










Services on Demand
article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Share

 Permalink
PermalinkRevista de Etologia
Print version ISSN 1517-2805
Rev. etol. vol.11 no.1 São Paulo 2012
Evidence of learning in the web construction of the spider Argiope Argentata (Araneae: Araneidae)
Evidência de aprendizagem na construção de teia na aranha Argiope Argentata (Araneae: Araneidae)
Selene Siqueira da Cunha NogueiraI; César AdesII
IApplied Ethology Laboratory, Department of Biological Sciences, State University of Santa Cruz. Address: Rod. Jorge Amado km 16, Salobrinho, Ilhéus-BA, Brasil, 45662900. E-mail: seleneuesc@gmail.com / selene.nogueira@pq.cnpq.br
IIIn memoriam. Department of Experimental Psychology, Institute of Psychology, University of São Paulo
ABSTRACT
We investigated the effects of past experience on the web construction by the spider Argiope argentata. The spiders were submitted to periods in horizontal position alternated with periods in vertical position (control phase). The observations revealed an improvement in the quality of the second vertical web when compared to the first; disordered web-building behavior when first in a horizontal phase, with resultant web deformities; improvements in web quality after successive web constructions in a horizontal phase; finally, we observed improvements in spiders' performance and a shorter time needed to build their webs when the spiders were submitted to the horizontal phase for the second time. The results suggest that improvements during the horizontal periods were due to changes in the building patterns in the early stages of spider web construction (mesh, frame and radius constructions). Our results favor the hypothesis that this improvement is due to past experience or learning.
Keywords: orb-weaver spider, gravitational effect, trial and error, experience, web construction, flexibility.
RESUMO
Investigamos os efeitos de experiência passada na construção da teia da aranha Argiope argentata. As aranhas foram submetidas a períodos em suportes horizontais alternados por períodos de suportes verticais (condição controle). Os resultados revelaram melhora da primeira para a segunda teia vertical; desorganização do desempenho no primeiro contato com a condição horizontal; melhora do desempenho após sucessivas construções horizontais e finalmente melhor desempenho e menor latência de construção no segundo contato com a horizontal. Os resultados sugerem que a melhora, na condição de horizontalidade, decorre de uma mudança nos padrões iniciais da construção (feitura de amarras/quadro/raios) e apoiam a hipótese de que esta melhora seja um produto de experiência passada ou aprendizagem.
Palavras-chave: aranha orbitela, efeito gravitacional, ensaio e erro, experiência, construção de teia, flexibilidade.
Introduction
The web construction of Argiope argentata spider, like other orb-weaver spiders, occurs following a sequence of responses that come from a typical pre-programming (Witt, 1965). The process of orb webs building is considered an instinctive action, a stereotypical behavior, regardless of experience (Szlep, 1952). This orb web involves elements made from non-sticky and elastic threads: (a) the moorings, which hold the web to its environmental supports, (b) the frame, the boundary line, (c) the radials, (d) the center, composed of the core and a non-sticky spiral. It also comprises a lax, sticky thread (e) the sticky spiral whose job is to capture and retain the insects. In the web there are empty spaces (f), the periferal gap, between the frame and the viscous spiral and (g) the clear zone between the viscous and non-sticky spiral (Comstock, 1948).
The basic steps of construction (construction of the frame and radials, placement of the non-sticky spiral and temporary spiral, placement of the viscous spiral etc.) follow a sequence typical for the species in which each step provides the context for the next (Ades, 1973; Eberhard, 1969). This classical view regarding construction of orb-weaver spider webs assumes the absence of possible learning effects. An observation consistent with this classic issue, is that the spiders build structured webs from early stages after hatching and, even when deprived of direct experience of building in these stages, manage to build perfect webs when they later have the opportunity to do so (Witt, 1968). On the other hand, there is a very large operational flexibility inherent in the construction program due to external factors (Ades & Cunha, 1991; Eberhard, 1988; Enders, 1976; Harmer & Herberstein, 2009; Nakata, 2007; 2009; Tso et al., 2007). The spider adjusts its behavioral routines at every step of construction, it seems driven by commands like run "R" until it reaches a satisfactory condition "S" rather than by simple stimulus-response commands. Thus, web construction depends on a constant "reading" of the environment in which their performance occurs (Ades, 1986; Nakata, 2012).
There are two basic ways for animals to solve problems in nature. They may have been born with ready answers, pre-formed in the nervous system (instinctive responses) or they may have a small number of ready answers, but on the other hand also have the ability to change their behavior as a result of experiences as they develop (see Bouton, 2007). It is clear, for example, that the web's internal dimensions and parameters are influenced by the dimensions of the available bracket (Ades, 1986) or as a result of the food supply in a given location (Nakata, 2012). The spider will expand its web up to the upper limit of the bracket space, and the same strategy will influence the lower limit making it small (Ades, 1986; Eberhard, 1986). Eberhard (1988b) demonstrated the existence of flexibility in the structure of webs in Leucauge mariana and Micrathena sexspinosa. According to the author, an assessment of the available silk occurs in the glands and these spiders exhibit variations in the number of radials, in the symmetry of capturing spiral and in the space between the spirals.
Another interesting research data regarding the spiders' flexibility and environmental factors is the existence of relationship between the web's location and its size. Leborgne & Pasquet (1987) reveal that both solitary and social spiders use to "investigate" the neighborhood, and depending on the size of the spider population in the vicinity, the spider will build more or less extensive webs. Therefore, should be rejected the concept that orb-weaver spiders have a rigid program and inflexible geometry web patterns.
Vollrath (1987) reports exhaustively that the geometry of orb webs is not predetermined rigid. The author shows geometrical changes of webs built by spiders with regenerated legs (Agustin diadematus) after mutilation. It was observed that behavioral rules that determine the pattern of each spiral in these animals were different (Benforado & Kistler, 1973). The behavioral processes that spiders provide a temporary or auxiliary spiral and a capture spiral are distinct mechanisms. This proceeding could explain previous experiments results, which reported the importance of the gravitational forces as a "compass" to guide the spiders during the web construction (Vollrath, 1985; 1988). These experiments show that the rotation to the vertical plane can temporarily disorient web construction, resulting in webs with drastic degenerations in the capture spiral, leaving, however, the auxiliary spiral intact. Vollrath's experiments (1985; 1988) demonstrated two factors: the existence of two distinct routines in the construction of the viscous and non-sticky spiral and the possibility of gravitational orientation effectively establishing rules in the web construction.
