










Serviços Personalizados
artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805
Rev. etol. vol.11 no.1 São Paulo 2012
Associative learning in anastrepha obliqua (diptera, tephritidae) wild females and males of different ages and nutritional states
Aprendizagem associativa em fêmeas e machos de Anastrepha obliqua (Diptera, Tephritidae) com diferentes idades e estados nutricionais
Carla Cresoni-Pereira; Luanda Medeiros-Santana; Fernando Sérgio Zucoloto*
Department of Biology, FFCLRP - University of São Paulo, Ribeirão Preto, SP, Brazil
ABSTRACT
Adult Anastrepha obliqua fruit-flies need protein sources to promote sexual maturation and egg production. Learning can be a mechanism to improve the foraging behavior to acquire this nutrient. Learning could be defined as the acquisition of a new representation of neuronal information, and it can only be indirectly observed through its potential effect on behavior. In the present study, the occurrence or not of associative learning in A. obliqua females and males of different ages and nutritional states concerning sucrose (carbohydrate) and brewer´s yeast (protein source) was observed. Associative learning for sucrose was demonstrated in young and old females and males only when deprived of food for 2 days after the experience period. As concerns yeast, the associative learning was demonstrated in deprived young and old females only.
Keywords: associative learning, fruit flies, protein, sucrose
RESUMO
Adultos de moscas-das-frutas, Anastrepha obliqua, necessitam de fontes proteicas para promover a maturação sexual e produção de ovos. A aprendizagem pode ser um mecanismo para melhorar o comportamento de forrageamento na aquisição deste nutriente. Dentre as diferentes definições de aprendizagem uma delas a define como a aquisição de uma nova representação de informação neuronal e pode ser indiretamente observada através de seu efeito potencial no comportamento. No presente estudo, a ocorrência ou não de aprendizagem em fêmeas e machos de A. obliqua de diferentes idades e estados nutricionais foram observadas em relação à sacarose (carboidrato) e lêvedo de cerveja (fonte de proteína). A aprendizagem associativa para sacarose foi demonstrada em fêmeas e machos jovens e velhos somente quando privados de alimento por dois dias depois do período de experiência. Em relação ao lêvedo, a aprendizagem associativa foi demonstrada em fêmeas jovens e velhas, mas não nos machos.
Palavras-chave: aprendizagem associativa, mosca-das-frutas, sacarose, proteína
Adult Anastrepha obliqua fruit-flies need carbohydrate sources to survive and protein sources to promote sexual maturation and egg production (Aluja, 1994; Braga & Zucoloto, 1981). In Tephritidae, some mechanisms to acquire these nutrients were already investigated, among them the females associative learning (Cresoni-Pereira & Zucoloto, 2001; 2006a).
Several studies have shown that a variety of insects, including predators, polyphagous and medical importance insects, can learn associations with resources, odors and tastes (Bernays & Weiss, 1996; Cresoni-Pereira & Zucoloto, 2006a; Glinwood et al., 2011; Papaj & Prokopy, 1989; Simpson & White, 1990; Stireman, 2002; Tomberlin et al., 2006). In these cases, learning can result in changes concerning the feeding preferences when the taste of a certain food is associated to satisfactory post-ingestive effects; consequently, taste acquires the neural interpretation of adequate food (Bernays & Weiss, 1996; Burke & Waddell, 2011; Fujita & Tanimura, 2011). Association learned as concerns partners for copulation in Drosophila melanogaster (Dukas, Clark & Abbott, 2006), and the choice of hosts for oviposition through colors learned by some Tephritidae genders (Papaj & Prokopy, 1989) and by Pieris brassicae (Smallegange, Everaarts & Van Loon, 2006) were already shown and can cause fitness intensification. In Blatella germanica, the importance of the spatial distribution, the persistence of the alimentary resources, and the associative learning in the choice of foraging tactics were shown (Durier & Rivault, 2001).
Learning could be defined as the acquisition of new representations of neuronal information, as spatial configurations, spatial information, associations among stimuli, environmental factors and motor patterns (Dukas, 2008). Learning can only be indirectly observed through its potential effect on behavior (Dukas, 2008), and therefore can often be confused with other behavioral aspects.
