










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkRevista Brasileira de Terapia Comportamental e Cognitiva
versão impressa ISSN 1517-5545
Rev. bras. ter. comport. cogn. vol.5 no.1 São Paulo jun. 2003
ARTIGOS
Ratos wistar sob regimes rotineiros de restrição hídrica e alimentar1
Wistar rats under water and food restriction
Gerson Yukio Tomanari, Alexandre de Souza Pine ; Maria Teresa Araújo Silva
Universidade de São Paulo
RESUMO
Restrição hídrica ou alimentar é uma das operações motivacionais mais utilizadas em estudos que envolvem processos básicos de aprendizagem operante com animais de laboratório. O objetivo do presente estudo foi descrever, de forma sistemática, a evolução dos pesos de ratos expostos ou não a uma rotina de restrição controlada de acesso à água ou ao alimento por cerca de 7 meses, incluindo breves interrupções ocasionais. Para tanto, foram tomados diariamente os pesos de 29 ratos (Rattus norvegicus), linhagem Wistar, criados e alojados nas condições específicas do Biotério de roedores do Departamento de Psicologia Experimental do IPUSP durante o período que compreendeu o 5º ao 217º dia de vida dos animais. Duas fases foram planejadas. A primeira referiu-se aos primeiros 106 dias de pesagem; nesta, todos os animais dispunham de água e alimento à vontade. A segunda fase referiu-se ao período do 107º ao 217º dia; nesta, foram formados três grupos de sujeitos: Grupo Ad Lib (n = 10), para o qual água e comida estiveram constantemente disponíveis, e Grupos Restrição Hídrica (n = 10) e Restrição Alimentar (n = 9), para os quais, a partir do 107o dia de vida, a quantidade diária de água e alimento foi fixada em, respectivamente, 23 ml e 17 g. Para os sujeitos do Grupo Restrição Hídrica, o acesso ao alimento era irrestrito; para os sujeitos do Grupo Restrição Alimentar, o acesso à água era irrestrito. Nesta fase, foram efetuadas duas interrupções sucessivas na privação por períodos de 15 e 5 dias. Os resultados mostraram que a curva de aquisição de peso de ratos com acesso livre à água e ao alimento revelou ganho de massa positivamente acelerado durante o primeiro mês de vida, ainda que relativamente lento, seguido imediatamente por um aumento relativamente mais rápido, porém negativamente acelerado, tendendo a valores assintóticos. Restrito o acesso à água ou ao alimento no 107º dia de vida, o peso dos animais diminuiu gradual e progressivamente. Quanto às interrupções no procedimento de privação, estas resultaram em ganhos de peso, os quais se mostraram, interessantemente, cada vez mais elevados a cada interrupção. Além disso, a cada nova retomada no procedimento de privação, verificou-se uma maior resistência à diminuição de peso a níveis anteriormente registrados. Estes dados permitem compreender padrões gerais de aquisição de massa corpórea de ratos, na presença e ausência de privação, ao longo de um período que compreende tipicamente a duração de um experimento comportamental. Nesse sentido, constituem parâmetros experimentais em estudos futuros.
Palavras-chave: Ratos wistar, Restrição hídrica, Restrição alimentar, Peso corporal.
ABSTRACT
Water or food restriction is among the most frequently used motivational operations in studies of basic processes of operant learning in laboratory animals. The aim of the present study was to investigate systematically the body weight development of rats submitted to controlled water or food restriction for about 7 months, as compared to animals under an ad Lib regimen. Brief occasional interruptions of the restriction schedule were carried out. Subjects were 29 Wistar rats bred at the Experimental Psychology Department Animal House. They were weighed daily from Day 5 to Day 217 after birth. In the first experimental phase all animals had ad Lib access to water and food for 106 days. In the second phase, from Day 107 to Day 217, subjects were divided in three groups. Animals in the Ad Lib group (n = 10) had permanent access to both water and food; animals in the Water Restriction group (n = 10) had daily access to 23 ml water and free access to food, whereas subjects in the Food Restriction group (n=9) were allowed daily access to 17 g of food and free access to water. During this phase there were two interruptions in the restriction regimen, lasting 15 and 5 days. Results showed a positive though slow acceleration of body mass during the first month after birth, followed by a relatively more rapid increase tending to asymptotic values. After the restriction schedules started on Day 107 body weight declined gradually. Interruptions in the restriction procedure resulted in increased body weight, which was more pronounced after the second interruption. In addition, after each interruption a greater resistance to body weight loss was observed. The data illustrate the general pattern of body mass acquisition in rats, in the presence and absence of water or food restriction, along a period corresponding to a typical behavioral experiment. They can thus be useful as experimental parameters in future experiments.
Keywords: Wistar rats, Water restriction, Food restriction, Body weight.
Uma das operações motivacionais mais utilizadas em estudos que envolvem processos básicos de aprendizagem (o comportamento operante, por exemplo) com animais de laboratório é a restrição hídrica ou alimentar. Estabelecida a motivação, a apresentação de água ou alimento é então utilizada como conseqüência de comportamentos cuja probabilidade de ocorrência futura, aumentada, permite caracterizar a água ou o alimento como estímulos reforçadores do comportamento que se investiga. Ambas as restrições de água e alimento são bastante utilizadas, e podem produzir efeitos muitas vezes distintos sobre o comportamento (Davey & Cleland, 1982). No entanto, a decisão por uma ou outra em geral se fundamenta em características do fenômeno investigado, do delineamento experimental empregado e das condições de infra-estrutura disponível.
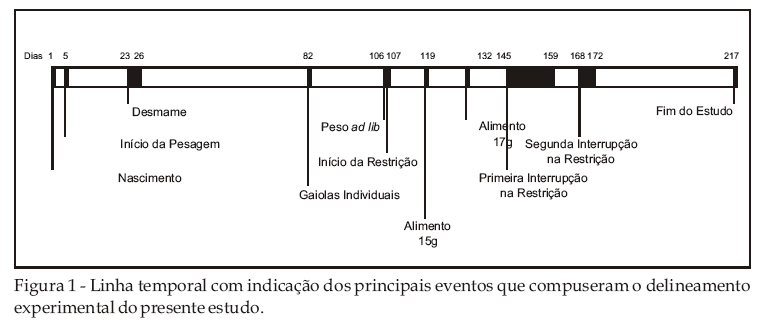
Os laboratórios de análise comportamental do Departamento de Psicologia Experimental do IPUSP (em particular os Laboratórios de Análise Experimental do Comportamento, Psicofarmacologia e de Análise Biocomportamental) utilizam ratos machos, da cepa Wistar, que, via de regra, são criados no próprio biotério do Departamento. Após o desmame, até seis animais compartilham gaiolas coletivas por cerca de 60 dias, quando então são transferidos para gaiolas individuais para que se inicie a execução do planejamento experimental. Até esse momento, o acesso dos animais à água e ao alimento é livre e irrestrito (ad Libitum).
