










Services on Demand
article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkRevista de Etologia
Print version ISSN 1517-2805On-line version ISSN 2175-3636
Rev. etol. vol.8 no.2 São Paulo Dec. 2006
ARTIGOS
Observer effects on the behavior of non-habituated wild living marmosets (Callithrix jacchus)
Efeitos do observador sobre o comportamento de Sagüis (Callithrix jacchus) não habituados de vida livre
Marcos Vinícius de AlmeidaI, *; Hugo Medeiros Garrido de PaulaII, **; Renata Souza TavoraIII, ***
I Universidade de São Paulo
II Universidade Estadual Paulista, Bauru
III Universidade de Brasília
ABSTRACT
In order to assess the influence of the presence of a human observer on the behavior of wild, non-habituated common marmosets (Callithrix jacchus), a group of marmosets was exposed to sessions of observation in which observers were visible and in which they hided into a blind. Results indicated that typical defensive reactions such as alarm calls and stares at observers were reduced when observations were made from inside the blind. There was no significant differences in other parameters such as foraging/social contact calls, minimal distance from observer and time duration of sessions. Both major and minor modifications in normal behavior of free living animals may thus arise from the conditons of observation and should be taken into account.
Keywords: Defensive behavior, Observer effect, Blind, Observational methods, Callithrix jacchus.
RESUMO
Para avaliar o impacto da presença de um observador humano sob o comportamento de sagüis silvestres (Callithrix jacchus), o comportamento de um grupo desses animais foi observado em sessões conduzidas, aleatoriamente por observadores visíveis e dentro de uma barraca camuflada. Verificou-se que as reações defensivas típicas, como chamadas de alarme e olhares fixos aos observadores, foram reduzidas no caso das observações feitas dentro da barraca, não havendo diferenças significativas em parâmetros como contato social, forrageamento, distância mínima do observador e a duração do tempo das sessões de observações. As condições de observação podem exercer modificações no comportamento de animais em estudos naturalísticos e deveriam ser levadas em conta.
Palavras-chave: Comportamento defensivo, Efeito do observador, Barraca de observação, Callithrix jacchus.
The act of observing an animal involves the inherent and inevitable risk of interference upon its normal behavior (Fentress, 1992). This disturbance is widely known as the observer effect and, due to its importance, is one topic frequently cited in handbooks on ethology (Lehner, 1996). The observer effect depends on the sensory capacities of the observed animals, their species, their previous experience with human beings, their age, gender and social status, among others (Estep & Hetts, 1992). The responses of animals towards human observers consequently show great variability, but most of them display defensive reactions as humans are discriminated as potentially dangereous. Even domestic animals such as horses, chicken and pigs that routinely maintain contact with humans show important signs of disturbance when being observed (Crowell-Davis, 1992; Duncan, 1992; Hemsworth, Barnett, & Coleman, 1992). The act of observing animals takes place not only on farms but also in zoos and safari parks as well as in laboratory or field research, and targets both domestic and wild animals. Thus the observer effect is a matter of concern for everyone interested in collecting information about animal behavior.
Research on the observer effect is relevant as it may supply support for the development of methods that reduce or control this effect. The usage of blinds or other devices which keep the observers hidden, automatic recording and habituation have been indicated as possible controlling alternatives (Lehner, 1996). However, considering the diversity of the observer effect among species and the viability of different controlling methods, there is no fixed protocol to be followed and each case deserves special attention.
The common marmoset Callithrix jacchus is a small neotropical primate with diurnal habits widely studied under naturalistic conditions. Regarding its saltatorial locomotion, the controlling method routinely used to observe this animal under field conditions is habituation, accomplished by repetitive and systematic following of the animals until they show signs of indifference towards the human observers. Using this technique, it has been possible to investigate many aspects of their ecology and natural behavior (Stevenson & Rylands, 1988). Despite habituation, the most frequent reaction that marmosets exhibit in their initial contacts with humans is a highintensity flight, suggesting that the human presence in the field has an aversive nature. Possibly, humans are perceived as predators, as happens with other neotropical primates (Galetti, 1996).