Other authors warn about the existence of a flexible operational system in the spider's behavioral routines, which shows a constant interaction with their environment (Ades, 1991b; LeGuelte, 1966; LeGuelte & Ramousse, 1979; Nakata, 2012; Witt & Reed, 1965). These observations are inherent for both hunting behavior and positioning of capture network. These factors, however, do not guarantee the existence of learning process. Thorpe (1963) defines learning as a process that manifests itself by adaptive changes in individual behavior as a result of experience. Considering construction as a complex program, it is important to check which environmental factors are taken into account by the animal, and in which way they are - one of these possible factors is the Earth's gravity.
There are species of orb-weaver spider that build their webs, both in nature and in artificial laboratory conditions, on an approximately horizontal plane while others use a vertical plane. It is observed that each species adapted its behavioral routines to the spatial disposition of the webs (Caramaschi, 1991; Edrich, 1977; Jander, 1975; Opell, 1987; Reed, 1969; Seyfarth et al., 1982). Gravity's influence is shown, among other things, by an up-down asymmetry of the webs constructed on a vertical plane. For Argiope argentata, the upper region of the vertically built web tends to be smaller, with fewer radials, wider angles between them and less extensive viscous spirals than in the lower region. Such asymmetry was determined by Ades (1973), who observed the species using a strategy of placing incomplete spirals, spirals that do not return full-circle, especially when it is at the peripheral and lower region - and that would require a supplementary fill - the spider breaks its placement trajectory of the viscous spiral and reverse its path, disposing of the following spiral. It is a "direction reversal", which also can be seen, in the finished web, from the number of "broken" spirals (Ades, 1973).
Gravity may not be the only generating factor in the up-down asymmetry (see Nakata, 2012), but without any doubt, it is directly linked to the placement of the web as a process of orientation, in which the spider takes benefit of a physical factor to gain advantage for prey capture (Maciejewski, 2010; Masters & Moffat, 1983; Nakata, 2012). The gravity influence can be experimentally manipulated in several ways. A classic way is to increase the spider weight, observing the changes that occur in the structure of the constructed web, in which the webs become more eccentric (Baum & Witt, 1962; Le Guete, 1967; Witt & Baum, 1960). A more drastic manipulation can be achieved in very exceptional circumstances, such as on a space ship, outside the Earth's gravitational field. Witt, Scarboro & Daniels (1977) obtained some data on web construction of two specimens of Agustin diadematus that were sent to the Moon by the Skylab project. In that study, the authors concluded that (1) the absence of gravity's effect turns the webs more symmetrical than in its presence; the number of angles at the top of the web was equal to the number of angles at the bottom, and also observed a decrease in direction reversals in the construction of viscous spirals; (2) is there an possible effect of "learning" in web construction in weightless conditions. Witt et al. (1977) reported that the initial webs were worse, in structural terms, than the subsequent ones. However, there isn't an accurate or quantitative record to support this conclusion. The work shows that gravity is one of the "clues" that an orb spider uses during web construction and that determines its typical asymmetry, and suggests that some behavioral adjustment can take place, similar to "training" (operational manipulation with or without reinforcement in order to modify a behavior).
Witt (1968) observed another learning indication when manipulated the weight of Agustin diadematus. The author noted that the thread of provisional spiral thickened when its weight was increased, and after returning to normal weight, prior to the experiment, the spiders slowly returned to the initial thread thickness. The author explained that this effect, returning to a standard thread thickness, could be a learning process, in which the spider would have gradually adjusted the necessary metrics. This hypothesis, however, has not been systematically investigated.
An easier way to manipulate the gravity's effects would be to force the spiders to build their webs in space differently to those that they would normally choose. By arranging supports, for example, Argiope argentata can be prompted to build webs vertically (preferred direction) or horizontally. In horizontal conditions, gravity is not eliminated, but is distributed equally throughout the building space, and therefore, cannot serve as a differential clue, because there is no "above" or "below" in a horizontal web. Through this structural manipulation, making comparisons between structures of webs built horizontally and vertically, it is possible to know how gravity participated in the spider's environmental "reading" during the construction program. Braga et al. (1986) studding Argiope argentata webs in the wild found that there were changes in the size of the webs. The horizontal webs were smaller than others built vertically. The average indices of peripheral spaces also confirmed the effect of the horizontal position on the size of the web. In addition, the data showed that there was lower usage of available space in the construction of the web. In a later study with Argiope argentata, coming from a limited number of observations, Cunha Nogueira & Ades (unpublished data) found that webs initially built horizontally - a dimension rarely used by this free range spiders - had significantly greater irregularities than the subsequent ones, built in the same conditions. This found suggests that the spiders had their performance partially affected by the change in support layout (from vertical to horizontal) and might have adjusted their performance based on new values of gravitational stimulation from the "training" (Ades & Cunha, 1991).
Ades (1989) analysing several problems faced by free range animals, such as prey distribution, presence or absence of predators, nest construction or a web, reports that the relevant information to such behaviors are not limited just in the immediate environment, as the adaptive performance would depend on the use of previously acquired experience - "...Learning and memory are, in spiders as in other animals, part of a general strategy of adaptation." (p. 18).
In this context, our aim was to study the gravity's influence on Argiope argentata's web construction. We quantify the web parameters changes through the previously established web, checking the influence of the spider past experience or evidence of learning by training web construction, by using paradigms (habituation, Pavlovian or functional conditioning), in which animals would have the opportunity to build successive webs in an environment favoring horizontal webs (or vertical) and then tested under conditions that favor vertical webs (or horizontal). The relevance of this work lies in the possibility to contribute to the related concept "instinct" and "learning" (Ades, 1982; Ades, 1987; Hollis, 1984).
Material and methods
Experimental animals and study site
We captured 23 adult females of Argiope argentata in the campus sites at the Universidade de São Paulo - USP, between 1989 and 1990. The animals were captured with an average weight of 90 mg. We discarded four individuals because they did not adapted to the experimental conditions, thus the study comprised 19 animals. After their capture the spiders were placed in boxes (details bellow) and maintained in the Spider Laboratory at the Institute of Psychology, Universidade de São Paulo - USP. The boxes were made with a wooden frame (28.0 cm height X 27.5 cm width X 5.0 cm depth), enclosed by two transparent glass plates, 0.5 mm in thickness. In these boxes, when arranged vertically, the spiders often built webs considered perfect, slightly tilted, but similar to those found in natural conditions. Inside the lab, the boxes were arranged on metal racks with 15 cm gaps between them with natural lighting provided by a wide window, without ever being subjected to direct sunlight. We offered a fly to the spiders three times a week.