Imediatamente antes do início de qualquer planejamento experimental que requeira um regime de restrição de água ou alimento, os ratos são pesados de modo a se obter o chamado peso ad lib, o qual é utilizado como parâmetro a partir do qual pesos por volta de 85% constituem o denominado peso de privação. Consagrado na literatura e na prática de pesquisadores na área, o peso de privação é aquele que, oriundo da operação de privação, estabelece o valor reforçador da água ou do alimento durante as sessões experimentais, alterando, assim, a probabilidade de emissão de comportamentos que os produzem.
Basicamente, a operação utilizada em nossos laboratórios para manter os animais no peso de privação ao longo do período de experimentação consiste em restringir a quantidade diária de água ou alimento fornecida aos animais. Controla-se diretamente essa quantidade (n ml de água ou y g de alimento) ou, por via indireta, controla-se o tempo de acesso a eles (x minutos de acesso à água ou ao alimento). Animais sob privação de comida recebem uma quantidade restrita de ração padrão por dia, mantendo-se o acesso à água livre; animais sob restrição hídrica recebem uma quantidade restrita de água, mantendo-se o acesso livre ao alimento. Seja tempo ou quantidade o parâmetro de controle para a privação por água ou alimento, estes são apresentados aos animais sempre cerca de 15 minutos após o término do experimento.
Muito freqüentemente, os estudos comportamentais empregam ratos que iniciam o procedimento de privação entre os 90 e 120 dias de vida. A essa idade, esses animais são considerados adultos jovens, ainda sob crescimento corporal, apesar das taxas menos acentuadas relativamente à fase precedente de maturação (Mitiko, 1993). Esses estudos normalmente submetem os animais a regime de privação hídrica ou alimentar por 3, 4, 6 meses ou mais. Portanto, o crescimento nesse período é um fator que muito provavelmente interage com a privação.
Durante o período que compreende um experimento, é prática manter o peso dos animais em torno de 85% do peso ad lib originalmente medido. Muitas vezes, como forma de contemplar o crescimento do animal durante o período que acompanha a privação, estabelece-se uma faixa de valores de peso pré-estabelecidos (por exemplo, de 80% a 85% do peso ad lib), permitindo pequenos aumentos graduais no peso ao longo do tempo. Entretanto, tais aumentos são conduzidos de forma não sistemática, na ausência de parâmetros consolidados para isso.
Com base nessas considerações, o objetivo deste trabalho foi investigar a evolução dos pesos de ratos em função de sua idade, isto é, desde o nascimento até o 217º dia de vida, sob condições de acesso irrestrito à água e ao alimento, bem como sob regime de restrição hídrica ou alimentar a partir do 107º dia de idade. Trata-se, portanto, de considerar a privação como uma variável motivacional que tem no peso corpóreo do animal um correlato direto. Ao fornecer parâmetros relativos ao procedimento de privação, o presente trabalho procura contribuir aos estudos em que os seus efeitos sobre o comportamento são diretamente investigados (por exemplo, Carlton, 1961; Conrad, Sidman, & Herrnstein, 1958; Fischer & Fantino, 1968; Leander, 1973; López-Pierce & Martínez, 2001; Epling & Boer, 1986).
Em estudos experimentais do comportamento, os períodos de privação, até mesmo por sua longevidade, são muitas vezes sujeitos a inevitáveis interrupções decorrentes de feriados prolongados ou interferências por manutenção do biotério. Na tentativa de reproduzir algumas das condições tipicamente encontradas em contextos experimentais, o presente estudo procurou ainda contemplar interrupções na privação com o objetivo de descrever os seus efeitos sobre o peso dos animais.
Método
Sujeitos
Foram utilizados 29 ratos albinos Wistar, machos, nascidos e mantidos no Biotério do Departamento de Psicologia Experimental do Instituto de Psicologia da USP. Os animais eram provenientes de linhagem do Biotério da Fundação Parque Zoológico do Estado de São Paulo. Ao longo do estudo, vigorou um ciclo claro-escuro de 12 h (luz acesa das 7:00 às 19h). Do nascimento até o 82º dia de vida, os animais ficaram alojados em gaiolas coletivas de plástico translúcido medindo 40 x 33 x 17 cm, com tampa de grade metálica onde água e ração balanceada Purina Labina2 encontravam-se permanentemente disponíveis. As gaiolas eram forradas com maravalha autoclavada. A partir do 83º dia, os animais foram transferidos para gaiolas individuais de aço inox, com dimensões 24 x 17 x 19 cm, cujas partes frontal e inferior eram constituídas de grade quadriculada.
Equipamento
Foi utilizada uma balança eletrônica Metler, modelo AS-F1, com precisão de 0,01g.
Procedimento
O procedimento pode ser dividido em duas fases. Na primeira, a todos os animais, água e alimento encontravam-se constantemente disponíveis, isto é, à vontade (ad libitum ou, simplesmente, ad lib). Na fase seguinte, tiveram início os procedimentos de restrição hídrica e alimentar aplicados a diferentes grupos de sujeitos.
Fase ad lib: Por meio de pesagem diária, acompanhou-se nessa fase o desenvolvimento da massa corpórea dos 29 ratos machos sob regime de amamentação e, posteriormente, sob regime de água e alimento ad lib, durante os 106 dias que antecederam a fase de restrição. Os sujeitos foram primeiramente instalados nas gaiolas coletivas usadas rotineiramente no Biotério (ver descrição detalhada acima). Cada gaiola continha uma fêmea que amamentava de 5 a 6 filhotes. O desmame, com a retirada da fêmea, deu-se entre os dias 25 e 28 de vida. Os animais permaneceram nessas gaiolas coletivas até o 82º dia de idade, quando então foram transferidos para as gaiolas individuais. Enquanto permaneceu nas gaiolas coletivas, cada sujeito foi identificado por marcas gráficas feitas nas regiões da cauda e da orelha com caneta hidrocolor de tinta permanente.
