










Serviços Personalizados
Journal

artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores
Compartilhar

 Permalink
PermalinkRevista de Etologia
versão impressa ISSN 1517-2805
Rev. etol. vol.12 no.1-2 São Paulo dez. 2013
Extra group copulation in Alouatta caraya (Primates, Atelidae) living on an island of urban forest from southeast São Paulo, Brazil
André Fernando de Oliveira FermoseliI, IV; Zelinda Maria Braga HiranoIII; Wagner Ferreira dos SantosI, II, *
IBiology Department, College of Philosophy, Sciences and Literature of Ribeirão Preto (FFCLRP), University of São Paulo (USP), Ribeirão Preto (SP), Brazil
IIInstituto de Neurociências e Comportamento (INEC), FFCLRP, USP, Ribeirão Preto (SP), Brazil
IIIUniversidade Regional de Blumenau (FURB), Blumenau (SC); Centro de Pesquisas Biológicas de Indaial (CEPESBI), Indaial (SC), Brazil
IVFaculdade Integrada Tiradentes (FITS), Maceio (AL), Brazil
ABSTRACT
Extra group copulations are rarely observed among the Alouatta genus, which lives in a fragmented area in Brazil. Here, we describe the second extra group copulation in A. caraya in literature, which was the first from Brazil. This happened in the southeast urban island forest, which measures 25 ha. In this forest there are two troops: one of them with thirteen howlers and the other with four. These howlers may be extinct in a few years since in the region there is only 3.89 % of native forests and high anthropic action due to agribusiness. We discuss the possible issues that led to the extra group copulation.
Keywords: Alouatta caraya; black and gold howler monkey; extra group copulation; urban forest; Brazil.
RESUMO
Cópulas extra grupo raramente são observados em animais do gênero Alouatta, que vivem em áreas fragmentadas do Brasil. Aqui, descrevemos a segunda cópula extra grupo em A. caraya na literatura, que foi a primeira do Brasil. Isso aconteceu na floresta ilha urbana sudeste, que mede 25 ha. Nesta floresta existem dois bandos: um deles com treze bugios e o outro com quatro. Estes bugios podem ser extintos em poucos anos, uma vez que na região há apenas 3,89% de florestas nativas e alta ação antrópica, devido ao agronegócio. Nós discutimos os possíveis aspectos que levaram à cópula extra grupo.
Palavras-chave: Alouatta caraya; bugio preto e bege; cópula extra grupo; floresta urbana; Brasil.
Introduction
In Alouatta social organization should be recognized through variable group sizes. Moreover, this variation may be due to characteristics of the species such as, sexual composition, organization kind, food resources and the occupied habitat (Chapman, 1989; Crockett, 1998; Chapman & Balcomb, 1998). Although there are many males in the group, the alpha usually monopolizes all reproductive females and all the other youngsters (Pope, 1990).
Because of monopolization, immigration of both sexes has been reported, resulting in new groups or acceptance of individuals from other groups (Neville et al., 1988; Mendes, 1989; Calegaro-Marques & Bicca-Marques, 1996). Nevertheless, extra group copulation (EGC) was only reported in A. pigra (Horwich, 1983), A. seniculus (Agoramoorthy & Hsu, 2000), A. guariba (Fialho & Setz, 2007) and A. caraya (Kowaleski et al., 2005; Kowaleski & Garber, 2010).
Migration of howlers is critical in Ribeirão Preto (SP, Brazil), because all troops studied live in small isolated fragments on farms or nearby cities. This prevents the usual junctions between couples of this genus creating intra group conflicts and short-term deaths and, henceforth, endogamy.
Here, we report the EGC in Alouatta caraya that occurred in southeast Brazil.