Experimental phases
The basic experimental manipulation was to leave the spiders to build webs: a) with the box in an upright position (V), according to a standard maintenance condition; b) with the box lying in a horizontal position (H). In the horizontal position, the available height was only 5 cm, with no possibility of building a web close to vertical, because it was difficult for spiders to attach threads on glass surfaces. The spiders had a virtually horizontal space to build their webs. In this position, the spider received a lower lighting level than in the vertical box. To minimize the difference in brightness, the racks were lined with white paper to guarantee a maximum reflection and uniformity of light.
The spiders went through five alternate and successive experimental phases: vertical (V1 and V2) - horizontal (H1) -vertical (V3) - horizontal (H2) - vertical (V4). The V1 phase (control) refers to the time interval between placing the spider in the box, left in an upright position, and complete construction of the first web by the spiders. The Phase V2 (control) refers to the time interval between the destruction of the first web through the removal of threads that could serve as guides for web reconstruction and complete construction of the second web. The phase H1 refers to the time interval between the placement of the box in a horizontal position and the construction of the first web, after the destruction of the vertical web. The duration in horizontal phase, either in the first horizontal condition (H1), or in the second (H2) was no more than 25 days, regardless of the achieved level of construction. A prior study indicated that the spider did not show web improvements after this period. There was no destruction of H11, H12 and subsequent webs could therefore rely on previous threads. We decided for this paradigm to do not difficult too much for the animals building, because they were in adverse condition. This action was taken after some deaths of four animals upon the withdrawal of previous threads in the horizontal phases. The phase V3 (control) refers to the third web built with the box placed in a vertical position, after the destruction of the web or prior structure, built in a horizontal position. Phase H21 refers to the time interval between the first web or structure built in the second stage, after destruction of the web or prior vertical structure. There was record of H22 and H23 in some spiders, following the same previous procedure without destruction of H21. The last phase V4 (control) corresponds to the fourth web built with the box in upright position, after the destruction of the previous web or structure. The total removal of previous construction (in the case of VI, V2, Hl, V3, H2 and V4) aimed to avoid a "guide" effect from the previously placed threads. It is known that spiders use previous web frameworks as a guide in the following construction (Ades, Hardt & Sassaki, 1986).
Data collection
The spiders were removed from their web or structure, as soon as its construction was completed, and kept in a jar during the photographic recording of the web. The animals' removing provides a complete picture of the web threads. After record the web by taken its picture, the spider was put back in the box and fed with a fly.
The photographs were obtained by putting the animals boxes in a picture box, wooden made that provide contrasting conditions for taking pictures that highlight spider's webs. This picture box had a cube shape (~ 60 cm2) and was internally lined with black cloth. Four fluorescent lamps, (15 W) were positioned to illuminate only the web structure. Whenever possible, the glass plates were removed, to avoid possible light reflections that could reduce the contrast in the photographs. Sometimes, however, there were threads fixed to the box's glass, unable to be removed. Next to the web, during the pictures, we positioned a standard unit (1 cm) as pattern for posteriori measurement of the webs. The photos were taken using black and white film (Panato C and Plus X, Kodak, Japan).
Data analysis
Measurement of web dimensions was performed by means of a modified software called Diracom 3 (executed by R.C. Borra) installed on a ffiM computer, compatible with AT 286, equipped with an IRIS-16 image acquisition board (Micro-image) and connected to a Tarnron Fotovix camera (video film processor model 40, USP-SP) and two video monitors, one color and one analogue. To measure the webs, we used the image of the webs produced by Fotovix, which was showed on computer monitor. This procedure allowed us to transform the image and the contrast to the amplifier factor. To got a better image, we maximized the web threads contrast.; the adjustment factor and expansion was regulated through the standard 1 cm contained in the frame. Following this procedure, all the images had the natural equivalent dimensions and the measurements could be taken in centimeters. In the scanned images, the measurements started from an arbitrary central point of the web (the refuge) that was established as criterion. We performed automatic calculations of the following parameters: total area of the web (or coverage), internal area, capture area, the angle between the radials, number of radials and web eccentricity index in V2 and H2; this index was calculated by the number of upper spirals divided by the number of lower spirals. To analyze the phases H1 and H2, the development of each web was considered according to three criteria: 1) capture area, 2) construction latency 3) construction level. For the analysis of web parameters and level of construction, the Chi-square (χ2) and Wilcoxon tests were used at a p<0.05 significance level. For each web built in a horizontal phase, a score of construction level was assigned ranging from 1 to 5, based on the classification criteria mentioned above. Thus the indices of 1 to 5 indicate the increasing quality of the webs (1 - worst and 5 - best). These indices were assigned to the webs through assessment conducted by three laboratory assistants who were not involved in the study, taking into account the web construction quality.
Results
Changes and consistencies in vertical webs: qualitative and quantitative aspects
We observed a remarkable improvement in V2 phase when compared with V1. It was observed that the V2 webs were larger than the V1 webs; the animals took more advantage of available space (Fig. 1). The V2 webs showed more regular framework (structural base of the web) with most of the radials at right angles to the box's wooden frame. The spider seemed to build easier by placing the initial "Y" (fork shape) and later structures between phases V1 and V2. Despite the qualitative differences found between V1 and V2, the basic pattern of most webs constructed in a vertical was not modified. The spiders constantly retained the structural characteristics that define a typical web. On the other hand, qualitative analyses did not detect striking differences between the subsequent vertical webs, comparing with the same parameters among phases V2, V3 and V4.