Some recently discussed questions as concerns to learning by insects refer to the adaptive value of learning in at least two aspects, because associative learning can be more beneficial for the insects than the innate behaviors, and because the insects must learn during short lifetimes (Dukas, 1999; Meri & Kaweki, 2005, Dukas, 2008).The hypothesis for these questions lies in the increased fitness during the insect lifetime, considering the short foraging time provided by the associations learned in relation to the resources.
The aim of this study was to observe the occurrence of associative learning in A. obliqua females and males of different ages and nutritional states concerning sucrose (carbohydrate) and brewer´s yeast (protein source). The relevance of the associated factors (nutrients) is discussed focusing costs and benefits of learning.
Method
Wild A. obliqua adults were obtained in Spondias venulosa, Anacardiaceae, infested fruits collected in the Bonfim Paulista district, Ribeirão Preto, São Paulo (21º17´75´´S, 47º81´2´´W).
The infested fruits were collected and placed in plastic boxes (60x40x10 cm) containing sand (approximately 5 cm). When the pulps were completely dry, the fruits were removed from the box and the pulps were separated from the sand with a sieve, washed with running water and placed in acrylic boxes (11x11x3 cm) containing sterilized sand.
The boxes containing the pupae and the experimental acrylic boxes were maintained in a wooden muffle with glass lid at 29 ± 1ºC on average, 70 to 80% relative humidity and 12 hours light/12 hours dark photoperiod, using 400 lux fluorescent lamps.
The experimental acrylic boxes (11x11x3 cm) had three lateral orifices (1 cm diameter) to offer diet and water. The water was offered in small cotton tapped test tubes. The solid diets were offered as pieces attached to a pin stuck in a cork.
When emerging, the flies (15 females and 15 males per treatment; 1 box = 1 replicate) were randomly chosen and individualized in the acrylic boxes for the experiment.
The diets were placed in Erlenmeyers (200 ml), autoclaved, distributed in (50x12 mm) Petri dishes, and stored under refrigeration (10ºC) for posterior use.
Six different diets were offered as follows:
Y = yeast diet (Mãe Terra, São Paulo, Brazil) (6.5 g)
YQS = yeast diet (6.5 g) + quinine sulphate (Quimibrás, Rio de Janeiro, Brazil) (25 mg)
S = sucrose diet (Synth, Diadema, Brazil) (11.0g)
SQS = sucrose diet (11.0 g) + quinine sulphate (25 mg)
A = agar diet (Vetec, São Paulo, Brazil) (2.5g)
AQS = agar diet (2.5g) + quinine sulphate (25 mg)
Diets Y, YQS, S and SQS contained also agar 2.5 g (Vetec, São Paulo, Brazil), distilled water 100 ml, and Nipagin 1.5 ml (Merck®).
The daily ingestion measurements were obtained through the formula I = TDM-RDM (Cangussu & Zucoloto, 1995), where I = ingestion (mg/day), TDM = total dry matter (total weight of the diet placed in boxes without flies, maintained under the same experimental conditions to control evaporation), RDM = remaining dry matter (total weight of the diet placed in the experimental box).
The diets withdrawn from the boxes after 24 hours were further dried in a muffle for 24 hours at 80 ± 1ºC and then weighted to calculate ingestion.
Groups
Females and males were tested with regard to learning in relation to sucrose and brewer´s yeast. The tests followed the schedule presented in Table I. The nutrient tested was sucrose or brewer's yeast for different classes of flies (young or old flies, deprived or non-deprived ones). In the period of food deprivation flies were not fed, except group G, that received only sucrose as food in this period. In the experience period flies fed the diet described in the third column (diet consumed during the experience period). The "test of choice" describes the day in which the tests of choice of food was conducted. In this test, two diets were given to flies, as described in Table 1 for 24 hours. After this period, the diet was weighed (as described above) to determine the amount of food ingested by each group.
There was not a group of sucrose deprived old flies simultaneously to the yeast deprived ones (groups G, deprivation 1-50 days), because the flies do not survive more than 2 days without a carbohydrate source. The group G did not received only yeast diet during deprivation period.
