso pode interferir na estrutura social do grupo quanto ao parentesco e familiaridade. Em espécies nas quais estes fatores têm um papel importante, essa alteração poderia diminuir a freqüência da cooperação, apesar da quantidade ilimitada de alimento. A ocorrência de AC não depende só das condições nutricionais, mas também da tolerância aos filhotes alheios, que pode estar relacionada ao parentesco e à familiaridade entre as fêmeas e entre estas e os filhotes.
No caso dos catetos, a estrutura social e o parentesco entre as fêmeas são determinantes na ocorrência dos comportamentos sociais. Nogueira-Filho (1997) concluiu que fêmeas não aparentadas brigam mais e recomendou a formação de grupos aparentados para evitar a morte dos filhotes, no caso de criação em áreas pequenas. Na natureza, catetos são extremamente cooperativos e tolerantes aos infantes; contudo, no cativeiro os infanticídios podem ser freqüentes (Nogueira-Filho, 1997; Packard, Babbitt, Franchek, & Pierce, 1991). Na Fazenda Vale Verde, foi observado um caso de infanticídio em um outro grupo: uma fêmea deu à luz a dois filhotes e logo em seguida uma outra fêmea (sem filhotes) matou os recém-nascidos (Observação Pessoal). Os registros mostraram que a fêmea que cometeu o infanticídio era de procedência diferente das outras fêmeas do recinto, portanto não aparentada. A partir desta evidência, o parentesco e a familiaridade entre as fêmeas devem ser investigados como fatores associados à tolerância aos filhotes alheios e potencialmente favoráveis à AC na natureza e no cativeiro.
As características das mamadas do ponto de vista da relação fêmea/próprio filhote/filhote alheio
Amamentação de um ou mais filhotes. Observou-se um equilíbrio entre os eventos em que os filhotes foram amamentados cooperativamente com um parceiro de mamada e aqueles em que estavam sozinhos. Isso poderia contrariar a idéia de que os filhotes alheios poderiam estar mamando de maneira oportunista durante a APP. Porém, os filhotes alheios poderiam estar obtendo maior sucesso na tentativa de mamar se estivessem acompanhados de um filhote próprio, pois a duração dos eventos em que eles estavam acompanhados foi maior do que quando estavam sozinhos. Eles poderiam estar tendo garantia de que não seriam expulsos pelas fêmeas, pois os filhotes próprios estariam mamando e se elas o fizessem, interromperiam a mamada deles também.
A duração da mamada de um filhote sozinho é maior na APP do que na AC. Isso faz pensar que o filhote alheio pode ter menos sucesso quando mama sozinho do que quando mama com um parceiro que geralmente é o próprio filhote da fêmea. Observou-se uma duração maior para as mamadas com parceiro também na APP. Talvez as motivações das fêmeas se reflitam na duração da mamada, em função do número de filhotes. Pode ser que a presença de mais de um filhote mamando aumente mais a motivação da fêmea para a amamentação.
Posição de mamada. Quanto à posição do filhote com relação à fêmea, os filhotes alheios mamaram por entre as patas traseiras, na maioria dos eventos. Tendência maior de mamar por entre as patas traseiras também foi observada na APP, o que afasta raciocínios relacionados à não identificação pela fêmea no primeiro caso. Além disso, os juvenis que são maiores e poderiam ser mais facilmente reconhecidos, mamaram na perpendicular nos três eventos observados. Pode-se pensar que o filhote que mama por entre as patas traseiras da fêmea poderia ter um maior sucesso, em termos de obtenção de leite ou talvez, essa posição seja a mais confortável para o filhote.
Contexto de rejeição da fêmea às tentativas de mamada. As rejeições das fêmeas às tentativas de mamadas dos filhotes alheios foram poucas, porém a maioria foi de caráter agonístico, principalmente se comparadas às da própria prole (cinco rejeições agonísticas em seis na AC versus duas em 73 na APP). Essas rejeições foram principalmente desempenhadas pela fêmea D. Este dado não deve ser tomado em absoluto, pois esta fêmea também foi a que mais amamentou cooperativamente. A conjunção destes dados sugere que esta fêmea exerceu uma atração sobre os demais filhotes. O fato de esta fêmea ter filhote relativamente mais novo do que as demais sugere que se deva investigar a hipótese de períodos mais favoráveis para a AC em função da idade da própria prole.
As rejeições, sejam elas de caráter agonístico ou não, poderiam também estar mais ligadas a conflitos motivacionais das fêmeas, envolvendo tendências incompatíveis com a amamentação, como para andar ou forragear, versus tendências facilitadoras da amamentação, que poderiam incluir condições da própria fêmea e de um conjunto complexo de estimu-lações provenientes dos filhotes. Em um episódio entre o filhote b1 e a fêmea D, ela ficou rejeitando-o, mexendo as patas traseiras, mas o filhote ficou insistindo e acabou conseguindo mamar. Este exemplo sugere utilidade na investigação da regulação da disponibilidade da fêmea em decorrência da insistência do filhote.