A pesagem dos animais iniciou-se no dia 5 do estudo, quando sua idade variava de 3 a 6 dias -- para efeito de uniformização, consideraremos que todos os animais foram pesados a partir do 5º dia de vida. Os animais eram transportados em suas próprias gaiolas até a balança, distante cerca de 7 m da sala onde estavam alojados. Um a um, eram então manuseados e colocados em um pequeno recipiente sobre o prato da balança. A pesagem tinha início entre 18:00 e 19:00 h, durava cerca de 1h 30m e foi efetuada diariamente, inclusive em fins de semana e feriados, à exceção dos dois períodos de interrupção descritos oportunamente.
Fase Restrição: Essa fase teve por objetivo acompanhar o desenvolvimento das massas corpóreas dos ratos quando submetidos à restrição hídrica ou alimentar. Três grupos de sujeitos foram formados na ocasião do alojamento dos animais nas gaiolas individuais. Um primeiro grupo foi destinado à privação de alimento (Grupo Restrição Alimentar), um segundo grupo à privação de água (Grupo Restrição Hídrica), e um terceiro grupo foi mantido nas condições ad lib, servindo como grupo controle.
A alocação de sujeitos aos grupos contou previamente com a elaboração de uma lista em que os seus pesos foram colocados em ordem crescente independentemente da gaiola coletiva em que esses se encontravam até aquele momento. A partir desta lista, os sujeitos foram alocados seqüencialmente a cada um dos três grupos, procurando-se garantir, assim, uma distribuição relativamente eqüitativa entre os grupos com relação aos pesos dos animais. O Grupo Restrição Hídrica foi constituído por 10 sujeitos. A partir do dia 107º dia, exceto durante as interrupções notadas abaixo, cada animal desse grupo recebeu a quantidade diária de 23 ml de água, a qual, na rotina de nossos laboratórios, mantém o peso dos animais em cerca de 85% do peso ad lib. O Grupo Restrição Alimentar foi constituído por 9 sujeitos. Entre o 107º e 118º dia, cada um dos sujeitos desse grupo recebeu 13 g de ração. A partir do 119º dia, a quantidade diária de alimento foi então aumentada para 15 g, permanecendo assim até o 131º dia, quando então passou a 17 g até o final do estudo, no 217º dia. A quantidade de alimento fornecida diariamente teve como parâmetro a manutenção do peso dos animais em cerca de 85% do peso ad lib. Tal como no Grupo Restrição Hídrica, o procedimento de privação foi suspenso nas duas oportunidades descritas adiante. O Grupo ad lib foi constituído por 10 sujeitos, e continuou com alimento e água à vontade do 107º ao 217º dia.
Duas interrupções no procedimento de privação foram efetuadas. A primeira teve a duração de 15 dias (do 145º e 159º dia) e a segunda de 5 dias (do 168º ao 172º dia). Durante esses dois intervalos, foram suspensos os regimes de restrição e a pesagem diária, permanecendo os sujeitos dos três grupos com alimento e água constantemente disponíveis.
Os eventos que compuseram o procedimento do estudo encontram-se esquematizados graficamente na Figura 1.
Resultados e Discussão
A Figura 2 mostra os pesos médios de Rattus norvegicus, linhagem Wistar, criados e alojados nas condições específicas do Biotério de roedores do Departamento de Psicologia Experimental do IPUSP durante o período que compreende o 5º até o 217º dia de vida. Nesta figura, encontram-se representados os dados dos três grupos de sujeitos: Grupo ad lib (n = 10), para o qual água e comida estiveram constantemente disponíveis ao longo de todo o estudo, e Grupos Restrição Hídrica (n = 10) e Restrição Alimentar (n = 9), para os quais, a partir do 107º dia de vida, a quantidade diária de água e alimento foi fixada em, respectivamente, 23 ml e 17 g. Para os sujeitos do Grupo Restrição Hídrica, o acesso ao alimento era irrestrito; para os sujeitos do Grupo Restrição Alimentar, o acesso à água era irrestrito. No eixo vertical, à direita no gráfico, constam valores de porcentagem que correspondem aos pesos dos animais tendo-se como referência (i.e., 100%) o peso médio de todos os 29 sujeitos na data imediatamente anterior ao início dos procedimentos de privação de água e de alimento (isto é, o 106º dia), o qual foi estabelecido como sendo o peso ad lib médio. As interrupções nas curvas (do 145º ao 159º dia e do 168º ao 172º dia) referem-se aos períodos em que os procedimentos de privação e pesagem estiveram suspensos.
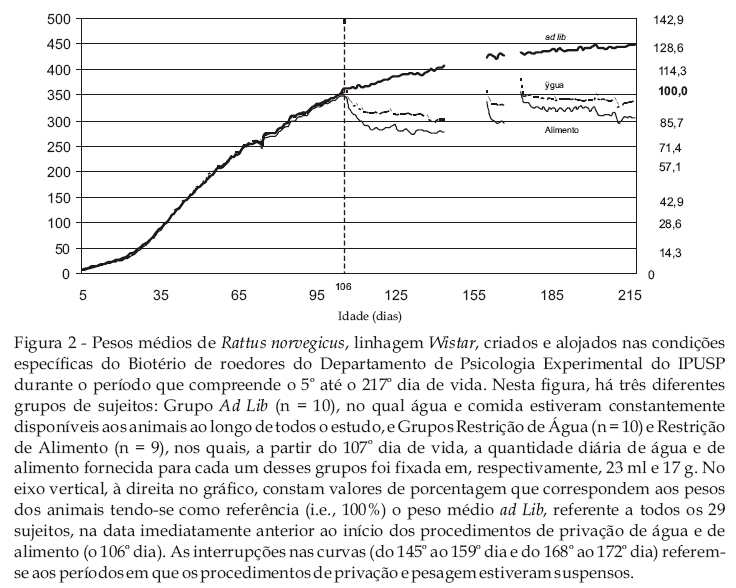
Esta figura permite observar, por meio dos dados do Grupo ad lib, a curva de crescimento -- tal como expressa por massa corpórea -- na condição em que água e comida estavam constantemente disponíveis. Trata-se de uma curva que mostra os primeiros 7 meses de vida dos animais e permite verificar, de modo geral, um ganho constante de peso que, inicialmente, mostra-se positivamente acelerado mas que, no decorrer dos dias, desacelera-se gradativamente tendendo a uma relativa estabilidade assintótica. Essa é a tendência conhecida na evolução ponderal de ratos Wistar (Curi, Hell, Bazotte, & Timo-Iaria, 1984; Machado, 1989). Nos dias iniciais de vida, antes de completar o primeiro mês desde o nascimento, apesar da aceleração positiva, o ganho de peso é menor do que se observa no período imediatamente a seguir (do segundo ao quarto mês de vida), período em que se destaca uma maior aceleração de crescimento, ainda que negativa. Uma possibilidade de se compreender esta menor aceleração no aumento do peso no primeiro mês de vida deve considerar o fato de que, nesse período, os animais eram mantidos em caixas coletivas que incluía a progenitora e outros animais. Havia, portanto, restrições impostas, por exemplo, pela disponibilidade de alimento e pela competição na amamentação que, em gaiolas individuais, com água e alimento à vontade, deixaram de estar presentes.