Therefore, for the sucrose tests, another group was formed not only with young flies but also with old ones (groups B and D, respectively), in which, after the experience period, there was a two-day period of food deprivation before the choice.
The sucrose tested groups (A, B, C and D) were not fed with yeast in any of the tests. The yeast tested groups (E, F and G) received also sucrose diet during the tests because they do not survive without a sugar source.
The results were analyzed by the Wilcoxon test with 5% significance level. The statistic tests were made using the SigmaStat for Windows, version 2.03, Jandel Corporation.
Results
All groups fed with the diet composed by the tested nutrient without quinine sulphate during the experience period (all groups from A to G, 1 and 2, Table I) did not show significant differences among the offered diets in the day of choice. This result has shown that the nutrient by itself did not influence decisively the choices in any of the groups, independently of age, sex and nutritional state.
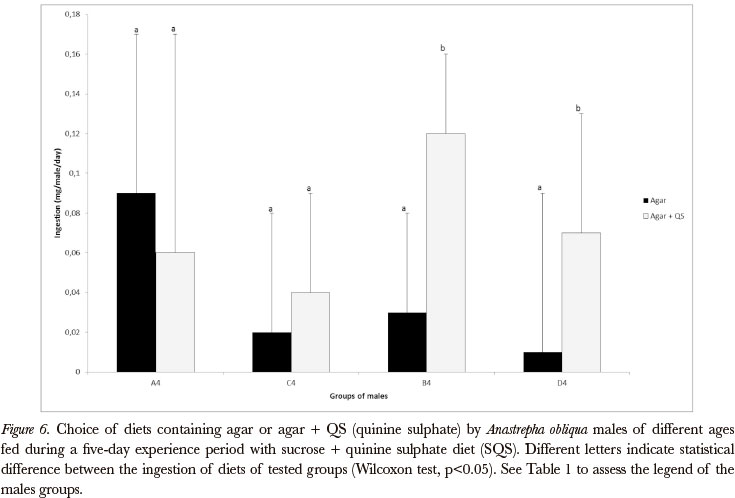
When the experience period diet contained quinine sulphate (all groups from A to G, 3 and 4, Table I), associative learning for sucrose was demonstrated in young and old females and males only when deprived of food for 2 days after the experience period (before the choice test) (Figures 1 and 2, and 5 and 6). When the choice test was carried out immediately after the experience period, no association between the experience period diet and the choice diets was observed.
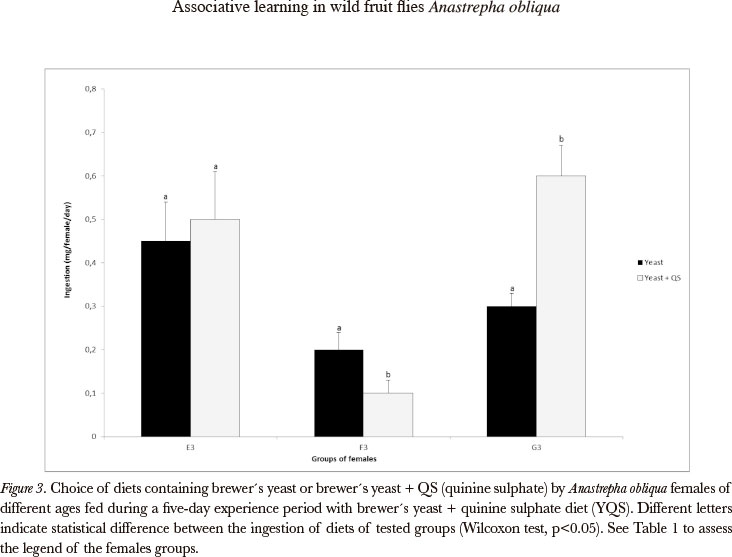
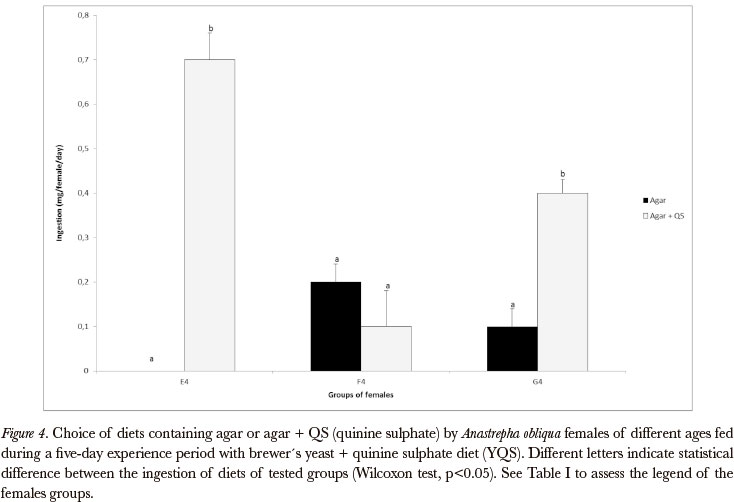
As concerns yeast, the associative learning was demonstrated in deprived young and old females (Figures 3 and 4). The males did not show associative learning in relation to yeast in any age or nutritional state (yeast deprived or non-deprived) (Figures 7 and 8), as well as the non-deprived females (Figures 3 and 4).