Methods
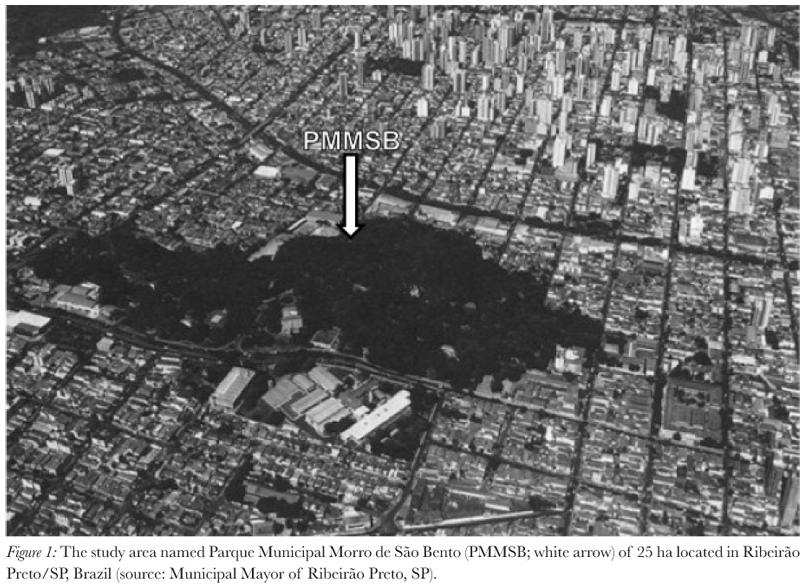
We studied black and gold howler monkey troops (A. caraya) that lived on the urban island forest from August 2006 to November 2008. This area is named as "Parque Municipal Morro de São Bento" (PMMSB) (21º10'17"S; 47º48'05"W; 25 ha), in Ribeirão Preto (São Paulo, Brazil). This place is an Environmental Protection Area (EPA), where the City Zoo is located, with a forest of 3 ha, and it is surrounded by houses, streets and stores (Figure 1).
In the PMMSB there are two groups of howlers, group 1 (G1) and group 2 (G2). G1 was composed by three adult males, one sub-adult male, five adult females, two juveniles and two infants. G2 was composed by two adult males and two adult females. Apparently, the alpha males have the same body size. Moreover, there are two solitary adult males living in the PMMSB.
The howlers were classified according to Calegaro-Marques and Bicca-Marques (1993) in: adult males (M), adult females (F), sub-adult males (SA), juveniles (J) and infants (I). All members were categorized and recognized according to their individual characteristics, such as, for example, hair coloration, body size and scars, which received specific codes. Thereby, the memberships were separated into the aforementioned classes (M, F, SA, J and I) and numbered, for example: the dominant male, or alpha male, received the M1 code; the other males received the M2 code and so on. They were later related with their original group as G1 or G2, for instance, the M1G1 (alpha adult male from G1), the F1G1 (dominant adult female from G1), and so on.
We used binoculars (Vanguard, 7x21x40 ZOOM, USA), a handheld computer (Palm One Zire 72, USA) and a video camera (JVC, Mini DV, Japan). The copulation was video recorded, and descriptive analysis was transcribed according to the video.
Results
Inter-group encounters that were frequently observed between G1 and G2 were always accompanied by long-term vocalization involving, generally, all males and females from the group. However, the inter-groups encounters were peaceful and without agonistic behavior.
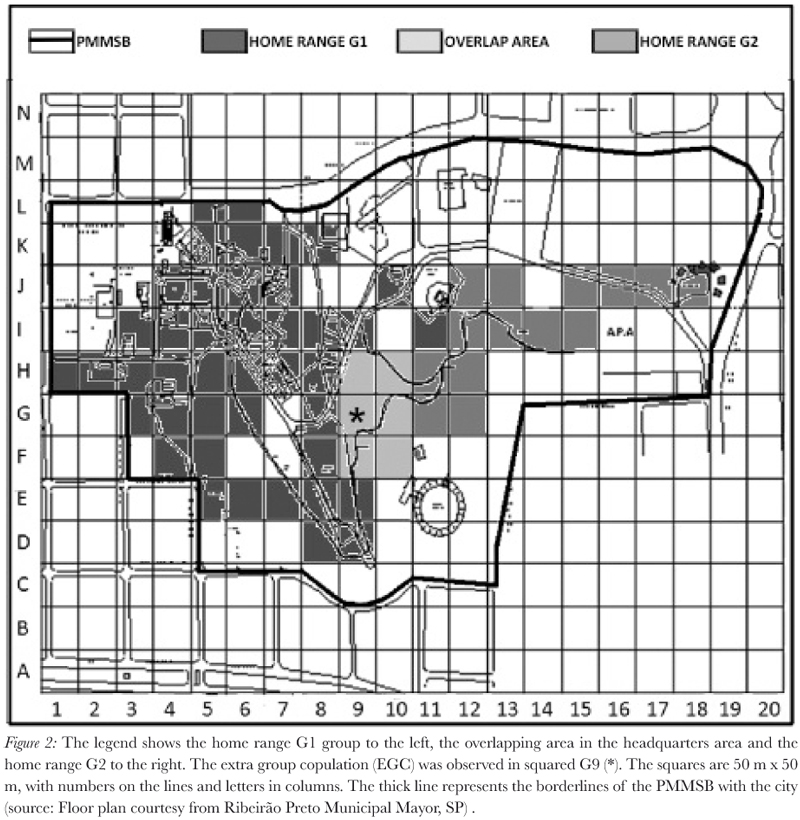
An inter-group encounter was observed on August 13, 2007, after locomotion led by the F1G1 (7: 32 a.m.). However, three males (M1G1, M2G1 and M3G1) exceeded this female near the overlapping area (Figure 2; squares F9-10, G9-10 and H9-10), which is used by both groups for feeding. The youngest adult male (M3G1) always remained ahead of other two males in the locomotion. The M1G2 kept observing the tree with adult males (M1G1, M2G1 and M3G1) and 6 min later, he approached them.