Observando-se os pesos médios dos animais do 5º ao 106º dia do estudo, verifica-se que os dados dos três grupos experimentais são praticamente indistintos. As três curvas que representam cada um dos grupos, nesse período, sobrepõem-se umas às outras. Até mesmo variações pontuais, como a queda de peso ocorrida no 75º dia, por exemplo, encontram-se representadas nos dados dos três grupos simultaneamente. Este fato (1) revela que o fator responsável por essa queda possivelmente decorre de uma variável intercorrente que afetou os pesos de todos os sujeitos, independentemente do grupo a que pertenciam e (2) demonstra o rigoroso controle experimental em vigor, seja em termos da precisão, sejam em termos da sensibilidade das medições efetuadas.
A partir do 107º de vida, entretanto, as curvas isolam-se umas das outras. Os pesos dos animais do Grupo ad lib aumentaram gradualmente, ainda que, nesse momento, sob aceleração negativa. Já os pesos dos animais pertencentes aos Grupos Restrição Hídrica e Restrição Alimentar refletiram, imediatamente, o início do procedimento de privação. Para estes dois grupos, porém mais acentuadamente para o segundo, houve uma diminuição gradual e constante dos pesos ao longo dos dias que se sucederam ao início da privação. No decorrer de todo o procedimento de privação, sem exceção, os pesos médios dos animais do Grupo Restrição Hídrica foram sempre superiores àqueles do Grupo Restrição Alimentar. Essa diferença sistemática entre os grupos deve estar estritamente ligada à quantidade fixa de água e alimento fornecida para cada um dos grupos, isto é, 23 ml de água e 17 g de comida. Teoricamente, poder-se-ia aproximar os pesos dos animais sob privação diminuindo-se a quantidade diária de água fornecida para os sujeitos do Grupo Restrição Hídrica e/ou aumentando-se a quantidade diária de alimento fornecida para os sujeitos do Grupo Restrição Alimentar. Entretanto, outros fatores devem ser levados em conta quando se tenta prever os efeitos de aumentos ou diminuições de água ou alimento sobre os pesos dos animais. Entre eles, a estabilidade das condições ambientais do Biotério, o esquema de fornecimento diário de água ou alimento, as variações incontroláveis das condições climáticas externas etc. A despeito de tais fatores, contudo, os efeitos de um ou outro nível de privação devem ser avaliados fundamentalmente com relação ao desempenho dos animais sob as contingências experimentais. Além disso, deve-se ter claro que os dados obtidos no presente estudo, até que se proceda com replicações que permitam a sua generalização, mantêm referência estreita com as especificidades dos sujeitos experimentais utilizados, as condições particulares de manejo a que foram submetidos, e as condições particulares do Biotério em que foram criados e mantidos. Sem se considerar tais especificidades, a relação entre a quantidade de água ou alimento fornecida e o peso resultante deve ser estendida a novos contextos com muita cautela.
Dados revelados na Figura 2, adicionalmente aos parâmetros fornecidos pela curva de crescimento ad lib, mostram os efeitos da privação quando acompanhada de seguidas interrupções e retomadas. Na curva do Grupo ad lib, pode-se verificar que os períodos que correspondem a essas interrupções simplesmente não afetam o curso contínuo de ganho de peso dos animais. No entanto, nas curvas dos grupos sob restrição de água e alimento, pode-se observar que aos períodos de ausência de privação seguem-se aumentos de peso que, a despeito de se manterem fixas as quantidades de água e de alimento fornecidas diariamente aos animais, são mais elevados, a cada nova interrupção, e parecem mais resistentes à diminuição, a cada retomada da privação.
Em termos específicos, pode-se verificar que, nos dias que imediatamente antecederam o início do primeiro período de interrupção do procedimento de privação, o peso médio dos ratos do Grupo Restrição Hídrica foi cerca de 85% do peso médio registrado no dia imediatamente anterior ao início da privação (peso ad lib, isto é, aquele obtido no 106º dia). O valor percentual do peso médio dos ratos do Grupo Restrição Alimentar, comparativamente, foi um pouco menor, isto é, cerca de 80%. Logo após o encerramento do primeiro período de interrupção, os pesos médios de ambos os grupos aumentaram para níveis superiores ao peso ad lib. Nos dias subseqüentes à retomada do procedimento de privação (do 160º ao 167º dia), observa-se uma queda gradual e acentuada nos pesos médios dos animais. Até o 167º dia, no entanto, os pesos não regrediram aos níveis registrados nos dias que antecederam a interrupção. Neste dia, o peso médio do Grupo Restrição Hídrica representou cerca de 93% do peso ad lib, contra 85% registrado no dia que antecedeu a interrupção. Para o Grupo Restrição Alimentar, os valores correspondentes foram 85% e 80%.
Na seqüência, vigorou, entre o 168º e 172º dias, o segundo período de interrupção no procedimento de privação. Nessa ocasião, verifica-se que as curvas reproduzem um padrão semelhante àquele da primeira interrupção. Ou seja, os pesos médios registrados imediatamente após a retomada da privação (173º dia) atingem valores mais elevados do que os registrados no início (dia 107º) e na retomada (dia 160º) dos procedimentos de privação.
