










Services on Demand

article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators
Share

 Permalink
PermalinkUniversitas Psychologica
Print version ISSN 1657-9267
Univ. Psychol. vol.7 no.2 Bogotá Aug. 2008
Evidencia de priming gustativo y su disociación con la memoria explícita*
Evidence of gustative priming and its dissociation with the explicit memory
Eugenia Razumiejczyk; Guillermo Macbeth; Alfredo López-Alonso**
Instituto de Investigaciones Psicológicas de la Universidad del Salvador, Ciudad Autónoma de Buenos Aires, Argentina
ABSTRACT
The implicit processing of the senso-perceptive modality of the taste in superficial codification and its difference with the explicit memory is hypothesized in the present study 30 argentine subjects participated in this study whose age average was 22.47 years (SD = 3.01 years). In relation to the implicit memory, the results showed that after exhibition to the gustative stimuli, the subsequent perceptual processing is more efficient producing faster reaction times. The data also showed that the amount of stimuli recovered in the explicit memory was smaller than the total number of stimuli presented in the experiment. The task performance of free memory was not different between the studied stimuli and the new stimuli.
Keywords authors Memory, Priming, Neuropsychology, Perception, Taste.
Key words plus Neuropsychology, Memory, Perceptual disorders.
RESUMEN
En el presente estudio se pronostica el procesamiento implícito de la modalidad sensorio-perceptiva del gusto en codificación superficial y su diferencia con la memoria explícita. Participaron del estudio 30 sujetos argentinos cuya edad promedio resultó de 22,47 años (de = 3,01 años). En relación con la memoria implícita, los resultados mostraron que luego de la exposición al estímulo gustativo, el procesamiento perceptual subsecuente es más eficiente produciendo tiempos de reacción más rápidos. Así mismo, los datos mostraron que la cantidad de estímulos recuperados en la memoria explícita fue menor al número total de estímulos presentados en el experimento. El rendimiento de la tarea de recuerdo libre no se diferenció entre los estímulos estudiados y los estímulos nuevos.
Palabras clave autores Memoria, Priming, Neuropsicología, Percepción, Gusto.
Palabras claves descriptores Neuropsicologia, Memoria, Trastornos de la percepción.
Introducción
Los patrones de rendimiento en tareas de memoria fueron caracterizados como una disociación entre procesos de recuperación explícita e implícita (Ellis & Young, 1992; Graf & Schacter, 1985; Schacter, 1999).
La memoria explícita hace referencia a un tipo de memoria consciente que permite recordar experiencias previas (Ruiz-Vargas, 2000; Schacter, 1999). Puede ser tanto intencional como no intencional (Schott, Richardson-Klavehn, Heinze & Düzel, 2002; Schott et al., 2004, 2006). Este tipo de memoria es evaluado a través de medidas directas, tales como el recuerdo libre o el reconocimiento, en las que el participante conscientemente recupera el material presentado previamente. En la tarea de recuerdo libre, por ejemplo, se presenta una serie de estímulos y, luego de un cierto tiempo, se pide a los participantes que nombren aquellos que recuerdan. La tarea de reconocimiento, en cambio, consiste en presentar una serie de estímulos y luego se presentan los mismos estímulos junto con otros nuevos para que el participante reconozca los estímulos antiguos y los distinga de los nuevos (Baddeley, 1999; Ballesteros & García Rodríguez, 1998; Düzel, Vargha-Khadem, Heinze & Mishkin, 2001; Kosslyn & Rosenberg, 2004).
La memoria implícita, en cambio, es recuperación no intencional de la experiencia previa que, sin que el sujeto sea consciente de que está experimentando un recuerdo, influencia su comportamiento actual. El efecto de priming de repetición hace referencia a la influencia que tiene un estímulo en el rendimiento subsiguiente del sistema de procesamiento (Schacter, 1987, 1999; Schacter, Alapert, Savage, Rauch & Albert, 1996). Este tipo de memoria es evaluado a través de medidas indirectas (Baddeley, 1999; Ballesteros & García Rodríguez, 1998).
Numerosos investigadores señalan que el sistema de memoria que subyace a la memoria implícita es el Sistema de Representación Perceptual (Tulving & Schacter, 1990; Schacter, 1992). Este sistema hace referencia a una clase de subsistema de memoria de dominio específico que procesa y representa la información sobre la forma y la estructura, pero no sobre el significado u otras propiedades asociativas de las palabras, objetos u otros estímulos (Schacter, 1992). Así, además del sistema de palabra en forma visual, del sistema de descripción estructural y del sistema en forma auditiva, por implicación, cabe suponer que habrá otros subsistemas encargados de otras modalidades sensoriales, como la háptica (Ballesteros, Reales & Manga, 1999), la olfativa (Koenig, Bourron & Royet, 2000) y, posiblemente, también la gustativa.
