










Serviços Personalizados
Journal

artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805
Rev. etol. vol.9 no.1 São Paulo jan. 2010
Artigos
Comportamiento maternal de Arctocephalus Australis, en Isla de Lobos - Uruguay*
Maternal behavior of Arctocephalus Australis, in Isla de Lobos - Uruguay
Valentina Franco TrecuI; Bettina TassinoI; Enrique PáezII
I Proyecto Pinnípedos. Sección Etología, Facultad de Ciencias, Universidad de la República. Uruguay
II Departamento de Biología Pesquera, Dirección Nacional de Recursos Acuáticos, Ministerios de Ganadería Agricultura y Pesca. Uruguay
Dirección para correspondencia
RESUMEN
Comportamiento maternal de Arctocephalus australis, En los mamíferos, generalmente las hembras realizan el cuidado de la progenie repartiendo su inversión entre las crías presentes y futuras, maximizando la relación costo-beneficio. La variación en los comportamientos del cuidado maternal puede afectar en forma diferencial la supervivencia de las crías. A. australis se presenta como un interesante modelo para evaluar la incidencia de los diferentes componentes del cuidado maternal en la supervivencia del cachorro e investigar la ocurrencia de variación mensual y diaria de los comportamientos madre–cría. Diariamente se registró la presencia de hembras marcadas en la colonia mediante muestreos focales y de barrido. La frecuencia de los estados y eventos comportamentales variaron entre los meses de muestreo. Los viajes de alimentación de las madres exhibieron una duración muy variable, y el patrón de viajes-asistencia así como la permanencia de la madre con su cría incidieron en la supervivencia de los cachorros. Los resultados se discuten enmarcados en diferentes hipótesis.
Palavras-chave: Arctocephalus australis, comportamiento maternal, Uruguay, variación temporal.
ABSTRACT
In mammals, paternal care is usually performed by females, dividing their investment between present and future offspring and maximizing the cost-benefit relationship. Variation in maternal care behaviour can affect pups survival differentially. A. australis is an interest model to assess the incidence of different components of maternal care behaviour on pups survival, as well as the monthly and daily variation in mother-calf behaviour. The presence of marked females at the rookery was recorded using focal and scanning sampling methods. State and event frequencies changed between months. Feeding trips had highly variable duration and the survival was strongly affected by the foraging cycle strategy used and the mother-pup contact recorded. The results are discussed in the frame of different hypotheses.
Keywords: Arctocephalus australis, maternal behaviour, Uruguay, temporal variation.
Introducción
Los individuos a lo largo de su vida deben asignar la energía obtenida entre su crecimiento, mantenimiento y reproducción buscando maximizar su eficacia darwiniana (Begon, Harper & Townsend, 1999; Clutton Brock, 1991; Gomendio, 1994a; Krebs & Davies, 1987). La selección natural y las restricciones filogenéticas modelan en cada especie la estrategia del ciclo de vida (Wilson, 1980). La estrategia K, frecuente entre los mamíferos, conlleva la evolución de cuidado parental con alta inversión por cría, con costos para los progenitores tales como aumento de su mortalidad, disminución de su longevidad y de la producción de crías futuras (Carranza, 1994). Los progenitores invierten en su descendencia con el fin de maximizar el éxito reproductivo, obteniendo el mayor número posible de descendientes viables a lo largo de su vida (Clutton-Brock, 1991; Trivers, 1972; 1985).
La anisogamia, la fecundación interna y el sistema de apareamiento poligínico, presente en la mayoría de los mamíferos, habilitan la deserción del macho del cuidado de las crías, debido a la incertidumbre respecto a la paternidad y a la pérdida de potenciales parejas futuras (Clutton-Brock, 1991; Krebs & Davies, 1987). Por lo tanto el cuidado de la progenie es exclusivamente materno, debiendo las hembras repartir su inversión entre la cría presente y las potenciales crías futuras. Dado que el amamantamiento es el componente principal del cuidado maternal post natal, tanto en mamíferos sociales como solitarios, la unidad nuclear es la díada madre-cría, con un vínculo fuerte y prolongado (Clutton-Brock, 1991; Trivers, 1972).
Cuando el costo de invertir en crías de ambos sexos es el mismo, la proporción sexual de la población debería ser 1:1. En cambio, si un sexo requiere mayor inversión parental, la proporción sexual resultante favorecería al sexo menos costoso (Fisher, 1930). Sin embargo, en especies en que los machos exhiben una elevada varianza en el éxito reproductivo relacionado directamente con su tamaño corporal o fortaleza, el modelo de Trivers y Willard (1973) predice que las hembras en buena condición física producirán machos, mientras que las de pobre condición producirán hembras. Esta relación entre la condición materna y el sexo de la cría, determina el éxito reproductivo potencial de las crías, y por lo tanto el de sus progenitores. En este sentido, los pinnípedos son un excelente modelo para probar la teoría de inversión diferencial entre sexos ya que poseen marcado dimorfismo sexual y sistema de apareamiento poligínico (Trillmich, 1996).
Dentro de este grupo existen diferentes estrategias de cuidado maternal. En la familia Otaridae, las hembras alternan períodos de asistencia en tierra durante los cuales amamantan a sus crías, con períodos de alimentación en el mar. Diferentes factores puede afectar la duración de estos últimos (Trillmich, 1990; Gentry, 1998) como ser la distancia a las áreas de alimentación, la condición física de la hembra y el sexo del cachorro entre otros. Dicha estrategia es conocida como foraging cycle strategy (Boyd, 1998; Gentry, 1998; Gentry & Kooyman, 1986; Trillmich, 1990) y se mantiene entre 4 meses y 2 años dependiendo de la especie (Bonnes & Bowen, 1996; Trillmich, 1996). Durante el período perinatal (tiempo transcurrido entre el nacimiento de la cría y el primer viaje de alimentación de la madre) y las posteriores asistencias, las crías necesitan acumular grandes cantidades de grasa para soportar el ayuno en ausencia de sus madres (García-Aguilar & Aurioles-Gamboa, 2003; Trillmich, 1990).
