










Serviços Personalizados
artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805versão On-line ISSN 2175-3636
Rev. etol. v.6 n.2 São Paulo dez. 2004
ARTIGOS
Joining a group decision by captive saffron finches (Sicalis flaveola) in competitive situations*
Participando de una decisión grupal por jilgueros (Sicalis flaveola) en situaciones competitivas
Celia María Lombardi**; and Walter Farina**
Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Universidad de Buenos Aires
ABSTRACT
Visual and auditory cues play an important role in promoting behavioral synchronization. Captive saffron finches (Sicalis flaveola, Passeriformes, Emberizidae) confronted with a steady size of food patch, fly to the ground more often and remain longer on it when bird density is higher. The observed effect, independently of being triggered by sensory cues resulting from more birds, may be a consequence of the increased competitiveness required by the situation. In this paper we have tested this hypothesis. We held birds in a steady group size and describe their decision to join a group at the ground at successive intervals, when variable food patch size was offered. We found that the smallest food patch size promoted the largest group size of birds on the ground. Analyses of the time domain showed that when the smallest food patch size was available, decisions to join the group on the ground were made at shorter time intervals. Apparently, birds are capable of adjusting their decisions according to the perceived competitiveness of the situation.
Index terms: Decision, Time series, Social facilitation, Competitive interaction, Finches, Sicalis flaveola.
RESUMEN
Tanto las señales visuales como las auditivas son importantes en elicitar la sincronización del comportamiento. Jilgueros (Sicalis flaveola, Passeriformes, Emberizidae) en cautiverio, aumentan la frecuencia de vuelos hacia una misma fuente de alimento y permanecen durante más tiempo en sus alrededores cuando la densidad de aves es mayor. Tal efecto, independientemente de haber sido provocado por más estímulos sensoriales como consecuencia de una mayor cantidad de aves, podría ser debido al incremento de la competitividad requerida por la situación, hipótesis puesta a prueba en el presente trabajo. Se mantuvo constante el tamaño del grupo de aves y se midió la decisión de unirse a un grupo, en intervalos sucesivos, variando el tamaño de la fuente de recursos. Se encontró que la menor fuente de alimento elicitó una mayor cantidad de aves a su alrededor, y que los grupos se formaban a intervalos de tiempo menores. Aparentemente, las aves fueron capaces de ajustar su decisión de acuerdo al grado de competitividad percibida.
Descritores: Decisión, Series de tiempo, Facilitación social, Interacción competitiva, Jilgueros Sicalis flaveola.
Local and stimulus enhancement are social learning mechanisms that promote behavioral synchronization. They arise when an animal is attracted to the site at which another one is present (Heyes, 1994; Krebs, 1974). Particularly in birds, visual and auditory signals play an important role in the configuration of such mechanisms (Clifton, 1979; Tolman, 1964). Lombardi and Charbuki (2002) have shown that in caged saffron finches (Sicalis flaveola, Passeriformes, Emberizidae) bird density influences the time spent on the ground, suggesting that birds fly to the ground as a function of the number of birds already present on it. However, another (though not exclusive) explanation would be that birds (matched in individual competitive ability) may be responding to the differential competitiveness required by the experimental conditions.
Holding bird density constant and modifying the food patch size, then, should help to clarify whether birds adjust the decision to join a group according to the different competitive levels of variable situations. In addition, if socially facilitated, joining a group at the food source should also be apparent through time in the form of a predictable function, regardless of food patch size. However, if food patches of different sizes are provided, different underlying probability models may account for the corresponding behavioral decisions, since time series would be expected to reflect condition variability. We undertook the present experiment with captive saffron finches to test: 1) the influence of food tray size on the number of birds on the ground when bird density is held steady and 2) the randomness of the sequences (i. e., of number of birds standing on the ground) in the time domain for the specified tray sizes.
Methods
Twelve wild caught saffron finches juveniles of unknown sex served as subjects (within the conditions of the article 22h, chp. VII, Law Nr 22421/81, Protection and Preservation of Wild Fauna). They were maintained collectively in an aviary (1.4 x 1.5 x 1.8 m), in natural light, and were fed a mixture of grains, fruits, vegetables and vitamins. Before the experiment began, birds had been in the aviary for three months. The aviary contained perches but no nesting places nor nest material. Juvenile birds were selected to minimize individual competitive ability (Marcondes-Machado, 1997). After the experiment (Spring season, 2002), birds were released at the same place where they were captured.