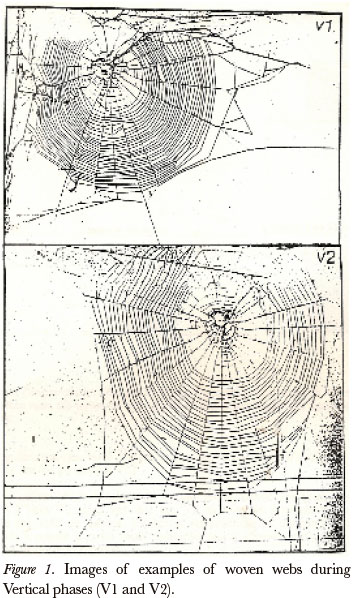
With respect to the quantitative aspects of the parameters analyzed between V1 and V2, significant differences were observed in almost all cases, except for the value of the internal area, in which the HO was not rejected (Wilcoxon, gl = 17; T = 45 P > 0.05). There was a significant difference of the following structures: total area (Wilcoxon, gl = 17, T = 22 P < 0.01), capture area (Wilcoxon, gl = 17; T = 21 P < 0.01), angles between radials (Wilcoxon, gl = 17; T = 20 P < 0.01), radials (Wilcoxon, gl = 17; T = 17 P < 0.01). The coverage of the webs was greater in V2 than in V1 (total area, internal area, capture area, number of radials greater in V2 than in V1; angle between the radials lesser in V2). Web regularity was also greater in V2 than in V1 (standard deviation of the angles lesser in V2 than in V1). No significant changes were observed when comparing the webs obtained from V2 and V3 (Wilcoxon, gl = 18, p > 0.05). With respect to comparisons between V3 and V4 of the studied parameters, it is important to note the total area measurements and capture area in V4 showed a slight increase in comparison with V3, but there were no significant differences observed when analyzing the parameters (Wilcoxon gl = 14 P > 0.05). Some spiders did not construct a web in V4, this fact may be related to the age of the animals.
Webs or horizontal structures: qualitative and quantitative aspects
The web building process was analyzed to determine if the sequential pattern remained. It was observed that this specific characteristic of the Argiope argentata did not change, however, unlike what happens in normal webs, the completion of one structure may not trigger the next step, i.e., the spider can stop the construction at the end or in the middle of a certain structure, not reaching the final completion of the web construction.
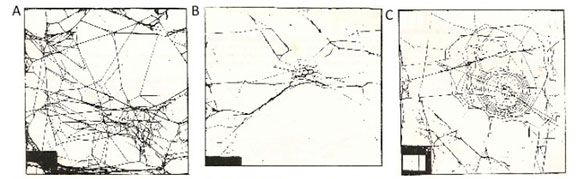
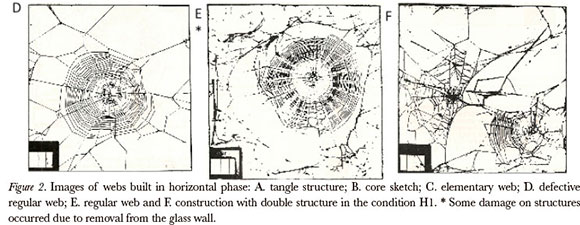
The constructions were classified according to overall and qualitative criterion with the assignment of values corresponding to the structure quality: a. Tangle (level 1): this structure is characterized by the presence of non-sticky threads arranged apparently randomly, attached at points with each other, but without any orb web structure. The tangle can be more or less dense, and can be found in association with other structures, often giving the impression that the spider takes advantage of random threads to attach a new web (Fig. 2.A); b. Core Sketch (level 2): one can see in this construction a few converging threads and, mainly, the placement of a few spirals (roughly parallel wires, placed according to the perpendicular rule to the radials). This structure can appear in the middle of a tangle or be supported, by a few threads, to the bracket (box) (Fig. 2.B) c. Elementary Web (level 3): this construction is characterized by the presence of the basic structures of an orb web - moorings, frame threads, radials and spirals - with defects in the substructure (irregular spacing between the viscous spirals, viscous spirals stuck to each other, radials partially or totally attached) and overall features (few radials, few spirals, viscous or otherwise), the core is well defined. It is a small web in comparison with regular webs; (Fig. 2.C) d. Regular Defective Web (level 4): in this construction there are all the structures of an orb web, endowed with regularity, however there are partial maladjustments: some spirals stuck together, irregular spacing of the viscous spirals (distances between irregular spirals), some radials stuck together, but having a "perfect" vertical web configuration (Fig. 2.D); e. Regular Web (level 5): The regular web is the "typical" Argiope argentata vertical web when found in nature, with its specific characteristics: moorings arranged on the bracket supporting the frame threads and radials, spirals running in parallel, refuge well bordered by a clearance (fig. 2.E).
Web eccentricity
The eccentricity of the horizontal webs approached one, differently of eccentricity in the vertical webs (H21 x V2 (Wilcoxon, T = 1 P < 0.01); H21 x V3 (Wilcoxon, T = 1 P < 0.01); V2 x Hll (Wilcoxon, T = 1 P < 0.01); H21 x V4 (Wilcoxon, T = 1 P < 0.01). This loss of eccentricity is possibly due to the absence of the gravity differential effect on the various parts of the web in a horizontal phase.
Temporal construction development and structure quality
During the H1 phase, we observed an unexpected emergence of a certain structure, which composes the complete web construction, for example, the emergence of an elementary web after a core sketch or a tangle. Some spiders, when placed in horizontal, presented building steps that, over time, through successive attempts, led to improvement of its web (Fig. 2). However, throughout these attempts, emerged more complete structures than the previous ones. In addition to the emergence of a particular framework, we observed a regression in the horizontal web quality. The spiders had as objective to improve their web, but sometimes this process stops, and the spider can go back, building a less regular web than the one before. Double constructions (Fig. 2.F), in the same period of construction, show a partial or total elimination of the previous construction phase and perhaps indicate an exacerbation of the behavior in searching for an adequate micro-site.
Change in construction from H11 to H12
Regarding the level of construction or web quality in H11 and H12, a small qualitative improvement in the web levels was observed from H11 to H12, but this was not statistically significant (Wilcoxon, gl = 19; T = 70.5 P > 0.05). In quantitative analysis, however, the difference between H11 and H12 did not taken into account the rudimentary constructions (Tangle and Core sketch), because it was impossible to measure its parameters. In this way, seven pairs of webs were counted for comparison. The comparison, however, did not unveil significant results: total area (Wilcoxon, gl = 7; T = 5 p > 0.05); internal area (Wilcoxon, gl = 7; T = 11 P > 0.05); capture area (Wilcoxon, gl = 7; T = 6 P > 0.05); angles (Wilcoxon, gl = 7, T = 12 p > 0.05); radials (Wilcoxon, gl = 7; T = 11 P > 0.05).