El otárido Arctocephalus australis (Zimmermann, 1783) o lobo fino sudamericano, se distribuye en América del Sur a lo largo de las costas de los océanos Atlántico y Pacífico, desde Brasil (Estado de San Pablo) hasta Perú (Sur de Lima) (Vaz Ferreira, 1976). Uruguay (35ºS 56ºO) alberga la mayor colonia reproductiva de dicha especie, estimada en 400.000 individuos (Páez, 2006).
Los machos de A. australis acceden a las cópulas mediante la defensa agresiva de territorios, determinando un sistema de apareamiento poligínico. Tanto el período de gestación como el de cuidado de la cría poseen una duración aproximada de 11 meses, produciéndose el solapamiento temporal de ambas inversiones (Ponce de León, 1983; Vaz Ferreira, Vallejo, Achaval, Melgarejo & Meneghel, 1981a; Vaz Ferreira, Vallejo, Achaval, Melgarejo & Meneghel, 1981b). Durante los viajes de alimentación de las hembras, los cachorros permanecen en ayuno, por lo que la capacidad de las madres de reconocer a su cría es un componente esencial para la supervivencia del cachorro (Phillips, 2003). Se desconoce aun cuáles son los comportamientos involucrados en la inversión maternal y cómo varían a lo largo del tiempo.
Nuestra hipótesis de trabajo propone que la duración de los componentes del patrón de asistencias afecta la sobrevivencia de las crías. Por un lado, las hembras con un período perinatal de mayor duración favorecerán la sobrevivencia de sus crías ya que éstas generarán mayor cantidad de reservas. Viajes de alimentación más largos incidirán de forma negativa en la sobrevivencia ya que implica mayores períodos de ayuno. En relación a la variación de los comportamientos en el tiempo se espera que la frecuencia de amamantamiento sea mayor al inicio de la lactancia, debido a que los cachorros deben generar reservas para enfrentar el ayuno durante la primera ausencia materna. El primer objetivo fue evaluar cómo afectan a la supervivencia del cachorro los diferentes componentes del patrón de asistencias, así como los comportamientos realizados en estos períodos. El segundo fue evaluar la variación mensual y diaria de los comportamientos madre – cría en la colonia reproductiva de A. australis.
Materiales y métodos
Los muestreos se realizaron en la Isla de Lobos (35°01S, 54°50W) (Figura 1) en la zona El Muelle ubicada en la porción NE de la Isla. El período de observación se extendió desde el 1° de diciembre hasta el 24 de febrero, abarcando el período reproductivo 2004/2005. Las observaciones se realizaron a una distancia entre 1 y 15 m de los individuos, sin provocar disturbios aparentes en el área. Los horarios de observación se establecieron de acuerdo al momento del día en que el número de hembras en el área fue mayor: de 7 a 10 y de 16 a 19 horas.
Figura 1. Fotografía satelital de la Isla de Lobos (35°01S, 54°50W). Se indica la zona de estudio, ubicada al NE de la Isla y conocida como El Muelle.
El 1° de diciembre, 8 hembras elegidas al azar de la zona de estudio fueron capturadas, medidas y marcadas con caravanas Allflex Nº4 (según la Escala Holandesa, Severidad Baja). Se incluyeron además dos hembras ya marcadas en el año de su nacimiento (1998), siendo las únicas hembras de edad conocida. El 18 de diciembre se capturaron, sexaron y marcaron con cortes de pelo las crías de tres hembras marcadas.
Diariamente a las 7, 9, 16 y 18 hs se realizaron muestreos de barrido (Lehner, 1996) en 20 hembras elegidas al azar (Scan20). En cada horario de observación se efectuaron además entre 6 y 10 muestreos focales (Lehner, 1996) (duración de 15 min, intervalos de 30 s), de hembras con cría elegidas al azar y de las hembras marcadas presentes en el área de observación. En función de la presencia o ausencia de las hembras marcadas en la colonia se obtuvo el patrón de asistencia a las crías. Se consideró una hembra en viaje de alimentación cuando no se la observó en la colonia por tres períodos de observación consecutivos.
Cuidado maternal y supervivencia del cachorro
A partir de los muestreos focales realizados durante el período perinatal en las díadas marcadas, se calculó la frecuencia relativa de permanencia de la hembra con su cría y del descanso (hembra registrada sola). Las diferencias en la frecuencia de estos dos comportamientos entre las hembras tomadas de a pares, se evaluaron por medio de la prueba Chi-cuadrado con tablas de 2x2 (Sokal & Rohlf, 1979). En este caso, una hembra (#10) fue descartada del análisis, debido a que su ubicación en la colonia no permitió realizar los muestreos focales desde el punto de observación.
Se evaluaron los siguientes componentes del cuidado maternal: duración del período perinatal, patrón de viajes de alimentación-asistencias en tierra y frecuencia de los comportamientos realizados entre la díada madre-cría. La duración del período perinatal se relacionó con la del primer viaje de alimentación.
Se consideró la muerte de crías marcadas cuando no fueron observadas por tres días, o cuando el cachorro fue encontrado muerto. Las crías no marcadas fueron consideradas muertas cuando la hembra volvía de un viaje de alimentación y permanecía durante dos días sin contacto con una cría.