Experimental sessions were conducted in the same aviary where the birds were caged, from 9:30 to 12:00 h. once or twice a week. One side of the aviary allowed a free view of the surrounding countryside, while the other sides were covered with an opaque screen. A small opening in the screen allowed the observer to collect data by hand without being noticed by the subjects.
The aviary floor was covered with sheets of paper to facilitate its cleaning before each session. Two strips of wood were placed on the floor to divide it in four equal parts. One of them was selected as the "food plane" and the one beside it, as the "water plane". To facilitate data collection the latter "plane" was at 50 cm height and consisted of a cardboard on a wood stand. Both food and water "planes" were of similar size and were used for placing food and water trays.
Four tray sizes were used (50 x 25 cm, 25 x 15 cm, 15 x 10 cm and 10 x 10 cm, and were called nr 1, 2, 3 and 4, respectively), all of which were 5 cm high. Only one tray size per session was employed, and each session involved one tray filled with food and another with water. The four sizes were used according to a quasi random sequence, avoiding the same size appearing more than three times consecutively. For each tray size 17 sessions (replicates) were conducted.
One week before the start of the experimental sessions, birds were exposed for short periods to the experimental trays without formally collecting data on their behavior. Before the beginning of each session the aviary floor was cleaned and the habitual food, sand and water trays were removed. The experimental food and water planes with their corresponding trays were positioned and the collection of data began 30 min later. A session consisted of 25 consecutive scan samples conducted at 5 min intervals. At the end of a session the experimental trays were removed and replaced with the habitual ones. All sessions involved the same group of individuals. During scan samples, the number of birds standing on food and water planes were recorded. Since different tray perimeters did not allow the same number of birds simultaneously standing on them, the number of birds only at the food tray may have represented the behavior of dominant individuals.
Statistical analyses were run with BMDP (Dixon, 1992), checking for normality (W statistic) and variability (Levene F). Since data were not normally distributed they were transformed to arcsine √p and then retransformed for presentation (Damon & Harvey, 1987). Regressions were done using the mean over replicates, i.e., each tray size yielded one datum point. The difference among mean proportion of birds at tray sizes was tested using repeated measures ANOVA with Huynh-Feldt adjustment, with replicates as the repeated measures (n = 17).
Analyses of data points at each time interval were done with the Box-Jenkins approach for time series (for each tray size, n = 425). Plots were checked for trends. Tests of white noise were done with the Ljung-Box Q statistic (LBQ) simultaneously at 26 lags of the autocorrelation function (ACF), for the undifferenced series (Cook & Campbell, 1979). The lag 26 was selected to look for a possible seasonal effect at the two hours that each session lasted. Exact P values for LBQ were found with StaTable (Mehta & Patel, 1994). Approximate 95% confidence limits (± 2/√N) were plotted on the correlograms (Chatfield, 1975; Dixon, 1992).
Results
Table 1 shows the mean proportion of birds standing on food and water planes at all tray sizes. Means related to the food plane increased according to tray size, with the highest value on the smallest tray. The comparison between tray sizes 1 and 4 was highly significant. No effect was found associated to means related to the water plane. Linear regressions explained 92% of the variance for food, and 11% for water (regression F1,2 = 23.224, P = 0.04 and F1,2 = 0.244, P = 0.67 for food and water, respectively).
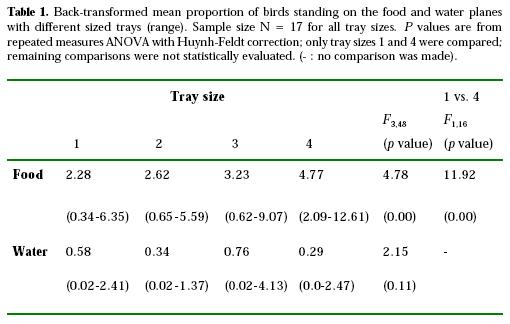
In order to test for a depletion effect on the food trays, a repeated measures ANOVA with tray size and hour (first and second hour) as within factors was carried out. No effect was found (F 1,16 = 1.11, P = 0.31).