Callithrix jacchus has been used as a laboratory animal for nearly a half century (Stellar, 1960) and its behavioral repertoire has been described in detail (Stevenson & Poole, 1976) including defensive behaviors, escaping and fighting responses, as well as postures and vocalizations displayed with minimal levels of threat (Lipp, 1978). Marmosets have also been even used in etho-experimental approaches of the neuropharmacological substrates of anxiety and fear in animals (Barros & Tomaz, 2002). the objective of the present study was to investigate, using such knowledge as a basis, the observer effect in a group of non-habituated wild living Callithrix jacchus. Records when the observers were visible were compared to those taken when observers hid inside a blind.
Method
In the beginning of July 2002, the presence of marmosets was detected indirectly by means of the typical tree scars provoked by exudate feeding, and directly through sporadic visualizations, in the 136-hectare UNESP/Bauru Legal Reserve located in Bauru city (SP)/Brazil (22º20' S 49º01' W), which has native cerradão vegetation mixed with semi-deciduous forest formation. In order to lure the animals to a place where the experiments could be conducted, a feeder was installed in a clearing and supplied every two days with two bananas cut into pieces. Systematic visiting allowed us to confirm that the feeder was being used by the marmosets: the fruit was frequently consumed during the diurnal period and typical teeth marks were found. . The focal group was composed of one adult pair, three sub-adult members (one female and two males), two young and two infants with undetermined gender.
Twelve observation sessions were conducted in the area of the feeder. In six of theses sessions, two observers stayed 8 meters from the feeder dressed in camouflage clothes and maintained constant visual contact with the animals. In the remaining six sessions, the observers were hidden inside a blind. In order to avoid habituation of animals to any of the conditions, the order of trials with the blind or with visible observers was randomized and other feeder visits which served only for food supplying and in which no observation was conducted were included in the schedule. Observers arrived at the area of the feeder at 7:00h. Observation started when the presence of the animal was noted and stopped when marmosets moved to other sites. Two sessions, out of the twelve sessions scheduled, could not The Observer Effect in marmosese´s behavior be carried out (one due to unfavorable climatic conditions and the other because the animals did not appear) and had to be replaced.
The blind used was adapted from models utilized in wildlife photography. It consisted of a metal frame, 1 meter in length, 1 meter in width and 2 meters in height, Covering this metallic structure, there was a camouflage fabric with one 1-meter opening serving as the entrance and six other 10-cm openings to allow external visualization.
Observers used ad libitum observation to obtain information about contextual contitions of sessions and to record the latency for marmosets arrival at the feeder, the total time of visualization (session duration) and the minimal distance that animals stayed from the observers .All occurrences sampling (taken as relative frequency of items per 10 min intervals) was used to record the following behavioral items: (1) alarm calls: high-pitched vocalizations with short duration - less then 0.5 s - sounding like a sharp whistle (warning calls, Epple, 1968), (2) mobbing calls: tsik vocalizations emitted in bursts of 3-5 calls per second, eventually associated to crackles and coughs; (3) staring at the observers: a behavior which frequently went with conspicuous defensive patterns such as lurking, head swaying piloerection and frowning. (4) staring in other directions: generally displayed to birds or terrestrial animals; (5) phee calls: loud monosyllabic vocalizations, approximately 1 second in duration and with a stable frequency. Phee and long phee are described as social contact calls, territorial or foraging vocalizations in other studies (Epple, 1968; Lazaro-Perea, 2001); (6) other vocalizations: remaining vocalizations that could not be classified in the previous categories of vocal patterns, such as twitters, conspecific aggressive chatters and squeals of submission that animals usually emitted while eating at the feeder.