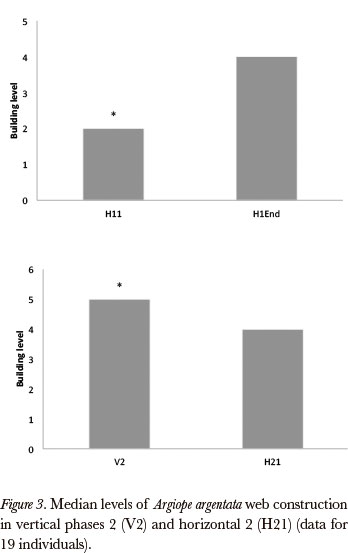
Additionally, during the first horizontal stage, most spiders that made a tangle in H11 built at least a rudimentary web in H12 (10 among 11 spiders). In H11, 47% of webs were constructed to at least a rudimentary level, in H12, on the other hand, there were 88.9% (χ2 = 5.5, P < 0.05). Moreover, the last web or built structure (H1End) was considered, because this is the web that one would expect the biggest change. Significant differences were noted in this last construction when compared with the H11 (Wilcoxon, gl = 14; P < 0.02) (Fig. 3).
Changes in construction from H11 to H21 and comparison of V2 with H2
In H2, it was observed that the first web obtained in this condition was more regular than the first obtained in H11, and it was significant in terms of the construction level (Wilcoxon, gl = 18; T = 24 P < 0.01).
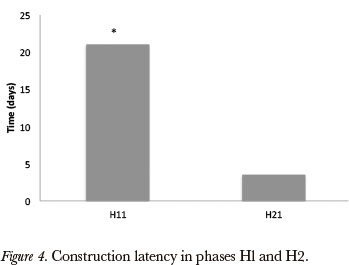
We observed that latency for the construction in the H2 phase was much lower than in H1 (Wilcoxon test gl = 16; T=5P < 0.01, Fig. 4). The data suggested, therefore, that in H2 the spiders were more readily to build better quality webs than in Hl. In H1 and H2 phases, we compared webs that at least reached the level 3 (Elementary web). This comparison showed no significant difference (Wilcoxon, gl = 9; P > 0.05). The analysis between V2 and H2, the best webs obtained from each one phases, showed that V2 were better than those found from the best horizontal web in all parameters: total area (Wilcoxon, gl = 17; T = 6 P 0.01), internal area < (Wilcoxon, gl = 17; T = 6 P < 0.01), capture area (Wilcoxon, gl = 17, T = 6 P < 0.01), angles between radials (Wilcoxon, gl = 17; T = 19 P < 0.01), number of radials (Wilcoxon, gl = 17; T = 21.5 P < 0.01) (Fig. 3). We observed that the spiders in V2 had a higher performance when compared with H21, (Wilcoxon, g1 = 18; T = 28.5 P < 0.02). It was observed that, regarding the webs' eccentricity index; the horizontal webs were more "centric" than the vertical webs (~ 1.0), horizontal construction pattern.
Discussion
On regard to flexibility and effects of prior experience in web construction, the variability of the web's geometric dimensions have been known for a long time and it was taken as an argument in favor that "instinct is not rigid". Tilquin (1942) reports that the changes in the web size of orb-weaver spiders are also accompanied of changes in its parameters. In the radials, for example, as larger are the web, greater are the number of radials. The experimental manipulation may show how the adjustment of construction to environmental conditions is systematic (Reed, Scarboro & Witt, 1969). Spiders make their webs larger or smaller according to the available space, including modifying the web's typical structural relationships (Ades, 1987), or even when there is a greater availability of food (Nakata, 2012).
The spider flexibility can be observed by the minuteness of motor behavior used during the positioning of threads. Vollrath (1987) reports that amputated spiders can reorganize the function of their remaining legs to produce a functional web. In experiments from drug ingestion, for example, they can still have disorders in their web construction behavior (Reed & Witt, 1968; Jackson, 1974). This variability, although an interesting phenomenon and of theoretical relevance, has nothing necessarily to do with effects of past experience: it only portrays the animal's immediate adjustment to construction conditions. After the passing of the drug's effects, the spider starts to build normal webs, that is, as they did before ingesting the drug.
Our study indicated that besides the spider momentary flexibility, resulting from open behavioral program, there is a lasting modification of behavior linked to the construction of the web. In addition, our study raised the question on the existence of learning effects on a pattern previously seen as stereotyped. Two instances of construction modification were detected, the first during the spider's first contact with the box (going from V1 to V2), the other occurred on contact with this box when placed horizontally (going from H1 to H2). It was observed, in the first case, that the V2 webs differed from V1 webs in a series of parameters, such as size, and presumably betters to capture insects. The difference between V1 and V2 must be understood as a decrease of construction quality in V1, considering the quality standards achieved in the environment and a recovery of this quality in V2 - an adaptation to the new lab environment. If we had taken measurements of the webs in the natural environment, this inference could be quantitatively proven, however we did not. Such difference from V1 to V2 cannot be attributed to a nutritional deficiency of the spiders, as might be supposed, because of the lengthy period that they spent in the laboratory, before they built their first web. To avoid nutritional problems, even without web, we fed the spiders regularly, by directly offering the insect to the mouth using tweezers, which adequately guaranteed their nutrition. Witt (1968) studying spider deprivation, reports their adaptation from nature to the laboratory. Spiders raise in narrow environments, when released, could built webs for the first time, producing smaller constructions than those of control spiders. However, they can expand their webs as they get more experience.
A second explanation looks at stress factors, and not in cognitive factors, as a reason for the decline and recovery of the performance. The procedures of capture and transport until get the lab, would have generated a defensive state, causing construction inhibition. Against this hypothesis, we could mention the fact that the destruction of the web (after V1 and before V2), which involved disturbance, did not perpetuate the supposed state of inhibition or disturbance. In addition, we must remember that the first laboratory construction occurred after a reasonable time lapse, during which it is assumed that the stressed state should have subsided. The design of this experiment does not yet allow a definitive choice between these hypotheses. It is interesting to note that the level of construction, as well as the values of the various parameters did not change from V2 to V3, or from V3 to V4. The construction of webs in horizontal box phase does not affect the performance in the vertical position.
One of the most curious structures found in the horizontal box phase was the tangle, the set of threads that occupies part or all of the available space. We observed that threads were not loose or positioned independently of each other; the networks had connection points. One of our questions was - what would be their functional origin? They could be attributed to a simple unordered escape route, without taken account a web-building program. They could, on the other hand, be seen as a disorganized web construction. The spider would find itself, in horizontal artificial conditions, in a mentally disorganized state, similar to what happens when subjected to psychotropic drugs or lesions in the central nervous system (Christiansen, Baum & Witt, 1962; Jackson, 1974).