It is important to observe that, when the choice test diets had nutritive value, learning was not always evident as in the cases when the choice diets were constituted by agar.
Discussion
Previous studies have shown that quinine sulphate is not phago-stimulant for A. obliqua females (Cresoni-Pereira & Zucoloto, 2006a). In this study, all groups fed during the experience period with the test diet containing the tested nutrient without quinine sulphate did not show significant differences in the day of choice as compared to the other offered diets. This fact may indicate that the results from other groups are not due to quinine sulphate adverse effects (a novelty effect, for instance). Dukas (2008) alerts that care must be taken in the experimental design to guarantee that the stimulus presentation and the nutritional or environmental status of the individuals do not generate behavioral patterns that may be interpreted as learning not being so. In addition, small sample sizes may generate wrong conclusions about the ability for individual learning and inter and intraspecific differences (Thornton & Lukas, 2012).
Learning can be very specific as concerns relevant stimuli for the insect fitness. The parasitoid reproductive fitness, for instance, depends on its ability to find nectar or honeydew, since sugar is a pre-requisite for survival and reproduction (Winkler et al., 2006). Associative learning is one of the means the parasitoids can optimize their reproductive success (Wäckers et al., 2006).
The ability to learn seems to be related to the insect nutritional status, sex, rearing condition and age (Thornton & Lukas, 2012). In this study, learning was evident for yeast, which is a protein source, and proteins are, admittedly, a sparse resource in nature (Bateman, 1972). For A. obliqua females, proteins are very important for the youngsters as concerns sexual maturation and for older females when they are deprived of this nutrient during their lives, since the females of this species are able to reproduce late in life when they have been deprived of protein in the beginning of their lives (Cresoni-Pereira, personal observation). Insects are considered organisms of short lifetime when compared to mammals, for instance. However, if the insect is able to reproduce late in life, as is the case for A. obliqua, due to the lack of a nutrient during the young phase, any learned aspect that can revert that reproductive failure is biologically relevant for the species. In this case, capability to learn is highly beneficial as any other strategy to search and select adequate foods.
Males did not evidence learning in relation to yeast. Apparently, the larval protein supply is determinant for the reproductive performance of the adult male, since it would influence important aspects as size (Blay & Yuval, 1999) and the production and release of pheromone (Ferro & Zucoloto, 1989). Regarding proteins in feeding adults, Shelly & Kennelly (2002), using C. capitata, have shown that protein non-deprived males obtained significantly more copulation than the protein deprived males, either wild or bred in the laboratory. Drew (1987) observed that Bactrocera tryoni males do not need a protein source to produce sperm, however they need one to maintain production along their lives.
As concerns A. obliqua males, Cresoni-Pereira & Zucoloto (2006b) observed that males maintained in a sucrose diet had reduced longevity when compared to males fed with sucrose and protein. Therefore, the A. obliqua males reproductive performance can be improved by the ingestion of a protein source but is not impaired by its absence. Maybe that is why there is not a learned mechanism that can improve the males efficiency to search and select this nutrient.