Results
The first animal that approached the feeder was frequently one member of the adult pair. Approaching was accompanied by alarm calls emitted by the animal itself or by surrounding group members. During this approach, any subtle movement of the observers, minimal sounds, or even facing the animal could evoke bursts of mobbing vocalizations. After reaching the feeder board, the animal quickly grabbed the food (one banana piece) and immediately climbed to adjacent trees until finding a leaf-abundant branch, where it took cover while eating. Other individuals then repeated this pattern, in general for a period of several minutes. The animals gradually began to spend more time at the feeder and ended consuming the food there. Sternal scent marking was frequently observed to be performed, at this occasion, on the tree trunk where the feeder was attached. In addition, alarm calls or mobbing were eventually emitted towards flying birds or terrestrial mammals (e.g. Dasyprocta agouti) that passed nearby.
As the feeder became empty, marmosets started to engage in area exploration. During this activity, some animals might approach the observers, whether hidden in the blind or not. This behavior was frequently performed by subadult marmosets and was accompanied by alarm calls emitted from the adult pair that stood quite distant from the observers. Approaching was done by jumping tree to tree at a moderate height (1.5 to 2.0 m) or walking on elevated (higher than 2 meters) branches. The approaching animal frequently presented tufts erected, pilo-erection, lurking, head swaying, evacuative behaviors (micturition or defecation), genital display, withdrawal attempts, and periodic visual contact with other members of the group. Approaching happened in the first session and occurred randomly across others. Animals sometimes grabbed the research material, especially bags or packages.
Playing behavior, during sessions, involved mainly young and infant members. It was marked by chases and fights in which animals actively engaged and did not result in serious aggressive patterns. Another behavior frequently recorded was resting on sunny branches, usually accompanied by selfscratching and autogrooming. Allogrooming was observed between the adult pair or among the sub-adults.
By the seventh observational session, the pattern of feeder exploration had changed drastically. Instead of appearing only when observers were located at a relative safe distance, monkeys started to come into the feeder while the experimenters were still filling it with food. On some occasions, approaching was so intense that the animals could eat the banana pieces still in the observers hand. This fact along with the bold behavior on some of the previously described occasions also involving approach led us to consider the possibility of rapid habituation. Thus, a parallel data analysis comparing the first and the last six sessions of the present study was carried out to assess the habituation effect, independently from experimental condition.
Results of the habituation analyses are presented as follows: in the first six sessions four observations were held with the observers visible to animals, and in two of them the blind was used. Consequently, in the last six sessions, in two sessions the observers were visible and in four they were hidden by the blind. A Chi-square test was used to search a possible imbalance in this distribution, but there was no statistical difference (X2 = 1.33; df=1; p <0.05). The latency for animals to arrive at the feeder showed great variability, ranging from 210 minutes to zero when the marmosets were already at the feeder by the session start. There was no statistically significant difference (t=1.08; df=5; p <0.05) between the latencies recorded in the first (mean + S.E.M.: 94.17 + 35.44 minutes) and in the last (61.67 + 26.85 minutes) six sessions. Session durations, in minutes, for the first and last six sessions, were 55.16 + 4.11 and 49.50 + 6.95 respectively , and also did not show a significant difference (t=0.59; df=5; p <0.05). The mean (+ S.E.M.) minimal distances kept from the observers were 3.21 + 2.88 meters in the first six sessions, and 4.01 + 3.99 meters in the last, also not differing statistically (t=-0.44; df=5; p < 0.67). Applying the same analysis to the results obtained with the all occurrences sampling, no statistical difference was detected for any of the parameters recorded (data not shown), with one exception. The frequency of stares at other directions recorded in the first six sessions (2.04 + 1.07) was significantly lower (t=-2.67; df=5; p <0.05) than that of the six last sessions (4.33 + 1.32).
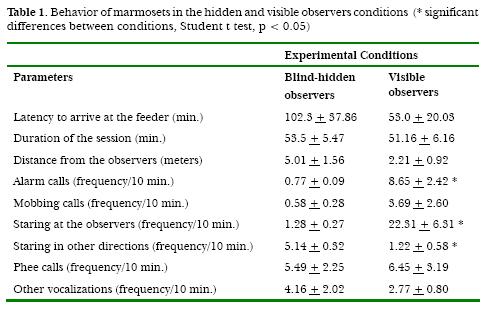
The latency of appearance at the feeder, the duration of sessions and the minimal distance from the observers did not differ significantly between conditions (observer Experimental Conditions visible or in the blind, Figure 1). There was no difference either in mobbing, phee calls and other vocalizations (all occurrences sampling). The mean frequency of the alarm calls was, however, 11-fold higher when the observers were visible to marmosets than when they were blind-hidden, a statistically significant difference (t = 3,245; p < 0.01). Stares at the observers was 17-fold higher in the visible-observers condition (t = 3.325; p < 0.001). There was a significant reduction in occurrences of staring in other directions (t = -5.858; p < 0.0001) under the observer visible conditions.