Variación mensual y diaria de los comportamientos madre - cría
Para determinar la variación mensual de los comportamientos madre-cría, se compararon los muestreos focales y los Scan20 correspondientes a diciembre, enero y febrero. Para estimar la variación diaria, se compararon los Scan20 correspondientes a las 7, 9, 16 y 18 hs. En los comportamientos registrados mediante muestreo focal, las unidades comportamentales se diferenciaron en estados y eventos (Martín & Bateson, 1991). Se registró la duración de cada estado, multiplicando sus frecuencias absolutas por la duración de cada intervalo de observación (30 s). Por otro lado, se registró la frecuencia de los comportamientos considerados eventos, vocalizaciones de la hembra y del cachorro, olfateo, búsqueda de los pezones para mamar, retención de la cría por parte de su madre, vocalizaciones de alarma (frente a machos, hembras o cachorros), contacto naso-nasal y agresión hacia o entre cachorros. Para determinar la variación de los comportamientos entre los diferentes meses se evaluaron por separado la frecuencia relativa de los estados y de los eventos, por medio de una prueba G replicada de Bondad de Ajuste (Sokal & Rohlf, 1979).
Debido a que el muestreo de barrido registra solamente ocurrencia, todos los comportamientos fueron tomados como eventos y por lo tanto se analizaron en forma conjunta. Fueron consideradas las siguientes categorías: amamantamiento, contacto (pasivo y activo), proximidad (pasiva y activa), vocalizaciones de la hembra y descanso. Para evaluar la variación entre los meses y horarios de muestreo de los diferentes comportamientos, se utilizó la prueba estadística no-paramétrica Kruskal-Wallis (Sokal & Rohlf, 1979). Para evaluar las diferencias entre meses consecutivos se utilizó la prueba a posteriori U Man-Whitney.
Resultados
Cuidado maternal y supervivencia del cachorro
De las 10 hembras marcadas, 9 parieron un cachorro entre el 26 de noviembre y el 10 de diciembre. Las tres crías capturadas fueron machos. La duración del período perinatal fue entre 10 y 19 días (X=13,1; DE=2,85). El primer viaje de alimentación tuvo un rango entre dos y 28 días (X=13,3; DE=10,4), período en el que murieron cinco crías (Figura 2). La primer asistencia en tierra varió entre dos y tres días (X=2,75; DE=0,5), mientras el segundo viaje duró entre tres y 19 días (X=11,5; DE=7,33; n=4). En todos los casos en que la duración del primer viaje de alimentación fue mayor a la del período perinatal, las crías murieron (Figura 2).
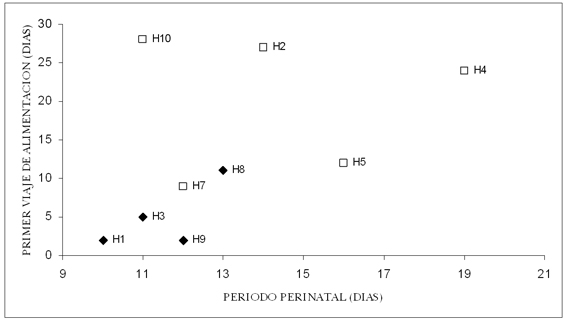
Figura 2. Relación entre la duración del período perinatal y de la duración del primer viaje de alimentación. En cada punto se indica el número de la hembra a la que corresponde (H#). Los rombos negros corresponden a las hembras cuyas crías sobrevivieron al primer viaje de alimentación. Los cuadrados blancos a las hembras que se les murió su cría durante el primer viaje de alimentación.
Considerando la permanencia y el descanso en cada una de las hembras (Figura 3), las frecuencias relativas de ambas categorías fue similar en las hembras # 2, 4 y 7 (hembra2-4 X=3,01; p=0,08; hembra4-7 X=3,01; p=0,09; hembra7-2 X=0,00; p=1), utilizando la mayor parte de su tiempo en descanso. En cambio, las hembras # 5 y 8 utilizaron la mayor parte de su tiempo en contacto con su cría (X=0,24; p=0,627). Las hembras # 1 y 9 destinaron aproximadamente la mitad de su tiempo en descanso y la otra mitad en contacto con su cría (X=1,29; p=0,256). La hembra # 3 permaneció la totalidad del tiempo con su cría (Figura 3). La frecuencia relativa de permanencia en el período perinatal osciló entre 16 - 100%.
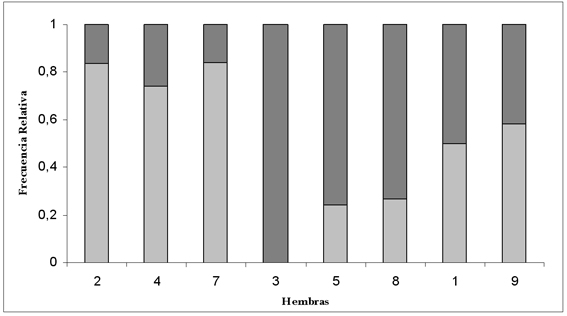
Figura 3. Frecuencia de contacto (gris oscuro) y frecuencia de descanso (gris claro) durante el periodo perinatal para cada díada para las hembras marcadas indicadas en el eje X.
Variación mensual y diaria de los comportamientos madre - cría
Los distintos estados comportamentales difirieron en su frecuencia de aparición en los tres meses considerados (dic-ene: G=37,72, p<0,001; ene-feb: G=20,32, p<0,001). De diciembre a enero aumentó el amamantamiento (G=11,57; p<0,001) y disminuyó la proximidad pasiva (G=17,98; p<0,001). Entre enero y febrero el contacto pasivo se incrementó (G=6,78; p=0,006) y la proximidad activa disminuyó (G=7,83; p=0,009) (Figura 4), siendo los únicos comportamientos que mostraron diferencias significativas entre los períodos indicados. Los eventos también exhibieron variación entre los tres meses (dic-ene: G=16,99, p=0,016; ene-feb: G=25,72, p<0,001) (Figura 5). Las alarmas con machos disminuyeron notoriamente de diciembre a enero (G=4,68; p=0,043), siendo el único evento que presentó una variación significativa entre ambos meses. En cambio entre enero y febrero las vocalizaciones de las hembras disminuyeron (G=10,92; p<0,001), mientras las alarmas hacia cachorros aumentaron (G=6,30; p=0,004) (Figura 5).