Associative learning for sucrose was demonstrated in young and old males and females. Similar results were obtained for Culex quinquefasciatus, in which case males and females are able to learn the association of an odor with a source of sugar (males and females) and blood (females) (Tomberlin, et al., 2006), and Microplitis croceipes where males responded as well as females to sucrose associated odor (Takasu, Rains & Lewis, 2007). In relation to sucrose, learning was evident only in cases of deprivation. Some studies show that hunger is necessary for more robust olfactory learning (Fujita & Tanimura, 2011). Carbohydrates are abundant in nature, consequently the development of learned responses for this resource may not be advantageous, except in deprivation cases. As the learned responses involve a neural cost, responses for abundant nutrients may represent an unnecessary cost.
A question for the future is to know the duration of the learned responses for an alimentary resource. In this case, deprivation immediately followed the experiment, therefore the response for the quinine sulphate was demonstrated. If that association is long lasting and translates a recognition response of the resource in future deprivation periods, that learned association can be highly advantageous for the insect as concerns increased foraging efficiency in adverse environments.
An important factor is the relevance of the learned stimulus. The classic conditioning theory anticipates that a high quality reward would increase the probability of learning in relation to a low quality reward (Hutt, 1954 apud Wäckers et al., 2006). The protein source undoubtedly is a high quality reward for young A. obliqua females and for protein deprived old ones. However, as concerns sucrose, though it is important for survival, it can be substituted by other highly nutritive carbohydrates easily found in nature (Fontellas & Zucoloto, 1999).
According to Papaj & Prokopy (1989), one of the attributes in order that a behavior is considered learning is its reversibility in the absence of a repeated stimulus, that is, the behavioral change that accompanies the experiment diminishes in the absence of a continuous experience of the same kind. This reversibility was demonstrated by Cresoni-Pereira & Zucoloto (2006a) in A. obliqua females. In the above mentioned study, the flies have shown association in the first day of the choice test, giving preference to the quinine sulphate diet that was the compound associated to yeast in the experience period. In the second day, as agar lacks nutritive value, the females directed their preference to a diet without quinine sulphate but also containing agar, and in the third day they ingested both diets indiscriminately.
This situation, registered by Cresoni-Pereira & Zucoloto (2006a), does not indicate lack of persistence as concerns the learned behavior; it indicates that the behavior ceased to be manifested in the absence of a positive post-ingestive response. Considering these data, it is not possible to affirm that the behavior could not be reactivated in a future situation with positive post-ingestive feedback.
Wäckers et al. (2006), working with the M. croceipes parasitoid, found that the gustatory responses, more than the post-ingestive feedback, act as a non-conditioned stimulus to learn odors related to phagostimulant carbohydrates. They also tested non-phagostimulant carbohydrates, however of similar nutritional value, that elicited weak learned responses, and this reinforced the hypothesis that taste is important and the post-ingestive feedback is not. These results do not agree with previous results found for A. obliqua (Cresoni-Pereira & Zucoloto, 2006a). However, Drosophila flies can learn the nutritional value of a non-sweet sugar when it is associated with an odor, and they can discriminate between sugars using a post-ingestive evaluation system, showing that the palatability and nutritional value contribute to form a more effective learning (Fujita & Tanimura, 2011; Burke & Waddell, 2011).
Aminoacids have different pre- and post-ingestive effects on behavior, and the nutritional and neuromodulating functions are primarily post-ingestive (Kim & Smith, 2000). The post-ingestive effects have a course of minutes or hours. Several insects have sensorial receptors sensitive to aminoacids (Simpson et al., 1991). Therefore, aminoacid detection and the responses could be oriented by sensorial pre-ingestive inputs that have a very short course of seconds (Kim & Smith, 2000).
Facility and readiness to form memory may vary among species and among individuals of the same species, depending on several factors as the environmental variability and the species longevity (Smid & Vet, 2006). The only influence of the apparent age on this study results was related to the reproductive capability. No other aspects resulting from aging as retarded responses, for instance, can be inferred here.