A third very plausible hypothesis is that the spider is, by weaving the tangle, putting into action a strategy of looking for a suitable place to build the web. In nature, it is common to see threads going from leaf to leaf, close to where the web is built; these are possible spider relocation products that stop when a suitable location is found. In the horizontal box, the search for a construction site can be extended greatly, and involve the spider going back to sites already visited. In extreme cases, this search can be indefinitely extended, since, for the animal, there is a lack of appropriate stimuli in order to trigger the actual construction. The core sketch also suggests that there is an absence of appropriate stimuli, not for starting the construction, but for the completion of the preparation of the radials/frame/central non-sticky spiral.
The problem that the spider faces, in the horizontal web, is the need to change some of its behavior, selected to the vertical position, or the need to adjust its motor program to an impoverished support situation (Wiu & Reed, 1965; Wiu, Reed & Peakall, 1968). Gravity, in particular, dramatically changes construction: the spider cannot use it as it does in a normal situation. There is an improvement of construction in horizontal phases, when analyzing H11 x H1End, in terms of the number of spiders that built at least one rudimentary web. Since the H11 constructions were not destroyed, prior to the construction of H1End, the improvement could be explained non-cognitively, assuming that the tangle threads or core sketch, made in the first place, provide appropriate contextual conditions for the performing of construction. So, the spider would use its own threads to support its web. Although it may have some facilitative influence, the presence of the tangle or core sketch threads does not seem sufficient, since some spiders sometimes spent weeks with a dense tangle, without building; others, promptly made a web, bypassing the tangle stage. This opens the possibility that the improvement from H11 to H1End resulted from a process of acquiring experience. Studies that extend the recording duration of webs in a horizontal position, systematically collecting data on H2, H3, H4 ....Hn could say to what extent this hypothesis is correct.
The sharp improvement in the construction quality from H11 to H21 and the decrease in latency for the web construction are the most relevant results of this research, by suggesting the influence of a past experience or evidence of learning. However, three non-cognitive explanations need to be examined: (1) the explanation for previous casual threads; (2) the explanation for the decrease in stress; (3) the explanation for the age of spiders. The first explanation is not valid, as it assumes the use of previous tangles as facilitators. Unlike in the case of H12 (or H1End), in H21, the spider constructs the web in a box clear of threads. Regarding the explanation of emotional factors, as seen in the above (VI x V2), it is based on the assumption that environmental changes and/or stimulation due to the capture and transportation generate disorder that somehow inhibits or disrupts the web construction. One might imagine that there was a habituation of emotional response in H1, thus, the spider would arrive in H2 more relaxed and ready to build. However, they would need to remember the context in which the initial habituation occurred, in order for no recuperation or de-habituation occur. This assumes a factor of past experience specific to the horizontal box condition. The third hypothesis, in respect of age, the spiders would build better webs in H21 as they were older than when they constructed in H11. Unpublished preliminary findings indicate instead that age should bring a worsening in the ability to construct. It was observed that younger, immature spiders, build regular horizontal webs faster than older spiders (Ades, 1973). The spiders, at the end of the experiment (when weaving H21) were already fairly old, five having died among them before V4. It is not plausible to assume that they were better prepared to build, at this stage of their life.
Spiders placed in a situation that is different from usual, and that offers poor supports compared with supports that the spider would usually find in nature, have a performance decrease that is significantly reverted throughout the experiment in this situation. The same spiders when are placed a second time in contact with a poor bracket, there is strong evidence that it is a process of taking advantage of the prior experience. But what is this experience? There was no recording of the spider's constructional behavior in a horizontal situation that could be compared to vertical records and that could provide clues about exactly which parts of the motor sequence were affected by environmental factors. However, it is possible to propose a hypothesis about this, from the examination of completed webs, or webs of more or less random structure, which the spiders often weave. The hypothesis must take into account not only the sequential construction improvement, but also of its two aspects which, apparently, seem to contradict an interpretation in terms of learning: the relatively sudden appearance of a web structure (jump), after random or nearly random forms and a return to less functional forms of construction (regression).
With respect to the jump, the classic definition of learning is gradual; it presupposes an acquisition of skills or information through successive and partial gains. It is therefore surprising for those who observe webs made successively horizontally, the sudden appearance of a whole structure, even with imperfections, after confused spinning. Might one think that the learning of a sensorimotor ability would be used in all phases of the construction? One can assume that learning has to do with the early stages of construction, which are the placement of threads on the frame with peripheral side moorings, radials on the inside. Given a reasonable initial framework, the following steps can be elicited more securely, even if they are defective. This does not eliminate the possibility of motor learning that has to do with the following steps (placement of the viscous spiral, for example), but, as we will show later, we do not have enough data to do so.
The process of web quality regression surprises as much as the jump, because the webs are on a continuous qualitative growth and suddenly the spider regresses and constructs a web with lesser quality than the last. One interpretation of the initial steps of learning also seems to give an account for these regressions. The trial and error in the initial construction, when not providing the adequate framework, is impaired, as well as the rest of the construction, whose quality decays as abruptly as it had improved. The hypothesis of an adaptive modification in the first steps of construction receives partial confirmation in the following point: when a spider has a perfect web (built vertically) as a starting point for a horizontal construction, it usually produces very good quality webs.
Constructing a web in modified conditions causes the spider to modify some of its behavioral routines in the construction of a new web, having to resort to different guiding "signs" (Vollrath, 1986; Vollrath, 1987; Biere & Deu, 1981; Suter, 1984). The choice of a site for the construction actually occurs and is the first step towards the construction of a web in nature (Ades, 1973). Several times, in horizontal conditions, the spiders built structures in one place and the next day they built in another, as if they were undecided as to the established coordinates set out in the first construction, until they find the ideal point to weave their mesh, which often appears suddenly. It seems that the Argiope argentata spider, when in contact with modified environmental situations, uses selective processes for acquiring environmental information and thus performs motor skills in which to obtain a positive functional answer to its needs. According to Gould and Marler (1987) "An animal is innately equipped to recognize when it must learn, so that stimuli directs its attention, how to store new information and how to retrieve it in the future" (p. 73).
The most relevant comparison is the one that can be made between H11 and H21; the constructions were made in identical external circumstances (box in a horizontal position, absence of web treads that could serve as guides). Any difference between webs is most likely the result of a difference in prior experience. The hypothesis of learning or effects of past experience in the case of the web construction should not sound like an isolated hypothesis.