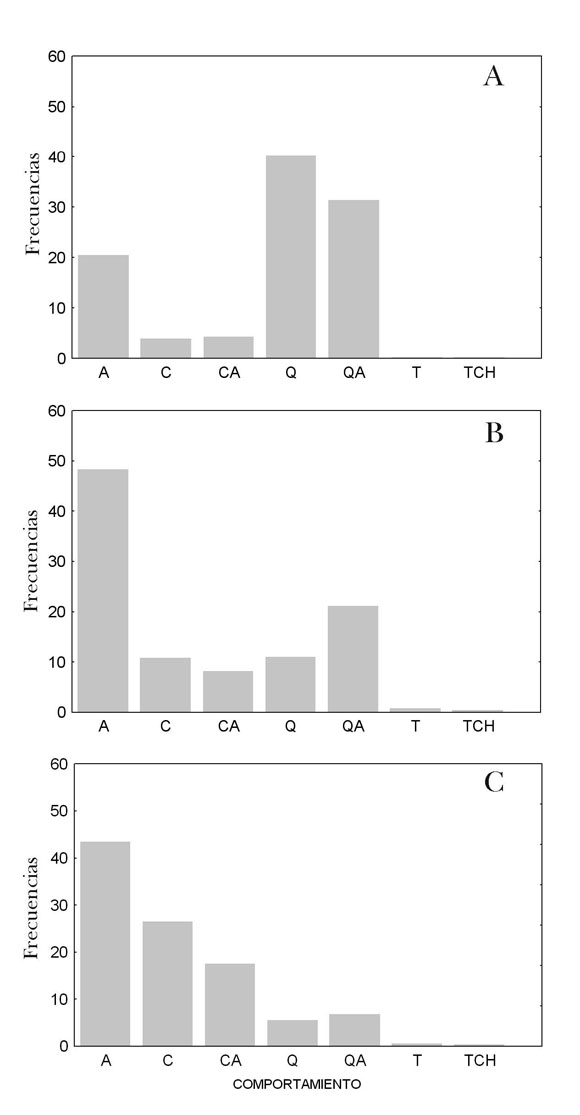
Figura 4. Frecuencias de los distintos estados eventos en diciembre (a), enero (b) y febrero (c). (A: amamantamiento, C: contacto pasivo, CA: contacto activo, Q: proximidad pasiva, QA: proximidad activa, T: traslado de la hembra, TCH: traslado del cachorro).

Figura 5. Frecuencias de los distintos eventos en diciembre (a), enero (b) y febrero (c). (OLF: olfateo de la hembra hacia su cría, VOC: vocalización de la hembra, VOCH: vocalización de la cría, B: búsqueda de los pezones en el vientre, ALM: alarmas hembra-macho, ALH: alarmas hembra-hembra, NN: contacto naso-nasal, AG: agresión hembra-cachorros, AGCH: agresión entre cachorros, ALCH: alarmas hembras-crías, R: retención de la hembra a su cachorro).
Las frecuencias obtenidas a partir del muestreo de barrido mostraron variaciones en la mayoría de los comportamientos a lo largo del período de muestreo, con excepción de las vocalizaciones de las hembras y el descanso. La frecuencia de amamantamiento (H=50,85; p<0,001; n=134), de contacto pasivo (H=19,97; p<0,001; n=113), de contacto activo (H=11,21; p=0,0037; n=50), de proximidad pasiva (H=39,80; p<0,001; n=124) y de proximidad activa (H=33,36; p<0,001; n=122) difirieron entre los tres meses. En cambio las vocalizaciones de las hembras y el descanso no mostraron variación. Entre diciembre y enero se registraron variaciones en la frecuencia de amamantamiento (U=275; p<0,001; n=91 / U=853; p=0,005; n=102) y proximidad pasiva (U=785; p=0,009; n=97 / U=286; p<0,001; n=84), mientras que entre enero y febrero también varió el contacto pasivo (U=490; p<0,001; n=88), activo (U=71; p=0,007; n=46) y proximidad activa (U=273; p<0,001; n=83). El amamantamiento fue el único comportamiento que fluctuó en los diferentes horarios (H=15,08; p=0,002; n=134), siendo su mayor frecuencia a la hora 7.
Discusión
Se constató un aumento en la duración de los viajes de alimentación de las hembras respecto a los registros previos (Ponce de León, 1983), con consecuencias negativas para la supervivencia de las crías. Por otro lado, durante el período perinatal el tiempo de permanencia de la madre con su cría fue ampliamente variable dentro de la muestra y la frecuencia de amamantamiento fue la más baja en comparación con los meses siguientes, contrariamente a la hipótesis propuesta.
Cuidado maternal y supervivencia del cachorro
Mientras las hembras permanecieron en tierra sus crías sobrevivieron. Durante el período perinatal se detectaron diferentes respuestas maternales en relación a las frecuencias de contacto y descanso. En algunas hembras predominó el tiempo de contacto con su cría mientras en otras el tiempo en descanso. Esta diferencia debería influir en el éxito de crianza ya que la permanencia de la hembra con su cría la protege de las agresiones de otros individuos de la colonia (Phillips & Stirling, 2000; Vila & Cassini, 1990) y aumenta la probabilidad de que amamante y genere las reservas necesarias para enfrentar la ausencia materna (García-Aguilar & Aurioles-Gamboa, 2003). A pesar de que no se detectó una relación clara entre la duración del período perinatal y del primer viaje, se observó que siempre que la duración del primer viaje excedió la del período perinatal las crías murieron. Cuando el primer viaje de alimentación fue breve (entre dos y cinco días) las crías sobrevivieron.