El propósito de este estudio es explorar y probar si se produce efecto de priming de repetición del gusto según el reconocimiento y el tiempo de reacción en condición de codificación superficial y comparar los resultados con la memoria explícita, en recuerdo libre. El concepto de codificación superficial hace referencia al nivel de procesamiento que se efectúa solo en relación al estímulo físico (Craik & Lockhart, 1972).
En coherencia con los antecedentes teóricos y empíricos, se pronostica en el presente estudio el procesamiento implícito de la modalidad sensorioperceptiva del gusto en codificación superficial y su diferencia con la memoria explícita.
La primera hipótesis experimental H1 afirma que el reconocimiento será mayor para los estímulos gustativos estudiados Ee en la fase de estudio del experimento que para los estímulos nuevos En (H1: Ee > En).
La hipótesis H2 afirma que el tiempo de reacción será menor para los estímulos gustativos estudiados Ee en la fase de estudio del experimento que para los estímulos nuevos En (H2: Ee < En).
Las hipótesis H1 y H2 justifican la presencia del efecto de priming de repetición gustativo desde las variables del reconocimiento y del tiempo de reacción.
La hipótesis H3 afirma que en la tarea de recuerdo libre, el número de estímulos recuperados (Er) será menor que la totalidad de estímulos presentados en el experimento Et = 8 (H3: Er < Et).
La H4 afirma que los estímulos estudiados Ee en la fase de estudio del experimento y los estímulos nuevos En no presentarán diferencias en la tarea de recuerdo libre (H4: Ee = En).
Las hipótesis H3 y H4 se justifican por el tiempo transcurrido entre la presentación de los estímulos y su posterior evaluación, luego de administrar una actividad distractora.
Método
Participantes
La estrategia de muestreo utilizada para el reclutamiento de los participantes fue no probabilística de sujetos voluntarios.
Participaron del experimento 30 sujetos argentinos reclutados de cursos universitarios de grado de la Facultad de Psicología y Psicopedagogía de la Universidad del Salvador. La edad promedio de los participantes resultó de 22,47 años (de = 3,01 años). La población femenina (66,7%) predominó sobre la masculina (33,3%). Los participantes fueron no fumadores y se les informó que no debían ingerir ningún alimento ni bebida que no fuera agua durante las tres horas previas al experimento. Se estableció esta consigna durante el reclutamiento de participantes que se realizó una semana antes del estudio. Al inicio del experimento, se verificó el cumplimiento de estas condiciones. Ningún participante tuvo que ser excluido por este criterio.
Experimento
Se efectuó un estudio preliminar cuyo propósito fue evaluar el rendimiento de un grupo de participantes en tareas de reconocimiento para los estímulos gustativos de banana, pera, manzana, frutilla, ananá, durazno, zapallo, zanahoria, zapallito y brócoli. Se seleccionó un conjunto confiable de estímulos que resulten adecuados para poner a prueba las hipótesis correspondientes al experimento vinculado al efecto de priming de repetición gustativo. Se aplicó como criterio de inclusión a la probabilidad de acierto, de manera que cada estímulo no resulte demasiado fácil ni demasiado difícil de reconocer. Los resultados obtenidos permitieron identificar ocho estímulos gustativos adecuados para el estudio del priming de repetición: banana, pera, manzana, frutilla, ananá, durazno, zapallo y zapallito. Ninguno de ellos resultó ni demasiado fácil ni demasiado difícil para las tareas de reconocimiento. Se excluyeron la zanahoria y el brócoli dado que resultaron demasiado difíciles de reconocer, de modo que su no reconocimiento en el experimento se debería, en parte, a la dificultad misma del estímulo más que a la manipulación de las variables independientes.
Se entrenó a un grupo de seis alumnos voluntarios pertenecientes a la Facultad de Psicología y Psicopedagogía de la Universidad del Salvador que cursaban al momento del estudio (2006) la materia Procesos Básicos IV cuyo profesor titular es el Dr. Alfredo O. López Alonso. Los voluntarios no conocieron el propósito del estudio hasta finalizada la toma de datos, de modo que funcionaron como experimentadores ciegos.