As motivações das fêmeas parecem estar influenciando também na finalização dos eventos, tanto de AC, quanto de APP. As fêmeas finalizaram a maioria dos eventos e, na maioria deles, elas simplesmente andaram e deixaram os filhotes para trás, para iniciarem outras atividades, ou mesmo para acompanhar o bando, que estava se dirigindo para outro canto do recinto. Muitas vezes, os filhotes correram atrás e ficaram tentando mamar; em algumas ocasiões elas pararam e deixaram que eles mamassem novamente, ou continuaram andando sem permitir que eles mamassem (Observação Pessoal), o que sugere futuras investigações sistemáticas das características da rejeição, do tipo de aceitação, do efeito da insistência dos filhotes e do tipo de finalização dos eventos, em função da identidade dos filhotes.
Apesar da baixa freqüência de AC observada, foi possível descrever o comportamento e identificar variáveis que podem favorecer ou dificultar a ocorrência dessa amamentação nesta espécie, como por exemplo, a familiaridade e a convivência das fêmeas, o tamanho da ninhada e o período do ciclo de desenvolvimento. Além disso, pôde-se obter um melhor entendimento também da APP.
Esta exploração inicial propiciou condições para o planejamento de novos estudos que visem investigar as hipóteses causais e funcionais do comportamento de APP e AC, e também o seu reflexo no desenvolvimento dos filhotes.
Referências
Altmann, J., Altmann, S. A., & Hausfater, G. (1978). Primate infant's effects on mother's future reproduction. Science, 201, 1028-1030. [ Links ]
Byers, J. A., & Bekoff, M. (1981). Social, spacing, and cooperative behavior of the collared peccary, Tayassu tajacu. Journal of Mammalogy, 62, 767-785. [ Links ]
Clutton-Brock, T. H., Albon, S. D., & Guiness, F. E. (1989). Fitness costs of gestation and lactation in wild mammals. Nature, 337, 260-262. [ Links ]
Fogden, S. C. L. (1971). Mother-young behaviour at grey seal breeding beaches. Journal of Zoology (London), 164, 61-92. [ Links ]
Hoogland J. L., Tamarin, R. H., & Levy, C. K. (1989). Communal nursing in praire dogs. Behavior Ecology and Sociobiology, 24, 91-95. [ Links ]
McCracken, G. F. (1984). Communal nursing in Mexican free-tailed bat maternity colonies. Science, 223, 1090-1091. [ Links ]
Nogueira, S. S. da C. (1997). Manejo reprodutivo da capivara (Hydrochaeris hydrochaeris L. 1766) sob sistema intensivo de criação. Tese de Doutorado, Instituto de Psicologia, Universidade de São Paulo, São Paulo, SP. [ Links ]
Nogueira-Filho, S. L. G. (1997). A organização social de queixadas (Tayassu pecari Link 1814) e caititus (Tayassu tajacu Wetzel 1977) em cativeiro. Tese de Doutorado, Instituto de Psicologia, Universidade de São Paulo, São Paulo, SP. [ Links ]
Packard, J. M., Babbitt, K. J., Franchek, K. M., & Pierce, P. M. (1991). Sexual competition in captive collared peccaries (Tayassu tajacu). Applied Animal Behaviour Science, 29, 319-326. [ Links ]
Packer, C., Lewis, S., & Pusey, A. (1992). A comparative analysis of non-offspring nursing. Animal Behaviour, 43, 265-281. [ Links ]
Paranhos Da Costa, M. J. R., Cromberg, V. U., & Andriolo, A. (1996). O bezerro, a mãe e as outras vacas: estudando os cuidados maternais e alomaternais em ruminantes domésticos. Anais do XIV Encontro Anual de Etologia (pp. 159-171). Uberlândia, MG: Sociedade Brasileira de Etologia. [ Links ]
Reiter, J., Stinson, N. L., & Le Boeuf, B. J. (1978). Northern elephant seal development: the transition from weaning to nutricional independence. Behavioral Ecology and Sociobiology, 3, 337-367. [ Links ]
Roulin, A. (2002). Why do lactating females nurse alien offspring? A review of hypotheses and empirical evidence. Animal Behaviour, 63, 201-208. [ Links ]
Setz, E. Z. F. (1991). Métodos de quantificação de comportamento de primatas em estudos de campo. Em A. B. Rylands & A. T. Bernardes (Orgs.), A primatologia no Brasil - 3 (pp.411-435). Brasília, DF: Sociedade de Brasileira de Primatologia. [ Links ]
Tronick, E. Z., Morelli, G. A., & Winn, S. (1987). Multiple caretaking of Efe (pygmy) infants. American Anthropologist, 89, 96-106. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Cibele Biondo
Av Marechal Fiúza de Castro, 435, Bloco 1 / Apto. 22
05596-000, Butantã, São Paulo, SP
E-mail: cibelebiondo@yahoo.com.br
Recebido em 26 de março de 2004
Revisão recebida em 22 de junho de 2004
Aceito em 17 de agosto de 2004
* Este artigo é derivado da dissertação de mestrado, "A amamentação cooperativa em catetos (Tayassu tajacu)", Instituto de Psicologia da USP.
Apoio financeiro: FAPESP. Agradecemos ao Sr. Manuel Carrano pela permissão de realizar o trabalho em sua fazenda, e aos Drs. Fernando José Leite Ribeiro (IP/USP) e Eleonore Zulnara Freire Setz (IB/UNICAMP) pelas valiosas críticas e sugestões na ocasião da defesa da dissertação.
** Instituto de Psicologia, USP, Bolsa Produtividade de Pesquisa CNPq. E-mail: vsbussab@usp.br.