Los antecedentes para la población uruguaya indican que las hembras demoran entre 3 y 15 días en sus viajes de alimentación, aumentado la duración con la edad de las crías (Ponce de León, 1983). A pesar de que se desconoce el tamaño de la muestra y la metodología utilizada en el estudio mencionado, los resultados aquí reportados indican que la duración de los viajes de alimentación es mayor. Esto puede deberse a un cambio en el comportamiento de forrajeo de las hembras relacionado con diversos motivos. En primer lugar, las condiciones ambientales determinan la estrategia de las hembras, provocando que la duración de los viajes de alimentación varíe de acuerdo a la disponibilidad de recursos (Zalophus californianus, García-Aguilar & Aurioles-Gamboa, 2003; Arctocephalus galapagoensis y A. gazella, Trillmich, 1990; Otaria flavescens, Soto, Trites & Arias-Schreiber, 2004). En A. gazella diferentes componentes del cuidado maternal, como la duración de los viajes de alimentación, la tasa de masa transferida y el éxito de destete se relacionan con las fluctuaciones en la disponibilidad de alimento (Lunn, Boyd & Croxall, 1994). En Perú, para el lobo fino sudamericano se observa que la disponibilidad de alimento y la tasa de crecimiento de las crías se correlacionan negativamente con la temperatura superficial del mar (Trillmich, 1990). La mayor duración de los viajes de alimentación de las hembras podría relacionarse con una disminución de recursos en el ambiente o con acceso a recursos de menor calidad energética. En segundo lugar, la condición corporal y la edad de las hembras pueden afectar la duración de los viajes. Cuando las condiciones ambientales son adversas o la condición física de la madre no es buena, la elevada inversión puede acarrear costos asociados a su reproducción futura. Además, el principio de asignación (Cody, 1966) establece que los costos reproductivos son mayores para las hembras jóvenes que deben repartir la energía obtenida entre su propio crecimiento y el de su cría. Por otro lado, el aumento en la edad de las hembras disminuye la posibilidad de volver a reproducirse, por lo que las hembras más viejas invertirán más en las crías presentes, que hembras jóvenes (Gomendio, 1994b). Sin embargo, si la experiencia en el comportamiento de forrajeo se relaciona con la edad de las hembras, las hembras mayores pueden obtener el alimento necesario en menor tiempo y retornar a la colonia en un período más breve. Las hembras mayores con más experiencia de A. gazella poseen mejor desempeño reproductivo que las jóvenes e inexperientes, con mayores tasas de natalidad, mayor éxito de destete y mayor probabilidad de tener una cría en la siguiente temporada (Lunn et al., 1994). En tercer lugar, considerando la teoría de la inversión sexual (Trivers & Willard, 1973) que propone que en especies poligínicas con dimorfismo sexual, las crías macho representan mayor costo energético para sus madres, es posible que las hembras con crías macho (los tres casos en nuestro estudio) necesiten acceder a recursos con mayor valor energético, teniendo que aumentar el tiempo de búsqueda y por lo tanto la duración de los viajes. Los estudios realizados sobre inversión diferencial entre sexos en otras especies de otáridos muestran resultados muy variados. Mientras en algunas especies las madres invierten más energía y nutrientes en las crías de sexo masculino (Costa & Gentry, 1986) en otros casos no se ha encontrado una relación entre la inversión de las hembras y el sexo de la cría (Arnould, Boyd & Socha, 1996; Boltnev, York & Antonelis, 1998; Trillmich, 1986). Otros autores plantean que el gasto maternal diferencial no brinda a las crías machos un beneficio adicional ya que se debería tener en cuenta la tasa metabólica diferencial (Lunn & Arnould, 1997; Guinet, Goldsworthy & Robinson, 1999). Las madres de A. forsteri con crías macho realizan viajes de alimentación más largos que aquellas con crías hembras, indicando el mayor costo que implica (Goldsworthy, 2006). Finalmente, es posible que el mayor tamaño poblacional y crecimiento constante de A. australis, promueva la búsqueda de nuevas áreas de alimentación y nuevas presas en las hembras reproductoras de esta especie. Esto generaría una ampliación de su espectro trófico, diversificando las estrategias desarrolladas por las hembras, influyendo en la duración del viaje que realicen.
Variación temporal en los comportamientos madre - cría
La relación materno-filial mostró variaciones entre diciembre y febrero y se vio influenciada por las interacciones macho-hembra, macho-cría y cría-cría. En diciembre la proximidad pasiva fue el principal comportamiento de la díada, indicando la importancia del cuidado que la madre ejerce sobre la cría en los primeros días de su vida, protegiéndolo del resto de las hembras y de los machos dominantes que están en plena defensa de sus territorios. La frecuencia de amamantamiento aumentó sustancialmente de diciembre a enero. Consideramos que los valores de diciembre son bajos, ya que se espera que durante el periodo perinatal amamanten a sus crías frecuentemente para generar las reservas que le permitan sobrevivir a la primer ausencia materna (García-Aguilar & Aurioles-Gamboa, 2003). Sin embargo, la frecuencia y el tiempo que los cachorros maman no es necesariamente indicativo de la cantidad de leche que ingieren, por lo que estos parámetros no deberían ser utilizados como índice de lactancia (Cameron, 1998). En algunas especies de mamíferos la estimulación producida por la succión de leche produce inhibición de los ciclos ováricos en la hembra, mientras la intensidad y/o frecuencia de amamantamiento es elevada (Gomendio, 1994a). Debido a que en esta especie la ovulación y la cópula ocurren durante el período perinatal, la baja frecuencia de amamantamiento observada en diciembre podría interpretarse como un mecanismo que evita la inhibición del ciclo reproductivo. Los fócidos presentan una elevada y constante intensidad de amamantamiento durante el corto cuidado materno, de forma tal que las crías tienen una elevada tasa de crecimiento diaria aumentando varios kilogramos antes del destete (Bonnes & Bowen, 1996; Boyd, 1998; Trillmich, 1996). La cópula ocurre una vez finalizado el cuidado de la cría, probablemente debido a que el amamantamiento tiene efectos de retroalimentación negativa sobre la liberación de gonadotropinas inhibiendo el estro y la ovulación en alguna medida (Fogden, 1971).