La agrupación de los estímulos gustativos en estímulos estudiados y estímulos nuevos fue aleatoria y se obtuvo mediante un programa estadístico para computadoras personales.
El experimento fue llevado a cabo en una de las oficinas del Instituto de Investigaciones Psicológicas de la Universidad del Salvador (IIPUS). Cada estudio fue realizado por dos experimentadores que debían tener el pelo recogido por razones de higiene.
Sobre un escritorio se ubicaban los ocho recipientes tapados, cada uno con un estímulo gustativo y, sobre ellos, un mantel que los cubría de la vista del participante al momento de ingresar a la sala. A un costado se encontraba un bote de basura cuya función era la de contener los materiales ya utilizados.
El experimentador 1 pedía el ingreso del participante y constataba que hubiera cumplido con los dos requisitos para el estudio, es decir, ser no fumador y no haber ingerido ningún alimento ni bebida que no fuera agua durante las últimas tres horas. Le solicitaba que tomara asiento frente al escritorio y explicaba el procedimiento de la fase de estudio del experimento enfatizando que la totalidad de los materiales utilizados eran descartables. Su propósito era que el participante supiera cómo se iba a proceder, garantizarle la higiene del estudio y otorgarle la oportunidad de rechazar el formar parte del mismo. El experimentador 1 pedía información acerca de su edad y sexo que el experimentador 2 completaba en el protocolo de respuestas. Luego, el experimentador 1 entregaba al participante una servilleta de papel y un vaso descartable con agua, recordándole que debía beber unos sorbos antes de cada ensayo.
En la fase de estudio, el experimentador 1 presentaba al participante cada uno de los cuatro estímulos gustativos en forma de papilla y a temperatura ambiente que debía degustar y realizar una codificación superficial, es decir, informar si el gusto que percibía era salado o dulce. Los estímulos presentados en esta fase fueron pera, manzana, zapallo y durazno.
El procedimiento de la fase de estudio del experimento consistía en que el experimentador 2 retiraba el mantel que cubría los recipientes de los estímulos de forma tal que lo sostenía como cortina, con el objetivo de que el participante no los observara. El experimentador 1 destapaba un recipiente que contenía el estímulo, lo servía en una cucharita de té descartable que solo se utilizaba para ese ensayo y volvía a tapar el recipiente. Luego, informaba al participante que iba a recibir en su boca un alimento. El participante respondía acerca de la cualidad del gusto, es decir, si éste era salado o dulce. El experimentador 2 registraba estos datos en el protocolo de respuestas. El experimentador 1 tiraba la cucharita utilizada para ese ensayo. Se informaba al participante que realizara una limpieza bucal ingiriendo unos sorbos de agua. Se consignaba que pusiera agua en su boca, enjuagara y tragara. Del mismo modo, se continuaba con los tres estímulos restantes.
A continuación, el experimentador 1 entregaba al participante una tarea de papel y lápiz. Esta actividad funcionaba como tarea distractora. El tiempo para responder a las preguntas que contenían la tarea era de aproximadamente 20 minutos. Al finalizar la tarea, se proseguía con la fase de prueba del experimento. El experimentador 1 explicaba el procedimiento de la fase de prueba con el mismo propósito que en la fase anterior. El experimentador 1 solicitaba al participante que bloqueara su visión con un antifaz. El experimentador 2 retiraba el mantel que cubría los recipientes. El procedimiento fue similar al de la fase de estudio, aunque aquí se entregaban los cuatro estímulos previos (pera, manzana, zapallo y durazno) más cuatro estímulos nuevos (frutilla, ananá, banana y zapallito). El participante debía reconocer el estímulo gustativo en el menor tiempo posible. Se tomó el tiempo en segundos desde el momento en que el estímulo era entregado en la boca del participante hasta que éste daba una respuesta. El experimentador 1 utilizó un cronómetro analógico Heuer para medir el tiempo de reacción en segundos. La tarea del experimentador 2 consistía en registrar los datos del estudio en el protocolo de respuestas.
Los alimentos fueron entregados en un orden aleatorio que se obtuvo mediante un programa estadístico para computadoras personales.
El experimentador 2 cubría los recipientes con el mantel. El experimentador 1 retiraba el antifaz de los ojos del participante y le preguntaba cuáles fueron los estímulos presentados en el experimento, es decir, se efectuaba una tarea de recuerdo libre. El participante nombraba los gustos que recordaba mientras el experimentador 2 los registraba en el protocolo de respuestas.