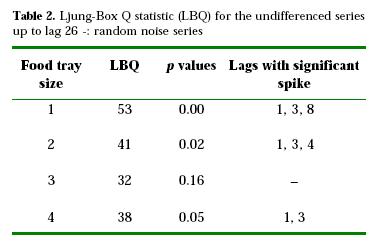
Table 2 shows the LBQ values for the ACFs of proportion of birds standing on the food plane. No seasonal effect was apparent at lag 25 (Figs. 1, 2, 3, 4). Three of the four series related to food yielded significant LBQ statistics, indicating that they were not random noise processes (food tray size 1, 2, 4; Figs. 1, 2, 4, respectively). Despite the fact that the ACF plot for food tray size 3 showed a significant spike at lag 2, its LBQ statistic indicates that the series was a random process (Table 2; Fig. 3). All sizes related to water (not shown) were identified as white noise, i.e., data points were uncorrelated with each other.
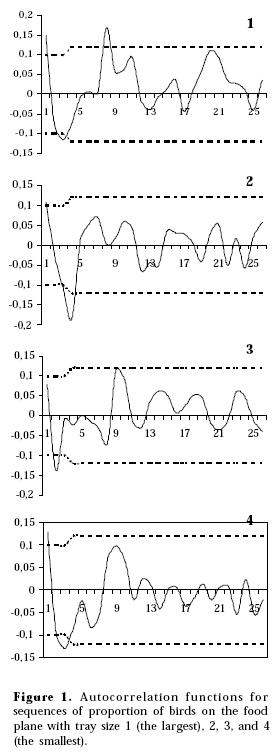
All significant sequences (food tray sizes 1, 2, 4; Figs. 1, 2, 4 respectively) showed a significant correlation at lag 1, suggesting a possible slight positive trend throughout the entire sequence, i.e. a portion of each observation in the series was predictable by the immediately preceding value. However, series were not differenced to avoid introducing spurious periodicities. Series appeared to be complex, mixed processes, hence, no attempt was made to identify mathematical models that describe them since there were no simple relationships among adjacent observations. All three significant series did not show a clear cyclic pattern (Fig. 1, 2, 4), though they did present some interesting features.
A short term correlation was present at different lags for different tray sizes, as showed by the positioning of spikes (Table 2). Significant spikes at lag 3 and 8 related with the largest tray (size 1), and at lags 3 and 4 related with the tray size 2 roughly indicate that a current outcome in the series was predictable by portions of two preceding values, with the largest tray eliciting joining on the ground decisions at greater time intervals than the remaining tray sizes. The spike at lag 8 of the largest tray sequence was positive, implying that large groups of birds on the ground tended to be followed by large groups and small groups by small ones. Only the series related to the smallest food tray (size 4) presented a somewhat clearer periodicity at lag 3, since the partial autocorrelation function (not shown) also had a significant spike at lag 6, being both negative. Therefore, birds tended to be joined on the ground, when the smallest food tray was present, at intervals of 15 min (since scans were taken at 5 min intervals) though with large values of the series followed by small values and vice versa.
Discussion
In this experiment we clearly show that birds modified their decision to join a group according to the degree of competitiveness of the situation, corroborating previous findings by Lombardi and Charbuki (2002). Holding bird density steady, birds gathered on the ground in larger groups if food was clumped in a smaller patch. The phenomenon may be similar to the well known socially facilitated feeding demonstrated in chicks (Tolman, 1964), and also called competitive interaction by Clark and Mangel (1986), since it appears that the competitiveness of the situation may trigger the specified behavior. The lack of any pattern related to the trays with water was probably due to their double function, since it is for drinking and bathing, and probably also due to lack of thirst.