Witt (1968), when studying the effects of increasing weight on the web structure of Araneus diadematus, observes that the spiders thicken the provisional spiral threads; however, with the decrease of weight, Araneus does not return immediately to its original web standards. The author proposes two explanations for the problem: "this could be explained by some damage during the weight reduction or by a process of 'learning' that does not immediately fade" (p. 50). This second hypothesis proposed by Witt (1968) suggests possible effects of past experience, confirming the importance for more intense investigations of cognitive processes in spiders. These signs of memory and learning are reported in various aspects of construction when spiders are subjected to unusual experiences (Olton, 1979). Witt et al. (1977) suggest evidence of past experience in the web construction of Araneus diadematus - considering that the processes of construction are not so rigid as they seem - because when subjected to weightlessness in the Skylab project the authors reported: "...The ability of an invertebrate with rigid behavioral patterns like in web construction, which is relatively independent of experience in finding alternative ways to complete a perfect mesh to eat and increase its chances of survival, is possibly the most important finding from the Skylab research project records" (p. 123). Witt was greatly surprised (1977) when he encountered behaviors susceptible to experience. This fact highlights the importance of being aware of the problems that the spiders are made to solve. The animal seems sensitive to certain conditions, being able to change their behavior in order to survive. It is interesting to see what kinds of cognitive processes are linked to these modified behaviors.
Eberhard (1988a), when studying the provisional spiral position in Leucauge mariana, reports the existence of memory of distances and directions in the construction of its web: "The influence of several factors are probably estimated in the basic clues from the position of provisional spiral lines and memories of distances in the determination of space of this spiral" (p. 64). "... the spider recognizes each radial individually and remembers its relative position to the others" (p. 65). There is an intrinsic adaptive function to the memory process; instinct and learning are elements that coexist within the same performance (Olton, 1979; Sherry & Schacter, 1987). "Today there is little objection to the idea that learning and memory play adaptive roles, that is, they make the animal more efficient, in certain circumstances, and probably contribute to its reproductive success" (Ades, 1991b, p. 32).
In studies on gravitational orientation, the use of information integration is more evident. Ades (1989), observed spatial mnemonic recording in Argiope argentata. The spiders were attracted to a point (p1) at the edge of the web and the vibratory source was moved so that the spider would move from the initial to the second point (p2), the spider would have two possible paths back to the center: (1) it would return through p1. or (2) it would go straight to the center. In 80% of the cases, the spiders returned straight to the center and 20% were observed turning back. "It is not possible to totally eliminate the hypothesis that the spiders, in the second case, were using local geometric clues, but the speed of their return suggests that there was a process of stimuli integration present" (p. 32). The observation of the author reports a mnemonic processes of the animal in which the memory composes this spatial orientation in "path integration". Systemic flexibility in Argiope contributes to its performance in the face of environmental changes. Adaptive instruments, such as memory and learning, are integrated and contribute on a daily basis, in solving environmental problems (Ades, 1987; Gould & Marler, 1987; Hinde, 1970).
References
Ades, C. (1973). A teia e a caca da aranha Argiope argentata. Doctoral dissertation, Institute of psychology, University of São Paulo, São Paulo. [ Links ]
Ades, C. (1982). Experiência passada e integração do comportamento em invertebrados. Science and Culture, 35 (2), 137-147. [ Links ]
Ades, C. & Rossetto, 1. A. Jr. (1983). Espaço e programa de construção da teia numa aranha orbitela: resultados preliminares. Em livro de resumos I Encontro Paulista de Etologia (p.247), UNESP, Jaboticabal, SP. [ Links ]
Ades, C. (1989). O que aprendem e de que se lembram as aranhas. Em Ades, C. (Ed.) Etologia: de animais e de homens, (pp.1-4). São Paulo, SP: EDUSP. [ Links ]
Ades, C. (1991a). Memória e instinto no comportamento de predacão da aranha Argiope argentata. Tese de livre docência, Instituto de Psicologia, University of São Paulo, São Paulo. [ Links ]
Ades, C. (1991b). O controle mnêmico como determinante dos comportamentos típicos da espécie. Biotemas, l, 21-36. [ Links ]
Ades, C. & Cunha, S. S. (1991). O ajustamento da teia da aranha Argiope argentata a condições gravitacionais modificadas (p.221). Em livro de resumos VIII Encontro de Etologia. [ Links ]
Ades, C. (1987). Notas para uma análise psicoetológica da aprendizagem. Boletim de Psicologia, 37(86), 24-35. [ Links ]
Ades, C., Hardt, A., Sasaki, R. (1986). A teia prévia como guia durante a reconstrução, na aranha Argiope argentata (p. 28). Em livro de resumos XIII Congresso Brasileiro de Zoologia, Cuiabá [ Links ].