No prosseguimento da privação que acompanha o período final deste estudo após as duas interrupções, verifica-se uma diminuição inicialmente abrupta dos pesos dos animais, seguida por um período em que é lenta e gradual. Transcorridos os 45 dias de privação que se seguiram ao segundo período de interrupção (do 173º a 217º dia), os pesos atingiram, no último dia de coleta de dados, 95% e 88% do peso ad lib (Grupos Restrição Hídrica e Restrição Alimentar, respectivamente). Esses valores, ainda que em queda, são superiores àqueles registrados ao final do período que antecedeu a primeira interrupção (do 107º ao 144º dia), o qual era comparativamente menor -- 38 versus 45 dias. Trata-se, aparentemente, de uma maior resistência à recuperação do peso sob privação que parece se manifestar a cada retomada do procedimento de restrição hídrica ou alimentar. Ainda que, verdadeiramente, não se possa afirmar a que pesos chegariam os animais caso o período de privação fosse prolongado ainda mais.
Em suma, os dados mostram que interrupções do procedimento de privação são seguidas imediatamente pela recuperação do peso dos animais, o qual, a cada nova interrupção, tende a valores progressivamente mais elevados. A introdução do regime restritivo é acompanhada por perdas graduais e constantes de pesos. Entretanto, a cada retomada da privação, nota-se que, mesmo mantendo-se fixa a quantidade diária de água ou alimento, os pesos resistem progressivamente à diminuição, não chegando a atingir os valores anteriormente registrados, ao menos para os períodos de privação efetuados, os quais tiveram duração de 15 e 5 dias. Interessantemente, tal resistência à diminuição no peso foi verificada mesmo no período que se sucedeu à segunda interrupção da privação, o qual foi mais longo do que o que antecedeu a primeira, o que, supostamente, teria propiciado um tempo maior para a redução progressiva do peso. Fenômeno semelhante foi observado em estudo anterior, no qual se demonstrou que ciclos repetidos de restrição calórica acarretam, em ratos, uma taxa menor de perda de peso, bem como uma aceleração posterior de ganho (Brownell, Greenwood, Stellar, & Shrager, 1986). Algumas hipóteses podem ser levantadas para explicar esse efeito:
1) durante os períodos de suspensão da privação, podem ter-se desenvolvido tecidos que posteriormente não foram susceptíveis a diminuição a despeito da restrição hídrica ou alimentar (tecido ósseo, por exemplo);
2) o perfil de peso corporal é função da idade em que determinado regime alimentar é adotado (Curi, Bazotte, Hell, & Timo-Iaria, 1989). Ora, animais alimentados à vontade chegam a um peso estável a partir dos 6 meses, aproximadamente. Até essa altura, vão reduzindo a ingestão de alimento, pois a estabilidade de seu peso tende a ser mantida por uma quantidade menor do que a ingerida durante a fase de aceleração de peso (Machado, 1989). No presente estudo, os animais do Grupo Restrição Alimentar continuaram a receber a mesma quantidade de comida após as interrupções de restrição. É possível que talvez estivessem então submetidos a um grau menor de privação, o que poderia explicar porque os pesos dos animais dos grupos Restrição Hídrica e Alimentar não retornaram ao nível pré-interrupção.
3) entretanto, a diferença entre o peso nos períodos pré-interrupção e pós-interrupção é muito grande e abrupta para ser entendida apenas como resultado de um possível aumento proporcional de quantidade de alimento ingerida. Uma outra possibilidade é a de que o organismo aprendesse a conservar energia/gordura quando de novo ameaçado de privação. O mecanismo dessa aprendizagem poderia ser um ajuste hormonal condicionado quando da privação anterior, provavelmente em resposta a estímulos condicionados internos. De fato, animais em restrição alimentar utilizam o alimento mais eficientemente (Curi et al., 1984), mostrando maior capacidade de converter carboidratos em material energético armazenado como lípides e glicogênio (Leveille, 1970). Ou seja, aprendem a tirar proveito máximo do pouco que podem comer, desenvolvendo mecanismos de alta eficiência metabólica, como maior absorção intestinal de nutrientes (Leveille & Chakrabarty, 1968), aumento de síntese e armazenamento de lípides no tecido adiposo (Leveille, 1970), aumento do glicogênio hepático (Lima, Hell, Timo-Iaria, Scrivoletto, Dolnikoff, & Pupo, 1981), manutenção da glicemia e menor mobilização de ácidos graxos livres (Curi et al., 1989). Possíveis mudanças adaptativas contribuindo para melhor utilização de nutrientes poderiam ser a redução observada na taxa de esvaziamento gástrico (Lima et al., 1981) e o aumento da área de absorção intestinal (Leveille & Chakrabarty, 1968) sob regime de restrição alimentar. Embora as mudanças metabólicas sejam revertidas após 5 semanas de livre acesso a comida (Curi et al., 1989), é plausível supor que elas ainda persistam por algum tempo após a suspensão da privação. Se esse tempo for superior a 15 dias, o aumento relativo de peso dos ratos do Grupo Restrição Alimentar poderia ser, ao menos em parte, devido à persistência dessas adaptações.
A Figura 3 mostra os pesos individuais de cada rato, do 5º ao 217º dia de vida, período que compreendeu a coleta de dados do presente estudo. O gráfico superior agrupa as curvas dos sujeitos do Grupo ad lib, os quais tiveram água e alimento constantemente disponíveis. Os gráficos central e inferior referem-se aos dados dos sujeitos pertencentes aos Grupos Restrição Hídrica e Alimentar, respectivamente. A reta vertical tracejada indica o início da privação para os sujeitos dos Grupos Restrição Hídrica e Alimentar. Encontram-se indicados nos gráficos os dois períodos em que houve interrupção nas rotinas de privação e pesagem dos animais.