The M3G1 and J1G1 headed to the M1G2 and F1G2 while keeping a distance of approximately 10 m. The M3G1 and J1G1 realized this new approach and stopped on the same tree (Anadenanthera sp.) where the M1G2 and F1G2 were. The four individuals fed together on the same tree and no agonistic behavior was observed. The other howlers from G1 and G2 neither participated nor vocalized. Two other G1 individuals, the F3G1 and M2G1, were moving towards the tree. This made F1G2 leave; however, she remained close. The M1G1 drove towards the tree and threw the M1G2 out of the overlapping area, and it stayed behind the M1G1 (7:49 a.m.).
The F1G2 was chased by the J1G1 while avoiding the contact with him. This fact was observed several times until the F1G2 found itself surrounded by G1 individuals. The M1G1 moved towards the F1G2. This made the female run away, but the J1G1 and F5G1 (the youngest adult female) began to chase her. The F1G2 escaped and remained more than 100 m away from the other individuals.
The M1G2 observed the return of the J1G1 and F5G1 together. These individuals remained between the G1 and G2 groups. The J1G1 and F5G1 (8: 40 a.m.) were chased for 5 min by the M1G2 and M1G1 and the other howlers did not react. The chasing started again after 3 min, when the J1G1 escaped to another tree. However, the F5G1 moved to a tree branch without moving to other trees; the female was stuck.
The M1G2 approached the F5G1 slowly. This female rubbed on a branch with her genital area. The M1G2 conducted olfactory verification on the scent-marked branch and kept close to the female. EGC did occur after the F5G1 moved ahead of the M1G2 in squared G9. Vocalizations before and during copulation were not observed. The M1G2 performed 38 shots of copulation behavior.
It is likely that the M1G1 had not been seen copulating. The F5G1 remained with the M1G2, and returned to its group with short intervals (9: 18 a.m.). The M1G1 approached the F5G1 and conducted olfactory inspection of the genital area and made rhythmic short vocalizations. The M1G2 went towards the M1G1 and started to vocalize (9: 23 a.m.). The M1G1 vocalized with other group members. The vocalization had a 5-min duration. The inter-group encounter did not present any antagonism and ended peacefully (11: 43 a.m.), with each group returning to their home ranges.
Discussion
Extra group copulation has rarely been observed in Alouatta (Horwich, 1983; Agoramoorthy & Hsu, 2000; Fialho & Setz, 2007). All studies related EGC with inter-group encounters. Although vocalizations were observed in A. guariba at EGC (Fialho & Setz, 2007), vocalizations were not observed in A. pigra (Horwich, 1983), A. seniculus (Agoramoorthy & Hsu, 2000) nor in our study with A. caraya.
Genital inspections that preceded EGC were observed in both A. seniculus (Agoramoorthy & Hsu, 2000) and A. guariba (Fialho & Setz, 2007). Our observation showed that there was rubbing behavior by the F5G1 before EGC, and after that there was genital inspection performed by the M1G1 by the time the F5G1 went back to its group.