Al finalizar el experimento, el experimentador 1 agradecía al participante la colaboración y lo acompañaba a la puerta de salida. Se arrojaban al bote de basura el vaso descartable y la servilleta de papel utilizados. Al concluir la jornada de experimentos, los experimentadores arrojaban, además, la totalidad de los recipientes que contenían los estímulos gustativos.
Resultados y discusión
La media del reconocimiento para los estímulos estudiados en la fase de estudio del experimento resultó de 2,97 (desviación estándar [DE] = 1,12). La distribución de esta variable resultó compatible con la presunción de normalidad y homocedasticidad por las pruebas de Kolmogorov-Smirnov y Levene, respectivamente.
La media del reconocimiento para los estímulos nuevos resultó de 2,53 (DE = 1,16). La distribución de esta variable también fue compatible con la presunción de normalidad y homocedasticidad.
Con el fin de comparar el reconocimiento de los estímulos estudiados y los estímulos nuevos, se efectuó una prueba t para muestras apareadas. La diferencia de medias para estas variables resultó significativa (t = 2,282; p = 0,03). Así, los resultados obtenidos en el reconocimiento de estímulos estudiados y de estímulos nuevos no se debió al azar sino que los participantes mostraron un mejor reconocimiento cuando los estímulos gustativos fueron presentados en la fase de estudio. Los resultados sugieren que se efectuó un proceso de facilitación en el reconocimiento de los estímulos estudiados en la fase de estudio, es decir, un efecto de priming de repetición gustativo en relación al reconocimiento. El análisis de estos datos sugiere el rechazo de la hipótesis nula correspondiente a la H1.
La hipótesis H2 resultó coherente con la evidencia experimental. La media del tiempo de reacción para los estímulos estudiados en la fase de estudio del experimento resultó de 33,27 segundos (DE = 17,31 segundos) mientras que la media del tiempo de reacción de los estímulos nuevos fue de 40,73 segundos (DE = 19,75 segundos). Con el fin de comparar el tiempo de reacción de las respuestas de los participantes en los estímulos estudiados y en los estímulos nuevos, se efectuó una prueba t para muestras apareadas. Los resultados mostraron la presencia de una diferencia significativa en el tiempo de reacción (t = -2,410; p = 0,023). Así, el tiempo de reacción fue significativamente menor cuando los estímulos habían sido presentados en la fase de estudio. Las variables comparadas resultaron normales y homocedásticas según las pruebas de Kolmogorov-Smirnov y Levene, respectivamente. Los resultados sugieren que se ha producido un efecto de priming de repetición gustativo debido a la facilitación del procesamiento de los estímulos estudiados en la fase de estudio del experimento en relación al tiempo de respuesta. El análisis de estos datos sugiere el rechazo de la hipótesis nula correspondiente a la H2.
El análisis conjunto de los datos se encuentran en concordancia con estudios previos que mostraron que luego de la exposición al estímulo, el procesamiento perceptual subsecuente es más eficiente produciendo tiempos de reacción más rápidos (Buckner, Petersen, Ojeman, Meizin, Squire & Raichle, 1995; Buckner et al., 1998; Henson, 2003; Schacter, 1999; Schott et al., 2004, 2006; Tulving & Schacter, 1990; Squire, 1987; Squire, Ojemann, Meizin, Petersen, Videen & Raichle, 1992; Wig, Grafton, Demos & Kelley, 2005; Wagner, Maril & Schacter, 2000).