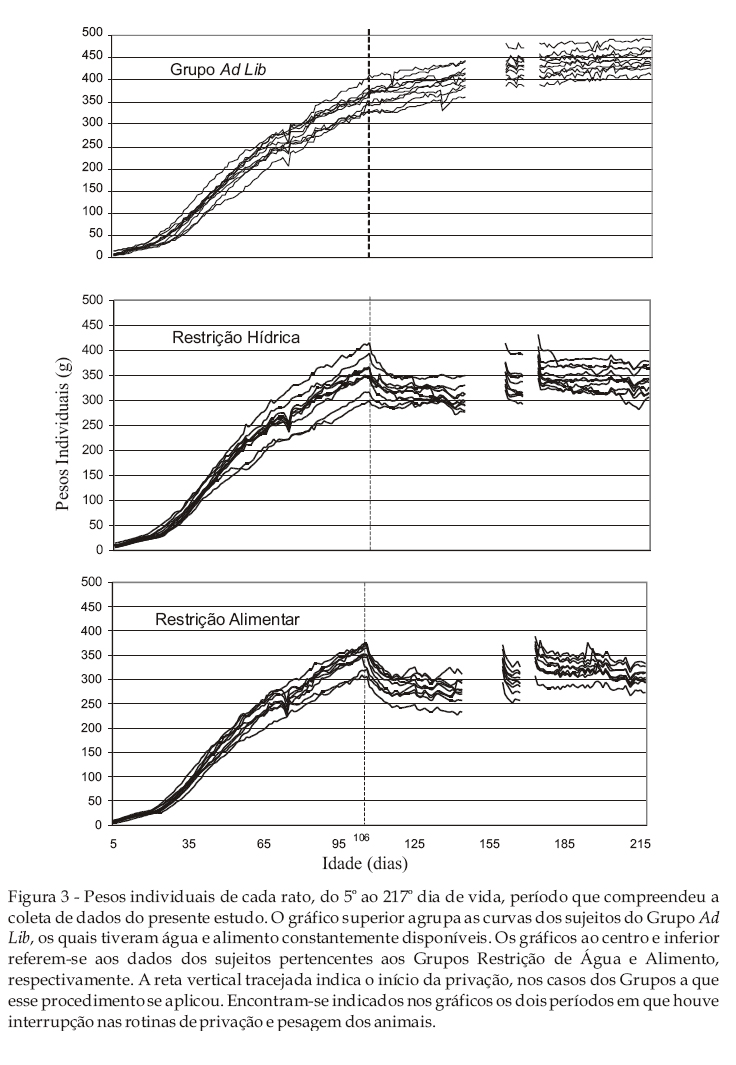
De modo geral, os dados apresentados nos três gráficos da Figura 3 mostram uma clara correspondência entre os pesos individuais dos sujeitos e a média que representa o conjunto deles (Figura 2). Exceto por diferenças de amplitude, as curvas individuais, de qualquer que seja o grupo de sujeitos, reproduzem muito fielmente as características básicas da curva de crescimento de sujeitos que dispõem de água e alimento ad lib, tanto quanto as curvas resultantes dos procedimentos de privação. Nas curvas dos sujeitos sob privação, em particular, os efeitos das duas suspensões e retomadas da privação podem ser observados com relativa precisão nos dados individuais de cada sujeito. Em um certo sentido, portanto, a sistematicidade dos dados entre os sujeitos demonstra um adequado controle experimental e propicia um alto grau de generalização interna dos mesmos. Essa sistematicidade é especialmente observada na regularidade nas curvas individuais no que diz respeito às distâncias que estas mantêm entre si ao longo de todo o estudo. Esta regularidade torna-se mais evidente à medida que os animais aumentam progressivamente de peso. Nesse caso, pode-se observar, por exemplo, que o rato que nos meses iniciais de vida era o mais pesado entre os membros de seu grupo, assim permanece praticamente até o final da coleta de dados. O mesmo pode ser verificado com relação aos demais sujeitos e a sua condição hierárquica correspondente. Ademais, é interessante notar que a manutenção da posição relativa estabelecida pelos pesos dos animais ocorre mesmo na vigência do procedimento de privação. Ao que parece, portanto, esse dado sugere que fatores constitucionais de cada indivíduo mantêm-se expressos a despeito dos efeitos de um fator circunstancial preponderante como é a privação de água ou alimento.
Considerações Finais
Em pesquisas que envolvem comportamentos operantes, ratos albinos são normalmente submetidos a um procedimento de privação hídrica ou alimentar que visa a tornar água ou alimento eventos reforçadores, possibilitando, assim, a manipulação experimental de variáveis comportamentais (Michael, 1982). Porque tais pesquisas fazem uso de delineamentos experimentais em que o comportamento de cada sujeito é analisado tendo-o como seu próprio controle (Sidman, 1960), o tempo de coleta de dados e a vigência do procedimento de privação podem se mantidos por até vários meses. Descrever, de forma sistemática, a evolução dos pesos de ratos expostos e não expostos a uma rotina de restrição controlada de acesso à água ou ao alimento por um período prolongado que se inicia desde os primeiros dias de vida foi, nesse contexto, o objetivo central do presente estudo.
Para tanto, procurou-se reproduzir as condições vigentes nos Laboratórios do Departamento de Psicologia Experimental da USP. Sendo assim, foram empregados ratos Wistar (Rattus norvegicus), machos, criados e mantidos no Biotério de Roedores do próprio Departamento, submetidos ao fornecimento diário restrito de água (23 ml) ou de alimento (17 g). Os pesos dos animais foram registrados diariamente por 213 dias (do 5º ao 217º dia de vida) e comparados aos de um terceiro grupo de sujeitos, mantidos sob as mesmas condições, exceto quanto ao acesso livre à água e ao alimento. Dados individuais de cada sujeito e médias dos grupos foram analisados e revelaram padrões claros de aquisição e perda de peso como resultado da instalação, suspensões e retomadas sucessivas das condições de privação. Esses padrões demonstraram uma forte consistência entre os dados individuais e a média grupal que os representam.
Resumidamente, a curva de aquisição de peso de ratos com acesso livre à água e ao alimento mostra ganho de massa corpórea positivamente acelerado durante o primeiro mês de vida, ainda que relativamente lento, seguido por um aumento mais rápido, porém negativamente acelerado, tendendo a valores assintóticos. Restrito o acesso à água ou ao alimento a partir do 107º dia de vida, o peso desses animais tendeu a diminuir gradual e progressivamente. O estudo contou com duas interrupções sucessivas na privação por períodos de 15 e 5 dias, nessa ordem. A cada interrupção no procedimento de privação, observaram-se ganhos progressivos de peso. A cada retomada desse procedimento, verificou-se uma maior resistência ao retorno dos pesos aos níveis anteriores. Aparentemente, os dados sugerem a possibilidade de os pesos dos animais privados tenderem a acompanhar os pesos que teriam em condições de acesso livre à água ou ao alimento.