According to Smuts (1987), females are more responsible for courtship than males, and this fact was observed by Agoramoorthy & Hsu (2000), and Fialho & Setz (2007) in Alouatta. However, in our study, the female remained stuck and afterwards rubbing, and then she copulated with the M1G2.
After copulation, the M1G2 chased the F5G1 for a few minutes, but no agonistic behavior was observed. Males' chasing females after EGC were observed in other Alouatta species (Horwich, 1983; Agoramoorthy & Hsu, 2000; Fialho & Setz, 2007) and other neotropical primates as Callithrix jacchus (Digby, 1999).
Two hypotheses by Smuts (1987) were considered by Fialho & Setz (2007) in order to explain female mate choice: 1st, the search for genetically superior males and 2nd, the preference for non-familiar males. However, in this study the male involved in EGC was not larger than those belonging to the females' group. This first hypothesis is not supported by us. The second hypothesis is more consistent. However, according to Cabral et al. (2005), the G2 resulted from the G1 fission after observations of aggressive behavior occurring in 2003 and 2004. This fact, and the isolation of the PMMSB, does not permit migratory movements, preventing gene flow and leading to a decrease in genetic variability. Therefore, we believe that genetic gain of the F5G1 in the EGC is related to the events described above.
Black howlers live in cohesive social units when the benefits to the group, such as increasing foraging efficiency and predator avoidance, exceed the costs. The costs are exemplified, for instance, as an increase in intra-group competition for space and food, and competition for mating (Chapman & Chapman, 2000). In Alouatta females are monopolized by one male, an alpha male, forming group-like harems (Jones, 1983). Among adult females the socio-ecological relationships are defined according to causes and consequences of feeding or copulating competition, for example, home range resource distribution (Neville et al., 1988).
We believed that the F5G2 may have realized EGC firstly as consequence of a search for reproductive success and/or secondly as a consequence of submissive behavior, because the female was in a dangerous situation during the chase by the M1G2. This situation made the F5G1 rub and perform the copulate position. Although apparently the M1G2 had copulated with the F5G1, this does not assure the penetration or the fertilization of the female (confirmed due to no occurrence of births). We agree with the hypotheses related by other authors, as Smuts (1987) and Fialho & Setz (2007). However, given the non occurrence of births, we should not reject the possibility of submissive behavior that is important in avoiding possible aggression (Neville et al., 1988).
Kowalewski and collegues (2005) described EGC for first time with A. caraya in northern Argentina, and after described tens of extra-group copulations in this species (Kowalewski & Garber, 2010). This information was provided by Dr. Paul Garber, and the paper was published in the August issue of American Journal of Primatology (personal communication). Nevertheless, we described for the first time an EGC in A. caraya in Brazil. This fact is very important for howler monkeys, because studies that show social interaction between them will help to promote more efficient management plans for their survival in isolated areas. We stress that these monkeys will be at extinction risk in the Ribeirão Preto region.
It is interesting that we have as an example Zunino's study (Zunino et al., 2007). This author described that group sizes and group composition of howlers (A.caraya), living near the southern limit of its distribution in a semideciduous forest located in northern Argentina, suggest that the population has remained stable over the past 22 years (from 1982 to 2003). According this researcher, the density, number of groups, and individuals appear not to have been affected by fragmentation and logging, but crude density was low compared with other less-disturbed habitats. The status of the population remains uncertain owing to isolation, and because there are no protected areas to ensure its stability for the future (Zunino et al., 2007).
Acknowledgements
We are thankful to CAPEs (Brazil) for the PhD scholarship for Mr. André F. O. Fermoseli. We would like to thank Marisa de Carvalho, Alexandre Gouvêa and Pedro Favaretto for the permission to study at the PMMSB. The research complies with the current laws of the country in which it was conducted. We also thank teacher Abilio Borghi for the grammatical review of the English manuscript.