Results similar to those found for A. obliqua in this study were found for omnivorous Periplaneta americana cockroaches (Gadd & Raubenheimer, 2000) and Schistocerca gregaria (Simpson & White, 1990) that do not learn to associate non-alimentary odors to sucrose, however they learn the association with proteins showing that learning can be nutrient-specific.
Future studies must concentrate in the learned responses duration and in the type of resources that trigger the responses in order that one can establish more precisely the adaptive value of the insect capacity in learning to associate the resources to conditioned stimuli.
Acknowledgements
We thank Laércio Massocato for laboratory assistance. Financial support: FAPESP (Proc. 02/1976-7) and CAPES.
References
Aluja, M. (1994). Bionomics and management of Anastrepha. Annual Review of Entomology, 39, 155-178. [ Links ]
Bateman, M. A. (1972). The ecology of fruit flies. Annual Review of Entomology, 17, 493-518. [ Links ]
Bernays, E. A. & Weiss, M. R. (1996). Induced food preferences in caterpillars: the need to identify mechanisms. Entomologia Experimentalis et Applicata, 78, 1-8. [ Links ]
Blay, S. & Yuval, B. (1999). Oviposition and fertility in the Mediterranean fruit fly (Diptera, Tephritidae): effects of male and female body size and the availability of sperm. Annals of the Entomological Society of America, 92, 278-284. [ Links ]
Braga, M. A. S. & Zucoloto, F. S. (1981). Estudos Sobre a Melhor Concentração de Aminoácidos para Moscas Adultas de Anastrepha obliqua (Diptera, Tephritidae). Revista Brasileira de Biologia, 41, 75-79. [ Links ]
Burke, C.J. & Waddell, S. (2011). Remembering nutrient quality of sugar in Drosophila. Current Biology, 21, 746-750. [ Links ]
Cangussu, J.A. & Zucoloto, F.S. (1995). Self-selection and perception threshold in adult females of Ceratitis capitata (Diptera, Tephritidae). Journal of Insect Physiology, 41, 223-227. [ Links ]
Cresoni-Pereira, C. & Zucoloto, F.S. (2001). Dietary self-selection and discrimination threshold in wild Anastrepha obliqua (Diptera, Tephritidae). Journal of Insect Physiology, 47, 1127-1132. [ Links ]
Cresoni-Pereira, C. & Zucoloto, F.S. (2006a). Associative learning in wild Anastrepha obliqua females (Diptera, Tephritidae) related to a protein source. Iheringia, série Zoologia, 96, 53-56. [ Links ]
Cresoni-Pereira, C. & Zucoloto, F.S. (2006b). Influence of male nutritional conditions on the performance and alimentary selection of wild females Anastrepha obliqua (Macquart) (Diptera, Tephritidae). Revista Brasileira de Entomologia, 50, 287-292. [ Links ]
Drew, R.A.I. (1987). Behavioral strategies of fruit flies of the genus Dacus (Diptera, Tephritidae) significant in mating and host-plant relationship. Bulletin of Entomological Research, 77, 73-81. [ Links ]
Dukas, R. (1999). Costs of memory: ideas and predictions. Journal of Theoretical Biology, 197, 41-50. [ Links ]
Dukas, R., Clark, C. W. & Abbott, K. (2006). Courtship strategies of male insects: when is learning advantageous? Animal Behaviour, 72, 1395-1404. [ Links ]
Dukas, R. (2008). Evolutionary biology of insect learning. Annual Review of Entomology, 53, 145-160. [ Links ]
Durier, V. & Rivault, C. (2001). Effects of spatial knowledge and feeding experience on foraging choices in German cockroaches. Animal Behaviour, 62, 681-688. [ Links ]
Ferro, M. I. T. & Zucoloto, F.S. (1989). Influência da nutrição protéica no desenvolvimento da glândula salivar de machos de Anastrepha obliqua. Científica, 17, 1-5. [ Links ]
Fontellas, T.M.L. & Zucoloto, F.S. (1999). Nutritive value of diets with different carbohidrates for adult Anastrepha obliqua (Macquart) (Diptera, Tephritidae). Revista Brasileira de Zoologia, 16, 1135-1147. [ Links ]