Por um lado, o presente trabalho, especialmente em seus aspectos metodológicos, aproxima-se de uma série de estudos disponíveis na literatura que procuram descrever padrões sistemáticos de aquisição, perda e manutenção de pesos em diferentes espécies de animais (Denise & Brinks, 1985; Morrow, McLaren, & Butts, 1978; Nadarajah, Marlowe, & Notter, 1984). Por outro, entretanto, o fato de estar inserido no âmbito das investigações comportamentais atribui a este estudo características particulares. Nesse sentido, o peso dos animais deve ser considerado um correlato de diferentes graus de privação. Como medida correlata, estabelece parâmetros que permitem, de alguma forma, antecipar o valor reforçador de um estímulo -- água ou alimento -- no momento em que a apresentação do mesmo constituir parte de contingências experimentais. Em última instância, portanto, esse papel reforçador é avaliado quando estes estímulos passarem a fortalecer comportamentos que os produzirem.
Conforme apontado anteriormente, procedimentos de privação nem sempre consideram o crescimento dos animais como uma variável preponderante. Prática corrente em laboratórios de estudos de condicionamento operante (Iversen & Lattal, 1991; Matos & Tomanari, 2002; Reese, 1967), o início da privação dá-se anteriormente ao início claro da estabilidade assintótica da curva de crescimento dos animais. Uma conseqüência disso é que o peso de privação do animal, com o passar dos dias, representa uma porcentagem cada vez maior com relação ao peso que o animal teria caso estivesse com água ou alimento constantemente disponível. Por exemplo, um animal que, no 100º dia de vida pesasse 350 g, considerado 85% deste, teria 300 g como peso em privação. No 200º dia de vida, este animal teria, teoricamente, 400 g como peso ad lib. Com relação a esse peso potencial, o peso em privação (i.e., 300 g) representaria 75%. Ou seja, considerando-se a projeção de crescimento do animal, a porcentagem de privação diminui, o que poderia significar um aumento progressivo no grau de privação imposto, pois a quantidade diária de água ou alimento que mantinha o animal a 85% do peso ad lib aos 100 dias de idade resiste em mantê-lo com este mesmo peso aos 200 dias, como os dados do presente estudo revelam. Com base nessa observação, é prática comum entre pesquisadores aumentar, ao longo de um experimento, a quantidade diária de água ou alimento que se fornece ao animal, de modo a regular, em longo prazo, o nível de privação inicialmente estabelecido às condições vigentes.
Considerando a possível relevância em integrar o crescimento do animal ao procedimento de privação, a Figura 4 apresenta diferentes modelos de aquisição e manutenção de pesos. De acordo com o modelo típico de privação, toma-se o peso ad lib de um animal com cerca de 90 a 120 dias de vida. Tendo esse valor como referência, estabelece-se uma porcentagem deste (85% a 90%, por exemplo) como sendo o peso em privação. O peso em privação é em geral mantido, por meio da restrição de água ou alimento, ao longo do período de coleta de dados de um experimento (linha tracejada horizontal na Figura 4).
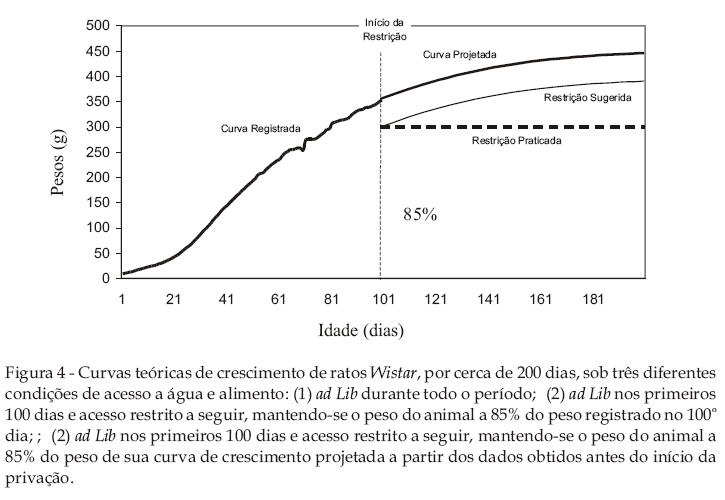
A partir dos dados descritos neste estudo, é possível estabelecer parâmetros que possam atender a um aumento experimentalmente controlado nas situações que envolvam longos períodos de privação e que utilizem animais em fase de crescimento progressivo. Assim, o estado de privação dos animais poderia atender continuada e uniformemente a sua função estabelecedora de um estado fisiológico e motivacional mesmo por longos períodos, dado que estaria em consonância com as curvas típicas de crescimento de animais que compartilham origens e condições de manejo semelhantes.
Teoricamente, animais que estivessem sob estabilidade de crescimento exigiriam pouco ou nenhum ajuste no fornecimento diário de água ou alimento. Os dados do presente estudo, preservadas as suas condições particulares, sugerem esse início a partir de 6 meses de idade. Obviamente, a implementação desta sugestão pode vir acompanhada de dificuldades decorrentes do tempo que se tem de esperar até que os animais atinjam essa idade. Por exemplo, os animais teriam de ser mantidos no Biotério em condições ad lib por mais tempo, assim como o início de um experimento talvez tivesse de ser postergado.
No entanto, à luz da prática corrente em experimentação animal, a qual prevê o início do procedimento de privação no período entre 90 e 120 dias de idade, os parâmetros aqui apresentados possibilitam o aumento progressivo e controlado do peso do animal em privação, fazendo-o acompanhar, de modo eqüidistante, a curva teórica de crescimento (v. curva sugerida de privação, Figura 4). A implementação precisa desta sugestão envolveria a construção de um modelo matemático que, a partir dos pesos ad lib que antecedem ao início da privação, projetasse, para cada sujeito, individualmente, a sua própria curva de crescimento (ver, como exemplo, Oliveira, Lôbo, & Pereira, 2000). A partir dessa curva ad lib projetada, obter-se-ia uma curva de privação que, ao longo do experimento, representasse aumentos graduais e progressivos de peso que mantivessem tentativamente constante a porcentagem a que os pesos de privação, relativamente aos pesos ad lib projetados, se referem. A avaliação desse procedimento, obviamente, dependeria de uma investigação empírica do mesmo, por meio da qual medidas comportamentais do valor reforçador da água ou alimento fossem diretamente apreciadas.