Benforado, L. & Kistler, K. H. (1973). Growth of the orb weaver, Agustin diadematus, and correlation with web measurements. Psyche-A Journal of Entomology, 80, 90-100. [ Links ]
Biere, M. & Uetz, G. W. (1981). Web orientation in the spider Micratena gracilis (Araneae: Araneidae). Ecology, 62 (2), 336-344. [ Links ]
Bouton, M.E. (2007). Learning and behavior: The comtemporany synthesis. (Sinauer Associates, Inc) Sunderland, MA, USA. Pp. 482. [ Links ]
Braga, M. M. (1986). As variações de teias na vertical para teias na horizontal em Argiope argentata (p.45). Em resumos IV Encontro de Etologia. [ Links ]
Caramaschi, S. (1991). Luz e Temperatura como Fatores na Orientacão Postural da aranha Argiope argentata. Dissertação de mestrado apresentada ao Instituto de Psicologia, Universidade de São Paulo, São Paulo. [ Links ]
Christiansen, A., Baum, R. & Witt, P. (1962). Changes in spider webs brought about by mescaline, psilocybin and an increase in body weight. Journal of Pharmacology and Experimental Theory, 136, 31-37. [ Links ]
Comstock, H. (1948). The spider book. Ithaca, N.y: Comstock Publishing Co., Inc. [ Links ]
Eberhard, W. (1969). Computer simulation of orb-web construction. American Zoologist, 2, 229-238. [ Links ]
Eberhard, W. G. (1986). Hub construction by Leucauge mariana, (Araneae: Araneidae). Bulletin British Arachnology Society, 7(4), 128-13l. [ Links ]
Eberhard, W. G. (1988a). Memory of distances and directions moved the cues during temporary spiral construction in the spider Leucauge mariana (Araneae: Araneídeo). Journal of Insect Behavior, 1, 51-66. [ Links ]
Eberhard, W. G. (1988b). Behavioral flexibility in orb web construction: effects of supplies in different silk glands and spider size and weight. Journal of Arachnology, 16, 295-302. [ Links ]
Edrich, W. (1977). Interation of light and gravity in the orientation of the waggle dance of honey bees. Animal Behavior, 25, 342-363. [ Links ]
Enders, F. (1976). Effects of prey capture, web destruction and habitat physiognomy on web-site tenacity of Argiope spiders (Araneidae). The Joumal of Arachnology, 1, 75-82. [ Links ]
Gould, J. L. & Marler, P. (1987). Learning by instinct. Scientific American, 256, 62-73. [ Links ]
Harmer, A. M. T. & Herberstein, M. E. 2009. Taking it to extremes: what drives extreme web elongation in Australian ladder web spiders (Araneidae: Telaprocera maudae)? Animal Behaviour, 78, 499-504. [ Links ]
Hinde, R. A. (1970). Animal behaviour. McGraw-Hill Kogakusha ed., Tokyo, Japan. [ Links ]
Hollis, K. L. (1984). Causes and function of animal learning processes. Marler, P. ed. (Berlin, Heldelberg, New York, Tokyo: Springer Verlag) (The biology of learning). [ Links ]
Jackson, R. R. (1974). Effects of d-amphetamine sulfate and diazepam on thread connection fine structure in the spider's web. Journal of Arachnology, 2.37-4 l. [ Links ]
Jander, R. (1975). Ecological aspects of spatial orientation. Annual Review of Ecology and Systematics, 6, 171-189. [ Links ]
Leborgne, R. & Pasquet, A. (1987). Influences of aggregative behaviour on occupation in the spider Zygiella x-notata (Clerck). Behaviour and Sociobiology, 20, 203-208. [ Links ]
LeGuelte, L. (1966). Structure de la toile de Zygiella-x-notata Cl. (Araignees, Argiopidae) et facteurs qui regissent le comportement de l`Araignée pendente la construction de la toile. These, publ. Université de Nancy. [ Links ]
LeGuelte, L. & Ramousse, R. (1979). Effect of environment on the rhythm of web building behaviour in Agustin diadematus CL. Biology of Behaviour, 4, 289-302. [ Links ]
Maciejewski, W. (2010). An analysis of the orientation of an orb-web spider. Journal of Theoretical Biology, 265, 604-608. [ Links ]
Masters, W. M. & Moffat, A. J. M. (1983). A funcional explanation of top-bottom asymmetry in vertical orb webs. Animal Behaviour, 31, 1043-1046. [ Links ]
Nakata, K. (2007). Prey detection without successful capture affects spider's orb-web building behaviour. Naturwissenschaften, 94, 853-857. [ Links ]
Nakata, K. (2009). To be or not To be conspicuous: the effects of prey availability and predator risk on spider's web decoration building. Animal Behaviour, 78, 1255-1260. [ Links ]
Nakata K. (2012) Plasticity in an extended phenotype and reversed up-down asymmetry of spider orb webs. Animal Behaviour, 83, 821-826. [ Links ]
Olton, D. S. (1979). Mazes, Maps, and Memory. American Psychologist, 34, 583-596. [ Links ]
Opell, B. D. (1987). Changes in web monitoring forces associated with reduction in the spider family Uloboridae. Canadian Zoology, 65, 1028-1034. [ Links ]
Reed, C. F. (1969). Cues in the web building process. American Zoologist, 9, 211-221. [ Links ]
Reed, C. F. & Witt, P. N. (1968). Progressive disturbance of spider web geometry caused by two sedative drugs. Physiology and Behavior, 3, 119-124. [ Links ]
Reed, C. F., Witt, P. N. & Scarboro, D. B. (1969). The orb web during the life of Argiope aurantia (Lucas). Developmental Psychobiology, 2, 120 -129. [ Links ]
Seyfarth, E. A., Hergenroder, R., Ebbes, H. & Barth, F.G. (1982). Iothetic Orientation of a Wandering Spider: compensation of detours estimates of goal distance. Behaviour Ecology Sociobiology, 11, 139-148. [ Links ]
Sherry, D.f. & Schacter, D. L. (1987). The evolution of multiple memory systems, 94, 4, 439-454. [ Links ]
Slep, R. (1952). On the plasticity of Instinct of a garden spider (Aranea . , L) Construction of a Cob web. Acta Biologiae Experimentalis, 16, (1): 5-24. [ Links ]
Suter R.B. (1984). Web tension and gravity the cues m spider orientation. Behaviour Ecology Sociobiology, 16, 31-36. [ Links ]
Thorpe, W. H. (1963). Learning and Instinct in Animals. 2 ed. Methuen, London. [ Links ]
Tilquin, A. (1942). La toile geométrique des araigneés. Presses Universitaires de France, Paris. [ Links ]
Tso, I.-D., Chiang, S.-Y. & Blackledge, T. A. (2007). Does the giant wood spider Nephila pilipes respond to prey variation by altering or silk web properties? Ethology, 113, 324-333. [ Links ]
Vollrath, F. (1986). Gravity as an orientation guide during construction in the orb-web spider Agustin diadematus (Araneae, Araneidae). Comparative Physiology .159, 275-280. [ Links ]
Vollrath, F. (1987). Altered geometry of webs in spiders with regenerated legs. Nature, 328, 247-248. [ Links ]
Vollrath, F. (1988). Spiral Orientation of Agustin diadematus orb webs built during vertical rotation. Comparative Physiology .162, 413-419. [ Links ]
Vollrath, F. & Mohren, W. (1985). Spiral geometry in the garden spider's orb web. Naturwissenschaften, 72, 666-667. [ Links ]
Witt, N. P. & Baum, R. (1960). Changes in orb webs of spiders during growth (Agustin diadematus Clerck and Neoscoría vertebrata Mccook). Behaviour, 12, 309-317. [ Links ]
Wiu, P. N. & Reed, C. F. (1965). Spider Web Building. Science, 149, 1190-1197. [ Links ]
Wiu, P. N., Reed, C. F. & Peakall, D. B. (1968). The Spiders Web: Regulatory Problems in Biology. Springer-Verlag Berlin, Heidelberg, New York. [ Links ]
Witt, P. N., Scarboro, D. B. & Daniels, R. (1977). Spider web-building in outer space: evaluation of records from Skylab experiment. Journal of Arachnology, 4.115 -124. [ Links ]
Received in September 30, 2012
Accepted in February 20, 2013