Fujita, M. & Tanimura, T. (2011). Drosophila evaluates and learns the nutritional value of sugars. Current Biology, 21, 751-755. [ Links ]
Gadd, C. A. & Raubenheimer, D. (2000). Nutrient-specific learning in an omnivorous insect: the american cockroach Periplaneta americana L. learns to associate dietary protein with the odors citral and carvone. Journal of Insect Behavior, 13, 851-864. [ Links ]
Glinwood, R.; Ahmed, E.; Qvarfordt, E. & Ninkovic, V. (2011). Olfactory learning of plant genotypes by a polyphagous insect predator. Oecologia, 166, 637-647. [ Links ]
Kim, Y. S. & Smith, B. H. (2000). Effect of an amino acid on feeding preferences and learning behavior in the honey bee, Apis mellifera. Journal of Insect Physiology, 46, 793-801. [ Links ]
Meri, F. & Kaweki, T. J. (2004). An operating cost of learning in Drosophila melanogaster. Animal Behaviour, 68, 589-598. [ Links ]
Papaj, D. R. & Prokopy, R. J. (1989). Ecological and evolutionary aspects of learning in phytophagous insects. Annual Review of Entomology, 34, 315-350. [ Links ]
Shelly, T.E. & Kenelly, S. (2002). Influence of diet on male mating success and longevity and female remating in the Mediterranean fruit fly (Diptera, Tephritidae) under laboratory conditions. Florida Entomologist, 85, 572-579. [ Links ]
Simpson, S. J. & White, P. R. (1990). Associative learning and locust feeding: evidence for a 'learned hunger' for protein. Animal Behaviour, 40, 506-513. [ Links ]
Simpson, S. J., James, S., Simmonds, M. S. J. & Blaney, W. M. (1991). Variation in chemosensitivity and the control of dietary selection behaviour in the locust. Appetite, 17, 141-154. [ Links ]
Smallegange, R. C., Everaarts, T. C. & Van Loon, J. J. A. (2006). Associative learning of visual and gustatory cues in the large cabbage white butterfly, Pieris brassicae. Animal Biology, 56, 157-172. [ Links ]
Smid, H. M. & Vet, L. E. M. (2006). Learning in insects: from behaviour to brain. Animal Biology, 56, 121-124. [ Links ]
Stireman, J. O., III. (2002). Learning in the generalist tachinid parasitoid Exorista mella Walker (Diptera: Tachinidae). Journal of Insect Behavior, 15, 689-706. [ Links ]
Takasu, K.; Rains, G.C. & Lewis, W.J. (2007). Comparison of detection ability of learned odors between males and females in the larval parasitoid Microplitis croceipes. Entomologia Experimentalis et Applicata, 122, 247-251. [ Links ]
Thornton, A. & Lukas, D. (2012). Individual variation in cognitive performance: developmental and evolutionary perspectives. Philosophical Transactions of the Royal Society B, 367, 2773-2783. [ Links ]
Tomberlin, J.K.; Rains, G.C.; Allan, S.A.; Sanford, M.R. & Lewis, W.J. (2006). Associative learning of odor with food- or blood-meal by Culex quinquefasciatus Say (Diptera: Culicidae). Naturwissenschaften, 93, 551-556. [ Links ]
Wäckers, F., Bonifay, C., Vet, L. & Lewis, J. (2006). Gustatory response and appetitive learning in Microplitis croceipes in relation to sugar type and concentration. Animal Biology, 56, 193-203. [ Links ]
Winkler, K.; Wäckers, F.L.; Bukovinszkine-Kiss, G.& Lenteren, Van J.C. (2006). Sugar resources are vital for Diadegma semiclausum fecundity under field conditions. Basic and Applied Ecology, 7, 133-140. [ Links ]
Received in October 29, 2012
Accepted in September 10, 2013
* Fernando Sérgio Zucoloto, Av. dos Bandeirantes, 3900. Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto - USP, Biology Department, Ribeirão Preto, SP, Brazil. CEP: 14040-901. Email address: zucoloto@ffclrp.usp.br

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}