Discussion
Results indicate that an observer effect may influence in several ways the behavior of non-habituated wild living marmosets Callithrix jacchus. The use of a blind was shown to be effective in reducing some of the most conspicuous defensive reactions marmosets perform towards humans (major effects of the human presence).
Major effects are typical defensive reactions that animals exhibit when being observed or potentially threatened by human beings. In marmosets, defensive reactions are very well described (Stevenson & Poole, 1976) and part of their neural substrate located (Lipp, 1978). The ventromedial hypothalamus is involved in the production of defensive reactions in many mammals, including primates (Canteras, 2002; Hess et al., 1943; Wasmann, 1962). In Callithrix jacchus, electric stimulation of the ventromedial hypothalamus triggers conspecific vocal threat that transform into flights or even defensive fighting as the current used in stimulation is progressively increased (Lipp & Hunsperger, 1978). This fits with the observation that mammals defensive reactions are more intense when danger (type and proximity of the predator) is higher (Hendrie, Weiss, & Eilam, 1996).
In the present study, high-intensity defensive reactions, like mobbing, were systematically recorded. Mobbing was always displayed. This means that the usage of the blind did not completely eliminate the animal defensiveness. Another high-intensity defensive reaction recorded was alarm calling. Differently from the mobbing response, it was consistently reduced by blind usage. According to neurophysiological studies, alarm calls are emitted mainly during excited flight, while mobbing is more associated with the medium intensity stimulation (Lipp & Hunsperger, 1978). Mobbing calls are usually emitted by a dominant member of the group when it makes itself visible to a potentially dangerous stimulus. In contrast, alarm calls are emitted by any escaping member of the group as imminent danger is perceived and immediately induce vigorous flight in the group peers (Epple, 1968). In the present study, the animals were lured to a feeder and were thus visible to observers, a condition in which mobbing would be expected to be performed. Mobbing calls only evoked a brief increase in vigilance level, whereas alarm calls triggered flights that forced the animals to abandon the food. Mobbing behavior thus may be taken as a lower-cost response in this context, evoked by less dangerous conditions. It leads to the conclusion that the visible presence of the observer can evoke the highest defensive vocal reaction in intensity in non-habituated marmoset (the alarm calls), and that this effect can be reduced with the usage of the blind.
Vigilance is another major sign of defensiveness among animals (Hendrie et al., 1996). In marmosets, exposure to potential threats (Barros & Tomaz, 2002) and weak ventromedial hypothalamic stimulation (Lipp & Hunsperger, 1978) induce arousal and visual scanning. Caine (1998) stresses that vigilance may be considered an advantageous low-cost defense strategy among marmosets, since it slightly disrupts other essential activities, such as foraging and playing. In the present study, the blind condition was associated with reduced frequencies of stares towards the observers and raised the frequencies of staring at other stimuli. This corroborates the idea that the presence of visible observers increases the level of vigilance and promotes a deviation from usual targets of attention (Caine, 1992).