La disminución de la frecuencia de alarmas hembra-macho entre diciembre y enero se relaciona con la finalización del período de cópulas (Franco-Trecu, Bóveda & Tassino, 2009), lo que conlleva a que disminuya el número de machos en el área (Franco-Trecu, 2005). La disminución de la proximidad pasiva y el aumento en la frecuencia de amamantamiento en enero, indican un cambio en el uso del tiempo por parte de la díada, asignándolo principalmente al amamantamiento. El tiempo dedicado al contacto pasivo aumentó de enero a febrero mientras que el tiempo de proximidad activa disminuyó, indicando que luego del reencuentro con su cría las hembras marcadas presentaron una actividad mínima, debido probablemente a la importante inversión energética que realizaron durante los viajes de alimentación.
Debido a que el componente vocal es el principal factor del reconocimiento y reencuentro de la díada (Charrier, Mathevon & Jouventin, 2001; Charrier, Mathevon, Hassnaoui, Arraro & Jouventin, 2002; Phillips & Stirling, 2000, Trimble & Insley, en prensa), la vocalización en la primera etapa del vínculo es sumamente importante. A pesar de que la frecuencia de las vocalizaciones de las hembras mostró diferencias significativas únicamente entre enero y febrero, se registró una disminución de las mismas a lo largo del período de estudio. Por otro lado el contacto naso nasal y el olfateo, también involucrados en el reconocimiento materno filial, mostraron una tendencia decreciente desde diciembre a febrero.
Las alarmas de las hembras hacia los cachorros aumentaron de enero a febrero, probablemente a causa de la mayor movilidad de las crías a medida que crecen, de las prolongadas ausencias de sus madres o de los frecuentes intentos de alo-amamantamiento registrados (Franco-Trecu, Tassino & Soutullo, en prensa). Este último comportamiento también puede ser la causa del aumento de las agresiones entre cachorros (Franco-Trecu et al., en prensa), que a pesar de no mostrar diferencias significativas exhibieron una tendencia al aumento en el mismo período, con un valor de probabilidad marginal. Respecto a la variación diaria de los comportamientos la mayor frecuencia de amamantamiento a la hora 7 puede relacionarse con el horario de menor temperatura, ya que en otros momentos las hembras termorregulan en el mar.
Los resultados que se desprenden de este trabajo son interesantes y novedosos, a la vez que generan nuevas interrogantes sobre la biología de esta especie. En este contexto se propone a las especies de pinnípedos en el Uruguay, como excelentes modelos para poner a prueba diferentes hipótesis mediante estudios longitudinales en ecología del comportamiento.
Agradecimientos
Los autores desean agradecer a Dr. Ciro Invernizi, Dr. Federico Riet, Dra. Helena Katz, y Lic. Carolina Abud por sus valiosos aportes al manuscrito, a los loberos Olivera, Veiga y Malo y a Rosina Frau por su colaboración en el muestreo.
Referências
Antonelis, G. A., Stewart, B. S., & Perryman, W. F. (1990). Foraging characteristics of female northern fur seals (Callorhinus ursinus) and California sea lions (Zalophus californianus). Canadian Journal of Zoology, 68, 150-158. [ Links ]
Arnould, J. P. Y., Boyd I. L. & Socha, D. G. (1996). Milk consumption and growth efficiency in Antartic fur seal (Arctocephalus gazella) pups. Can. J. Zool., 74, 254-266. [ Links ]
Blanckenhorn, W. U. (2005). Behavioural causes and consequences of sexual size dimorphism. Ethology, 111, 977-1016. [ Links ]
Begon, M., Harper, J. L. & Townsend, C. R. (1999). Ecología. Individuos, poblaciones y comunidades. (3ª ed.). Barcelona. Ediciones Omega. [ Links ]
Boltnev, A. I., York, A. E. & Antonelis, G. A. (1998). Northern fur seal young: interrelationships among birth size, growth, and survival. Can. J. Zool., 76, 843-854. [ Links ]
Boness, D.J. & Bowen, W.D. (1996). The evolution of maternal care in pinnipeds. Bioscience, 46, 645-654. [ Links ]
Boyd, I. L. (1998). Time and energy constraints in pinniped lactation. The Am. Nat., 152, 717-728. [ Links ]
Cameron, E. Z. (1998). Is suckling behaviour a useful predictor of milk intake? A review. Anim. Behav., 56, 521-532. [ Links ]
Carranza, J. (1994). Etología. Introducción a la ciencia del comportamiento. En: J. Carranza (Ed.). Reproducción (pp. 341-361). Universidad de Extremadura. [ Links ]
Charrier, I., Mathevon, N. & Jouventin, P. (2001). How does a fur seal mother recognize the voice of her pup? An experimental study of Arctocephalus tropicalis. The Journal of Experimental Biology, 205, 603-612. [ Links ]
Charrier, I., Mathevon, N., Hassnaoui, M., Arraro, L. & Jouventin, P. (2002). The Subantarctic fur seal pup switches its begging behaviour during maternal absence. Can. J. Zool., 80, 1250-1255. [ Links ]
Clutton Brock, T. (1991). The evolution of parental care. Princeton. Princeton University Press. [ Links ]
Cody, M. L. (1966). A general theory of clutch size. Evolution, 20, 174-184. [ Links ]