References
Agoramoorthy, G., Hsu, M. J. 2000. Extragroup copulation among wild red howler monkeys in Venezuela. Folia Primatologica. 71: 147–151. [ Links ]
Agoramoorthy, G., Rudran, R. 1993. Male dispersal among free-ranging red howler monkeys (Alouatta seniculus) in Venezuela. Folia Primatologica. 61: 92–96. [ Links ]
Cabral A, Pereira T, Fermoseli AFO, Santos WF. 2005. Fissão de um bando de bugios-pretos (Alouatta caraya) em mata urbana de Ribeirão Preto-SP". XI Congresso Brasileiro de Primatologia. Porto Alegre, RS, Brazil. [ Links ]
Calegaro-Marques, C., Bicca-Marques, J. C. 1996. Emigration in a black howling monkey group. International Journal Primatology. 17(2): 229–237. [ Links ]
Chapman, C. A. 1989. Ecological constraints on group size in three species of neotropical primates. Folia Primatologica. 73: 1–9. [ Links ]
Chapman, C.A., Balcomb, S. R. 1998. Population characteristics of howlers: ecological conditions or group history. International Journal Primatology. 19: 385–403. [ Links ]
Chapman, C.A., Chapman, L. J. 2000. Determinants of group size in social primates: The importance of travel costs. In: Boinski S, Garber PA, editors. On the move: how and why animals travel in groups. Chicago: University of Chicago Press. p 24–42. [ Links ]
Crockett, C. M. 1998. Conservation biology of the genus Alouatta. International Journal Primatology. 19: 549–578. [ Links ]
Digby, L. J. 1999. Sexual behavior and extragroup copulations in a wild population of common marmosets (Callithrix jacchus). Folia Primatologica. 70: 136–145. [ Links ]
Fialho, M. S., Setz, E. Z. F. 2007. Extragroup copulations among brown howler monkeys in southern Brazil. Neotropical Primates. 14 (1): 28–29. [ Links ]
Horwich, R. H. 1983. Breeding behaviors in the black howler monkey (Alouatta pigra) of Belize. Primates. 24: 222–230. [ Links ]
Jones, C. B. 1983. Social organization of captive black howler monkeys (Alouatta caraya): "Social competition" and the use of non-damaging behavior. Primates. 24: 25 – 39. [ Links ]
Kowalewski, M., Peker, S.M., Zunino, G. E. 2005. Promiscuous howlers: Extra-group copulation in female Alouatta caraya in northern Argentina. In: Programa e livro de resumos XI Congresso Brasileiro de Primatologia. Bicca-Marques, J. C. Editors. Porto Alegre/RS. Sociedade Brasileira de Primatologia. p: 116. [ Links ]
Kowalewski, M. M. and Garber, P. A. (in press). Mating promiscuity, energetics, and reproductive tactics in black and gold howler monkeys (Alouatta caraya). American Journal of Primatology. https://netfiles.uiuc.edu/mkowalew/www/Publications.html. [ Links ]
Mendes, S. L. 1989. Estudo ecológico de Alouatta fusca (Primates: Cebidae) na Estação Biológica de Caratinga, MG. Revista Nordestina de Biologia. 6(2): 71–104. [ Links ]
Neville, M. K., Glander, K.E., Braza, F., Rylands, A. B. 1988. The howler monkeys, genus Alouatta. In: Ecology and behavior of neotropical primates. Mittermeier, A. B., Coimbra – Filho, G. A. B., editors. World Wild Life Foundation. Washington DC: 349–453. [ Links ]
Pope, T. R. 1990. The reproductive consequences of male cooperation in the red howler monkey: Paternity exclusion in multi-male and single-male troops using genetic markers. Behavioral Ecology and Sociobiology. 27: 439–446. [ Links ]
Smuts, B. B. 1987. Sexual competition and mate choice. In: Primate Societies, BB. Smuts, D.L., Cheney, R. M., Seyfarth, R. W., Wrangham, T. T., Struhsaker (eds.), pp. 385–399. The University of Chicago Press, Chicago. [ Links ]
Zunino GE, Kowalewski MM, Oklander LI, González V. Habitat fragmentation and population size of the black and gold howler monkey (Alouatta caraya) in a semideciduous forest in Northern Argentina. Am J Primatol. 2007 Sep; 69(9): 966-75. [ Links ]
* Corresponding Address: Av. Bandeirantes, 3900, Monte Alegre, Campus USP, Ribeirão Preto, São Paulo, Brazil. FFCLRP-USP; departamento de Biologia. Zip code: 14040-901. Phone: 55 16 36023657; FAX: 55 16 36024886. e-mail: wagnerf@usp.br


{kind=link}
{kind=link}