The absence of significant differences in other response parameters suggests that the presence of human observers may not interfere with aspects of the animals behavior, such as foraging and feeding, time budget in ordinary activities and area usage. Approaching behavior, frequently seen in our sessions, might be taken as reinforcing such a conclusion but it must be noted that etho-experimental studies conducted with laboratory animals point out the association between anxiety states and risk assessment activities, involving cautious investigation of a given stimulus perceived as anxiogenic by the animal (Blanchard, Yudko, Rodgers, & Blanchard, 1993). In marmosets, approaching develops into active visual exploration associated with clear manifestations of conflict and defensiveness (piloerection, attempts to escape, etc.). In marmosets, risk assessment is reproducible experimentally through the human threat test, in which one observer plays the role of the aversive stimulus. In this paradigm, approaching and visual scanning are often reported, but they are referred to as jumping towards the cage front and spending time in the cage front, respectively. It has been reported that presence of a human observer near the cage front increases 16-39% the number of jumps towards this part of the cage (Carey, Costall, Domeney, & Jones, 1992). In addition, the time spent in the cage front is systematically increased by anxiolytic agents like diazepam and buspirone (Costall, Domeney, Gerrard, Kelly, & Naylor, 1988), and decreased by anxiogenic drugs (Carey et al., 1992) revealing that staying near a human being may be anxiogenic for marmosets. The approaching behavior observed in the present study may be a risk assessment activity and may be motivated by moderate levels of anxiety.
That blind usage did not completely eliminate defensiveness indicates that the blind itself may be, as a novel stimulus, a moderate anxiogenic stimulus to marmosets (Smith, McGreer-Whitworth, & French, 1998). Distance kept from the observers, latency to arrive at the feeder and session duration, in the blind condition, reinforce such an idea. Such parameters may be interpreted as minor aspects of impacts, in contrast with the major effects, and may signal moderate levels of anxiety. Our results reinforce the results of Caines study (1992) in which it was shown that minor effects of human observation are more persistent. Minor effects may eventually be ignored or misinterpreted by observers as indicating habituation.
Our results may not be interpreted in terms of habituation, as no habituation effects were detected throughout the study. Staring at other stimuli was the only parameter which differed from the first to the second half of the observation period. We think it is not sufficient as an indication of habituation. It is possible (but further study is needed) that it reflects an indirect effect, through the habituation of other animals to the presence of marmosets: birds, for instance, seemed to be more active as sessions proceeded.
Observer effects may, in conclusion, have major and minor impacts upon behavior of nonhabituated wild living Callithrix jacchus. The former are reduced by the usage of a blind, the latter are more resistant to control and deserve attention. Our results should not be taken as invalidating existing reports of field work with marmosets: in most of such work, the process of habituation seems to have been adequately conducted and the presence of observers did not deter, apparently, the full performance of interactions in the groups (Lazaro-Perea, 2001). The contribution of the present research is to call attention to a problem that is seldom discussed in current publications on primate behavior and to indicate the relevance of considering the levels of defensiveness of animals in the elaboration of habituation criteria.
References
Barros, M., & Tomaz, C. (2002). Non-human primate models for investigating fear and anxiety. Neuroscience and Biobehavioral Reviews, 28, 187-201. [ Links ]
Blanchard, R. J., Yudko, E. B., Rodgers, R. J., & Blanchard, D. C. (1993). Defense system psychopharmacology: An ethological approach to the pharmacology of fear and anxiety. Behavioural and Brain Research, 58, 155-165. [ Links ]
Caine, N. G. (1992). Humans as predators: Observational studies and the risk of pseudohabituation. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 357-364). New York: Cambridge University Press. [ Links ]