Costa, D. P. & Gentry, R. L. (1986). Fur Seal. Maternal Strategies on land and at sea. In: Gentry and Kooyman (eds.) Free ranging and reproductive energetics of the northern fur seal (pp. 79-101). Princeton. Princeton University Press. [ Links ]
Dirección Nacional de Recursos Acuáticos. (2003). Informe sectorial pesquero 2000 - 2001. Montevideo, Uruguay: Ministerio de Ganadería, Agricultura y Pesca. Disponível em: http://www.dinara.gub.uy/web_dinara/ [ Links ]
Fisher, R. A. (1930). The Genetical Theory of Natural Selection. Clarendon Press, Oxford, UK. [ Links ]
Fogden, S .C. (1971). Mother-young behaviour at grey seal breeding beaches. Journal of Zoology London, 164, 61-92. [ Links ]
Franco-Trecu, V. (2005). Comportamiento maternal y aspectos reproductivos de Arctocephalus australis, en Isla de Lobos, Uruguay. Licenciatura, UDELAR, Montevideo, Uruguay 74 pp. [ Links ]
Franco-Trecu, V., Bóveda A., & Tassino, B. (2009). Reclassification of South American fur seals mating system in Uruguay. Abtracts of the 46th Annual Meeting of the Animal Behavior Society (p. 79). Pirenópolis, Brazil. [ Links ]
Franco-Trecu V., Tassino B., & Soutullo A., (en prensa) Allo-suckling in the South American fur seal (Arctocephalus australis) in Isla de Lobos- Uruguay: cost or benefit of living in group?. Ethology, Ecology and Evolution [ Links ]
García-Aguilar, M. C., & Aurioles-Gamboa, D. (2003). Cuidado materno en el lobo marino de California de los Islotes, Golfo de California, México. Ciencias Marinas, 29(4B), 573-583. [ Links ]
Gentry, R. L. (1998). Behavior and ecology of the Northern Fur Seal. Princeton: Princeton University Press. [ Links ]
Gentry, R. L., & Kooyman, G. L. (1986). Fur Seal. Maternal Strategies on Land and at Sea. In Gentry & Kooyman (Eds.), Introduction (pp. 3-27). Princeton: Princeton University Press. [ Links ]
Goldsworthy, S. D. (2006). Maternal strategies of the New Zealand fur seal: Evidence for interannual variability in provisioning and pup growth strategies. Australian Journal of Zoology, 54, 31-44. [ Links ]
Gomendio, M. (1994a). Etología. Introducción a la ciencia del comportamiento. In J. Carranza (Ed.), La evolución del cuidado parental (pp. 407-441). Universidad de Extremadura. [ Links ]
Gomendio, M. (1994b). Etología. Introducción a la ciencia del comportamiento. In J. Carranza (Ed), Estrategias individuales de inversión parental (pp. 443-464). Universidad de Extremadura. [ Links ]
Guinet, C., Goldsworthy, S. D., & Robinson, S. (1999). Sex differences in mass loss rate and growth efficiency in Antartic fur seal (Arctocephalus gazella) pups at Macquarie Island. Behav. Ecol. Sociobiol., 46, 157-163. [ Links ]
Krebs, J. R., & Davies, N. B. (1987). An introduction to behavioural ecology (2nd ed.). Oxford: Blackwell Scientific Publications. [ Links ]
Lehner, P. N. (1996). Handbook of ethological methods. Cambridge: Cambridge University Press. [ Links ]
Lezama, C. (2002). Daños ocasionados por el león marino Sudaméricano (Otaria flavescens) a la pesca artesanal en el puerto de Piriápolis. Informe de Pasantía. Sección Zoología de Vertebrados. Facultad de Ciencias. Universidad de la República Oriental del Uruguay. [ Links ]
Lunn, N. J., Boyd, I. L., & Croxall, J. P. (1994). Reproductive performance of female Antartic fur seals: the influence of age, breeding experience, environmental variation and individual quality. J. Anim. Ecol., 63, 827-840. [ Links ]
Lunn, N. J., & Arnould, J. P. Y. (1997). Maternal investment in Antartic fur seals: evidence for equality in the sexes? Behav. Ecol. Sociobiol., 40, 351-362. [ Links ]
Luque, S. P., & Aurioles Gamboa, D. 2001. Sex differences in body size and body condition of California sea lion (Zalophus californianus) pups from de Gulf of California. Marine Mammals Science, 17(1), 147-160. [ Links ]
Martín, P., & Bateson, P. (1991). La medición del comportamiento. Madrid: Alianza. [ Links ]
Naya, D. E., Vargas, R., & Arim, M. (2000). Análisis preliminar de la dieta del león marino del Sur (Otaria flavescens) en Isla de Lobos, Uruguay. Boletín Sociedad Zoológica Uruguay, 12, 14-21. [ Links ]
Naya, D. E., Arim, M., & Vargas, R. (2002). Diet of South American fur seals (Arctocephalus australis) in Isla de Lobos, Uruguay. Mar. Mamm. Sci., 18(3), 734-745. [ Links ]
Páez, E. (1996). Simulaciones estocásticas en la población de Otaria flavescens en Uruguay. In Libro de Resúmenes, 7º Reunión de Trabajo de Especialistas en Mamíferos Acuáticos de América del Sur. SOLAMAC (p. 116).Chile: Viña del Mar. [ Links ]
Páez, E. (2005). ¿Yo...? Otaria. ¿Y usted...? In Libro de Resúmenes, III Jornadas de Conservación y Uso Sustentable de la Fauna Marina (p. 28). Montevideo, Uruguay. [ Links ]
Phillips, A. V. (2003). Behavioral cues used in reunions between mother and pup South American fur seals (Arctocephalus australis). J. Mammal., 84(2), 524-535. [ Links ]
Phillips, A. V., & Stirling, I. (2000). Vocal individuality in mother and pup South American fur seals, Arctocephalus australis. Mar. Mamm. Sci., 16, 592-616. [ Links ]