Time series analyses allow the study of behavioral sequences taking in account earlier behaviors. Traditionally, they have been applied to single organisms. Since in the present study bird density was held steady, we took the approach that a group could be assumed to be a closed system. We have defined closed system as that where the only inputs come from the system itself. Accordingly, we have taken a single dependent variable as a measurement for the behavior of the whole group. Analyses in the time domain were used to identify possible cyclic oscillations in the behavior of the group. Though clear periodicity through time was not apparent in the sequences, the size of food trays did seem to control, to a certain degree, the decision of birds to join on the ground. Three of the four sequences were not random noise. However, each of these three sequences presented its own pattern, appearing to represent specific decisions according to the degree of competitiveness of the experimental condition. Current values of each series seem to be roughly predictable by different models. Apparently, there were shorter-term correlations associated with the series of the smallest food patch than those related to the largest food patch. Since for free ranging birds, greater protection against predators and more information about food are the most obvious advantages for joining a group, despite the decreased intake due to competition (Sirot, 2000), there is a benefit in non randomness. However, only a certain degree of predictability may be the most adapted behavior, as is the case for vigilance (Desportes, Metcalfe, Cezilly, Lauvergeon, & Kervella, 1989). If sequences of attraction to a food source were clearly periodic, the advantages of joining a group could diminish, especially those regarding protection against predators, as birds would be an easier prey for waiting predators.
References
Chatfield, C. (1975). The analysis of time series: Theory and practice. London: Chapman and Hall. [ Links ]
Clifton, P. G. (1979). The synchronization of feeding in domestic chicks by sound alone. Animal Behaviour, 27, 829-832. [ Links ]
Clark, C. W., & Mangel, M. (1986). The evolutionary advantages of group foraging. Theoretical Population Biology, 30, 45-75. [ Links ]
Cook, T. D., & Campbell, D. T. (1979). Quasi-experimentation: Design & analysis issues for field settings. Boston: Houghton Mifflin Company. [ Links ]
Damon, R. A., & Harvey, W. R. (1987). Experimental design, ANOVA and regression. New York: Harper & Row. [ Links ]
Desportes, J.-P., Metcalfe, N. B., Cezilly, F., Lauvergeon, G., & Kervella, A. (1989). Tests of the sequential randomness of vigilance behaviour using spectral analysis. Animal Behaviour, 38, 771-777. [ Links ]
Dixon, W. J. (1992). BMDP: Statistical software manual. Berkeley: University of California Press. [ Links ]
Heyes, C. M. (1994). Social learning in animals: Categories and mechanisms. Biological Review, 69, 207-231. [ Links ]
Krebs, J. R. (1974). Colonial nesting and social feeding as strategies for exploiting food resources in the great blue heron (Ardea herodias). Behaviour, 41, 99-134. [ Links ]
Lombardi, C. M., & Charbuki, M. (2002). Effect of bird density on the decision to join a group in the Sicalis flaveola pelzeni (Passeriformes, Emberizidae). Iheringia Série Zoologia, 92, 105-109. [ Links ]
Marcondes-Machado, L. O. (1997). Comportamento social de Sicalis flaveola brasiliensis em cativeiro (Passeriformes, Emberizidae). Iheringia Série Zoologia, 82, 151-158. [ Links ]
Mehta, C. R., & Patel, N. R. (1994). StaTable: Electronic tables for statisticians and engineers. Cambridge: Cytel Software Corporation. [ Links ]
Sirot, E. (2000). An evolutionarily stable strategy for aggressiveness in feeding groups. Behavioral Ecology, 11, 351-356. [ Links ]
Tolman, C. W. (1964). Social facilitation of feeding behaviour in the domestic chick. Animal Behaviour, 12, 245-251. [ Links ]
 Correspondence
Correspondence
Celia Lombardi
Museo Argentino de Ciencias Naturales "Bernardino Rivadavia"
Av. Gallardo 470,
C1405DJR, Buenos Aires, Argentina
E-mail: celia@mail.retina.ar
Received January 10, 2005
Revision received May 25, 2005
Accepted June 17, 2005
* We thank S. Hurlbert and C. Herrera, and two anonymous reviewers for their helpful comments on the manuscript and for improving our English. We also thank M. Cirulo, C. Kobashigawa and N. Smud for their kind help with word processing, and G. Carrizo and H. Carcacha for their advice on wild fauna management. We are greatly indebted to Rumivet S.R.L. for generously providing the bird medicaments.
** Requests for reprints should be sent to Celia Lombardi, Museo Argentino de Ciencias Naturales, "Bernardino Rivadavia".