Referências
Brownell, K. D., Greenwood, M. R. C., Stellar, E., & Shrager, E. E. (1986). The effects of repeated cycles of weight loss and regain in rats. Physiology & Behavior, 38, 459-464. [ Links ]
Carlton, P. L. (1961). The interacting effects of deprivation and reinforcement schedules. Journal of the Experimental Analysis of Behavior, 4, 379-381. [ Links ]
Conrad, D. G., Sidman, M., & Herrnstein, R. J. (1958). The effects of deprivation upon temporally spaced responding. Journal of the Experimental Analysis of Behavior, 1, 59-65. [ Links ]
Curi, R., Bazotte, R. B., Hell, N. S., & Timo-Iaria, C. (1989). Reversibility of metabolic changes induced by feeding schedule in rats. Physiology & Behavior, 45, 249-254. [ Links ]
Curi, R., Hell, S., Bazotte, R. B., & Timo-Iaria, C. (1984). Metabolic performance of free fed rats subjected to prolonged fast as compared to the metabolic pattern in rats under long term food restriction. Physiology & Behavior, 33, 525-531. [ Links ]
Davey, G. C. L., & Cleland, G. G. (1982). Topography of signal-centered behavior in the rat: Effects of deprivation state and reinforcer type. Journal of the Experimental Analysis of Behavior, 38, 291-304. [ Links ]
Denise, R. S. K., & Brinks, J. S. (1985). Genetic and environmental aspects of the growth curve parameters in beef cows. Journal of Animal Science, 61, 1431-1440. [ Links ]
Fischer, K., & Fantino, E. (1968). The dissociation of discriminative and conditioned reinforcing functions of stimuli with changes in deprivation. Journal of the Experimental Analysis of Behavior, 11, 703-710. [ Links ]
Iversen, I. H., & Lattal, K. A. (1991). Experimental Analysis of Behavior Amsterdam: Elsevier. [ Links ]
Leander, J. D. (1973). Effects of food deprivation on free-operant avoidance behavior. Journal of the Experimental Analysis of Behavior, 19, 17-24. [ Links ]
Leveille, G. A. (1970). Adipose tissue metabolism: influence of periodicity of eating and diet composition. Federation Proceedings, 29, 1294-1301. [ Links ]
Leveille, G. A., & Chakrabarty, K. (1968). Absorption and utilization of glucose by meal-fed and nibbling rats. Journal of Nutrition, 93, 69-75. [ Links ]
Lima, F. B., Hell, N. S., Timo-Iaria, C., Scivoletto, R., Dolnikoff, M. S., & Pupo, A. A. (1981). Metabolic consequences of food restriction in rats. Physiology & Behavior, 27, 115-123. [ Links ]
López-Espinoza, A. & Martínez, H. (2001). Efectos de dos programas de privación sobre el peso corporal y el consumo total de agua y comida en ratas. Acta Comportamentalia, 9, 1, 5-17. [ Links ]
Machado, U. F. (1989). Alteração da secreção de insulina em rato Wistar com 12 meses de idade. Unpublished Ph.D., Universidade de São Paulo, São Paulo. [ Links ]
Matos, M. A., & Tomanari, G. Y. (2002). A Análise Experimental do Comportamento no Laboratório Didático. São Paulo: Manole. [ Links ]
Michael, J. (1982). Distinguishing between discriminative and motivational functions of stimuli. Journal of the Experimental Analysis of Behavior, 37, 149-155. [ Links ]
Morrow, R. E., McLaren, J. B., & Butts, W. T. (1978). Effect of age on estimates of bovine growth-curve parameters. Journal of Animal Science, 47, 352-357. [ Links ]
Nadarajah, K., Marlowe, T. J., & Notter, D. R. (1984). Growth patterns of Angus Charolais, Charolais x Angus and Holstein x Angus cows from birth to maturity. Journal of Animal Science, 59, 957-966. [ Links ]
Oliveira, H. N., Lôbo, R. B., & Pereira, C. S. (2000). Comparação de modelos não-lineares para descrever o crescimento de fêmeas da raça Guzerá. Pesquisa Agropecuária Brasileira, 35, 9. [ Links ]
Pierce, W. D., Epling, W. F., & Boer, D. P. (1986). Deprivation and satiation: The interrelations between food and wheel running. Journal of the Experimental Analysis of Behavior, 46, 199-210. [ Links ]
Reese, E. P. (1967). Experiments in operant behavior. New York: Appleton-Century-Crofts. [ Links ]
Sidman, M. (1960). Tactics of scientific research. New York: Basic Books. [ Links ]
Recebido em: 02/04/03
Primeira decisão editorial em: 26/05/03
Versão final em: 27/06/03
Aceito em: 27/06/03
1 A realização deste trabalho contou com auxílios do CNPq por meio de bolsas Produtividade em Pesquisa concedidas aos primeiro e terceiro autores, bem como a bolsa de Iniciação Científica concedida ao segundo. Os autores agradecem as valiosas contribuições do Prof. César Ades e de Márcio Cruz. Este trabalho foi apresentado no 8o Simpósio Internacional de Iniciação Científica, realizado na Universidade de São Paulo em 2000. Endereço para correspondência: IPUSP - PSE. Av. Prof. Mello Moraes, 1721, São Paulo, SP, 05508-900.
2 Características Nutricionais segundo o fabricante: Proteína Bruta (max.) 23%; Gordura (max.) 4%; Fibra (max.) 5%; Cálcio (max.) 1,5%; Fósforo (min.) 0,85%; Cinzas (max.) 10%; Energia met. Kcal/kg 2750,00; Arginina % 1,5%; Isoleucina 1,0%; Lisina 1,4%; Metionina 0,4%; Cistina + Metionina 0,8%; Treonina 0,9%; Triptofano 0,3%; Vitamina A (UI/kg) 25000; Vitamina D3 (UI/kg) 4000; Vitamina E (UI/kg) 80,00; Menadiona (ppm) 6,4; Colina (ppm) 2800,00; Niacina (ppm) 220,00; Pantotenato de Cálcio(ppm) 90,00; Riboflavina (ppm) 12,00; Vitamina B12 (ppb) 40,00; Biotina (ppm) 0,20; Ácido Fólico (ppm) 13,00; Piridoxina (ppm) 11,00; Tiamina (ppm) 11,00; Magnésio (g/kg) 3,5; Potássio (mg) 9,00; Sódio (g/kg) 2,0; Ferro (ppm) 180,00; Cobalto (ppm) 2,0; Cobre (ppm) 30,0; Iodo (ppm) 1,0; Manganês (ppm) 110,00; Zinco (ppm) 110,00; Selênio (ppm) 0,2.