Ponce de León, A. (1983). Aspectos de la reproducción, crecimiento intrauterino y desarrollo postnatal del lobo de dos pelos sudamericano Arctocephalus australis (Zimmermann, 1783) en islas de la República Oriental del Uruguay. Tesis Licenciatura Oceanografía Biológica, Universidad de la República, Facultad de Humanidades y Ciencias. [ Links ]
Sokal, R. R., & Rohlf, F. J. (1979). Biometría. Principios y métodos estadísticos en la investigación biológica. Universidad de Málaga. [ Links ]
Soto, K. H., Trites, A. W., & Arias-Schreiber, M. (2004). The effects of prey availability on pup mortality and the timing of birth of South American sea lions (Otaria flavescens) in Peru. J. Zool. Lond., 264, 419–428. [ Links ]
Szteren, D. (1999). Impacto económico del león marino Sudamericano (Otaria flavescens) en la pesca artesanal en Uruguay. Tesis de Maestría en Manejo de Vida Silvestre. Centro de Zoología Aplicada, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. [ Links ]
Szteren, D., & Páez, E. (2002). Predation by southern sea lions (Otaria flavescens) on artisanal fishing catches in Uruguay. Marine and Freshwater Research, 53, 1161-1167. [ Links ]
Szteren, D., Naya, D. E., & Arim, M. (2004). Overlap between pinniped summer diets and artisanal fishery catches in Uruguay. LAJAM, 3(2), 119-125. [ Links ]
Trillmich, F. (1986). Maternal investment and sex allocation in the Galapagos fur seal, Arctocephalus galapagoensis. Behav. Ecol. Sociobiol., 19, 157-164. [ Links ]
Trillmich, F. (1990). The behavioral ecology of maternal effort in fur seals and sea lions. Behaviour, 114, 1-4. [ Links ]
Trillmich, F. (1996). Advances in the study of behavior. In: Rosenblatt & Snowdon (Eds.), Parental investment in pinnipeds (pp. 533-577). California: Academic Press. [ Links ]
Trimble, M. & Insley, S. J. (In press). Mother-Offspring Reunion in the South American Sea Lion Otaria flavescens at Isla de Lobos (Uruguay): Use of Spatial, Acoustic and Olfactory cues. Ethology Ecology & Evolution 22(3). [ Links ]
Trivers, R. L. (1972). Parental investment and sexual selection. In Campbell (Ed.). Chicago: Aldine Press. [ Links ]
Trivers, R. L. (1985). Social evolution. Benjamin Cummings. California: Menlo Park. [ Links ]
Trivers, R. L., & Willard, D. E. (1973). Natural selection of parental ability to vary the sex ratio of offspring. Science, 179, 90-92. [ Links ]
Vaz Ferreira, R. (1950). Observaciones sobre la Isla de Lobos. Rev. Facultad de Humanidades y Ciencias. Universidad de la República Oriental del Uruguay, 5, 145-176. [ Links ]
Vaz Ferreira, R., & Sierra de Soriano B. (1963). Tolerancia en grupos biespecíficos de pinnípedos. In Libro de Resúmenes, 16º Internacional Congreso de Zoología (p. 250). Washington, D.C. [ Links ]
Vaz Ferreira, R. (1976a). Arctocephalus australis (Zimmerman), South American fur seal. Advisory Committee on Marine Resources Research. Scientific consultation on Marine Mammals. Bergen, Norway. [ Links ]
Vaz Ferreira, R. (1976b). Otaria flavescens (Shaw) South American sea lion. Advisory Committee on Marine Resources Research. Scientific consultation on Marine Mammals. Bergen, Norway. [ Links ]
Vaz Ferreira, R., Vallejo, S., Achaval, F., Melgarejo A., & Meneghel, M. (1981a). Parto y comportamiento perinatal en Arctocephalus australis (Zimmermann), lobo de dos pelos de Sudamérica, (Pinnipedia: Otariidae). In Resúmenes y Comunicaciones de las Jornadas de Ciencias Naturales (pp. 63-64). Uruguay. [ Links ]
Vaz Ferreira, R., Vallejo, S., Achaval, F., Melgarejo A., & Meneghel, M. (1981b). Comportamiento de baño postnatal y otras interacciones materno - filiales en Arctocephalus australis, lobo de dos pelos de Sudamérica. In Resúmenes y Comunicaciones de las Jornadas de Ciencias Naturales (p. 65). Uruguay. [ Links ]
Vila, B. L., & Cassini, M. H. (1990). Agresividad entre hembras y separación madre – cría en lobo marino del sur, en Chubut, Argentina. Revista Chilena de Historia Natural, 63, 169-176. [ Links ]
Wilson, E. O. (1980). Sociobiología: la nueva síntesis. Barcelona: Omega. [ Links ]
York, A., Lima, M., Ponce de León, A., Malek, A., & Páez, E. (1998). First description of diving female South American fur seals in Uruguay. Abstract WMMSC, Monaco. [ Links ]
Recibido en: 5/02/2008
Acepto en: 7/01/2010
![]() Dirección para correspondencia
Dirección para correspondencia
Valentina Franco Trecu, Iguá 4225, CP 11400, Montevideo-Uruguay. E-mail:pinnipedosuy@gmail.com. Tesis de Licenciatura de Valentina Franco Trecu. Sección Etología – Facultad de Ciencias - Universidad de la República Oriental del Uruguay.
* Presentado en: III Jornadas de Conservación y Uso Sustentable de la Fauna Marina. Organizadas por la ONG-PROFAUMA (Protección de Fauna Marina) y la Escuela Naval. Montevideo–Uruguay, 14 al 17 de Septiembre de 2005 y VIII Jornadas de la Sociedad Zoológica del Uruguay. Sociedad Zoológica del Uruguay, Facultad de Ciencias, 24 al 28 de Octubre de 2005.