Caine, N. G. (1998). Cutting costs in response to predatory threat by Geoffroys marmosets (Callithrix geoffroyi). American Journal of Primatology, 46, 187-196. [ Links ]
Canteras, N. S. (2002). The medial hypothalamic defensive system: Hodological organization and functional implications. Pharmacology Biochemistry and Behavior, 71, 481-491. [ Links ]
Carey G. J., Costall, B., Domeney, A. M., & Jones, D. N. C. (1992). Behavioural effects of anxiogenic agents in the common marmoset. Pharmacology Biochemistry and Behavior, 42, 143-153. [ Links ]
Costall, B., Domeney, A. M., Gerrard, P. A., Kelly, M. E., & Naylor, R. J. (1988). Zacopride: Anxiolytic profile in rodent and primate models of anxiety. Journal of Pharmacy and Pharmacology, 40, 302-305. [ Links ]
Crowell-Davis, S. L. (1992). The effect of the researcher on the behavior of horses. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 365-382). New York: Cambridge University Press. [ Links ]
Duncan, I. J. H. (1992). The effect of the researcher on the behavior of the poultry. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 285-294). New York: Cambridge University Press. [ Links ]
Epple, G. (1968). Comparative studies on vocalization in marmoset monkeys (Hapalidae). Folia Primatologica, 8, 1-40. [ Links ]
Estep, D. Q., & Hetts, S. (1992). Interactions, relationships, and bonds: The conceptual basis for scientist-animal relations. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 6-26). New York: Cambridge University Press. [ Links ]
Fentress, J. C. (1992). The covalent animal: On bonds and their boundaries in behavioral research. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 44-71). New York: Cambridge University Press. [ Links ]
Galetti, M. (1996). Comportamentos antipredatórios de quatro espécies de primatas do sudeste do Brasil. Revista Brasileira de Biologia, 56, 203-209. [ Links ]
Hemsworth, P. H., Barnett, J. L., & Coleman, G. J. (1992). Fear of humans and its consequences for the domestic pig. In H. Davis & D. Balfour (Eds.), The inevitable bond: Examining scientist-animal interactions (pp. 264-284). New York: Cambridge University Press. [ Links ]
Hendrie, C. A., Weiss, S. M., & Eilam, D. (1996). Exploration and predation models of anxiety: Evidence from laboratory and wild species. Pharmacology Biochemistry and Behavior, 54, 13-20. [ Links ]
Lazaro-Perea, C. (2001). Intergroup interaction in wild common marmosets Callithrix jacchus: Territorial defence and assessment of neighbours. Animal Behavior, 62, 11-21. [ Links ]
Lehner, P. N. (1996). Handbook of ethological methods. New York: Cambridge University Press. [ Links ]
Lipp, H. P. (1978). Aggression and flight behaviour of the marmoset monkey Callithrix jacchus: An ethogram for brain stimulation studies. Brain, Behavior and Evolution, 15, 241-259. [ Links ]
Lipp, H. P., & Hunsperger, R. W. (1978). Threat, attack and flight elicited by electrical stimulation of the ventromedial hypothalamus of the marmoset monkey Callithrix jacchus. Brain, Behavior and Evolution, 15, 260-293. [ Links ]
Smith, T. E., McGreer-Whitworth, B., & French, J. A. (1998). Close proximity of the heterosexual partner reduces the physiological and behavioral consequences of novel-cage housing in black tufted-ear marmosets (Callithrix kuhli). Hormones and Behavior, 34, 211-222. [ Links ]
Stellar, E. (1960). The marmoset as a laboratory animal: Maintenance, general observations of behavior and simple learning. Journal of Comparative and Physiological Psychology, 53, 1-10. [ Links ]
Stevenson, M. F., & Poole, T. B. (1976). An ethogram of the common marmoset (Callithrix jacchus jacchus): General behavioural repertoire. Animal Behavior, 24, 428-451. [ Links ]
Stevenson, M. F., & Rylands, A. B. (1988). The marmosets genus Callithrix. In A. F. Coimbra-Filho (Ed.), Ecology and behavior of neotropical primates (Vol. 2, pp.131-231). Contagem, MG: Littera Maciel. [ Links ]
 Endereço para correspondência
Endereço para correspondência
Hugo Medeiros Garrido de Paula
Campus de Bauru
Av. Eng. Luiz Edmundo C. Coube, s/n
17033-360 Bauru, SP, Brazil
Recebido em 16 de março de 2007
Aceito em 17 de outubro de 2007
* Universidade de São Paulo, CEBTEC, ESALQ, Piracicaba. E-mail: viunesp@hotmail.com.
** Departamento de Ciências Biológicas, Faculdade de Ciências, Universidade Estadual Paulista (UNESP).
*** Centro de Desenvolvimento Sustentável da Universidade de Brasília(CDS-UnB). E-mail: retavora@yahoo.com.br
We would like to thank Ms. Maria Palmira Zagato de Almeida and Mr. Orandi de Almeida for their essential help with the production of the blind used in this study