Se estudiaron los resultados de la memoria explícita correspondiente a la tarea de recuerdo libre. Los datos mostraron que la media total de estímulos recuperados fue de 5,10 (DE = 1,56) con una mediana de 5 y un modo de 6. Así, la cantidad de estímulos recuperados en la memoria explícita fue menor al número total de estímulos presentados en el experimento, es decir, ocho estímulos. Estos resultados se presentan en la Figura 1
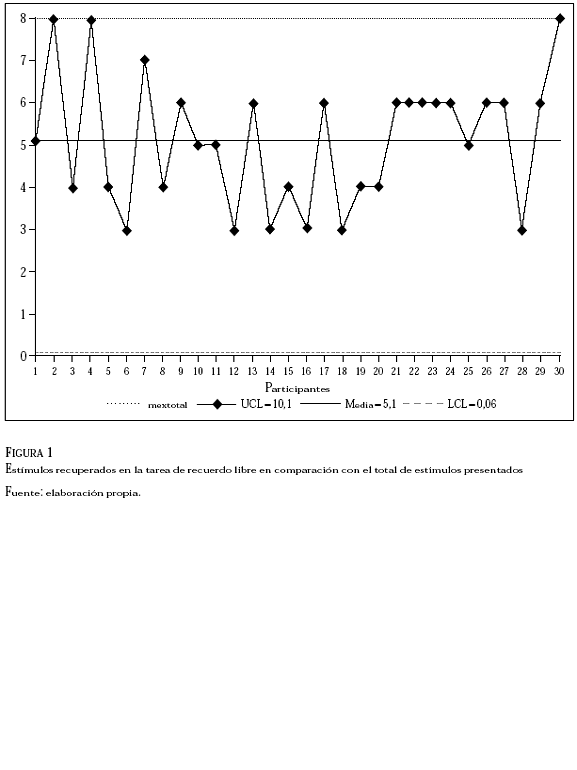
En la ordenada se ubica la cantidad de estímulos gustativos, mientras que la cantidad de participantes se ubica en la abscisa. Cada punto en el gráfico representa la cantidad de estímulos recuperados en la tarea de recuerdo libre por cada participante, de modo que el total de puntos es de 30 (n = 30). La media de los estímulos recuperados es de 5,10 (DE = 1,56). Así, la media de los estímulos recuperados fue menor que el total de los estímulos gustativos pertenecientes al experimento. Se puede apreciar en la figura 1 que tres participantes lograron recuperar de su memoria explícita, la totalidad de los estímulos, esto es, los ocho gustos.
Estos resultados son consistentes con estudios previos en los que se manipuló el intervalo de retención y recuperación de estímulos. Los resultados mostraron que el decaimiento en la recuperación de la memoria explícita resultó acelerado (Cave & Squire, 1992; Tulving & Schacter, 1990). El análisis de estos datos sugiere el rechazo de la hipótesis nula correspondiente a la H3.
La hipótesis H4 resultó coherente con la evidencia experimental. Los resultados mostraron que el recuerdo libre para los estímulos estudiados obtuvo una media de 2,73 (DE = 0,94) y una mediana de 3. La media del recuerdo libre de los estímulos nuevos fue de 2,37 (DE = 1,15) y una mediana de 2,50. La distribución de las variables resultó incompatible con la presunción de normalidad y homocedasticidad por las pruebas de Kolmogorov-Smirnov y Levene, respectivamente. El predominio del recuerdo libre para los estímulos estudiados sobre el recuerdo libre para los estímulos nuevos resultó no significativo por la prueba de Wilcoxon (p = 0,158). El rendimiento de la tarea de recuerdo libre no se diferenció entre los estímulos estudiados y los estímulos nuevos. Estos resultados son coherentes con los estudios de Craik & Lockhart (1972), Craik & Tulving (1975), Lockhart & Craik (1990) y Tulving, Kapur, Markowitsch, Craik & Houle (1994) que mostraron que la codificación superficial de los estímulos produce un peor aprendizaje que cuando esa misma información se procesa a un nivel profundo. Al ser los estímulos estudiados codificados a nivel superficial, no se diferenciaron de los estímulos nuevos, en concordancia con la H4.
Conclusiones
Se ha estudiado el priming de repetición de la modalidad sensorio-perceptiva del gusto en condición de codificación superficial y se han comparado los resultados obtenidos con el rendimiento de la memoria explícita evaluada a través de una tarea de recuerdo libre.
Las hipótesis de trabajo H1 y H2 intentaron evaluar el priming de repetición gustativo a través de dos variables, esto es, el reconocimiento y el tiempo de reacción. Ambas hipótesis resultaron coherentes con la evidencia experimental. El reconocimiento de los estímulos gustativos estudiados en la fase de estudio resultó mayor que el reconocimiento de los estímulos nuevos presentados en la fase de prueba, necesitando menos tiempo para el procesamiento de los estímulos estudiados. Así, se produjo una facilitación tanto en el reconocimiento, como en el tiempo de procesamiento requerido en los estímulos gustativos estudiados. El análisis conjunto de estos resultados sugiere que se ha producido un priming de repetición en la modalidad sensorio-perceptiva del gusto.
Así mismo, la administración de una tarea distractora entre la fase de estudio y la fase de prueba no produjo un decaimiento significativo de la recuperación implícita de los estímulos estudiados. Estos resultados se encuentran en concordancia con estudios anteriores. La evidencia experimental ha mostrado que el efecto de priming de repetición puede durar intervalos de días, semanas o meses para la lectura de texto invertido (Kolers, 1976) y para el desempeño de completar raíces de palabras (Sloman, Hayman, Otha, Law & Tulving, 1988).
Las hipótesis de trabajo H3 y H4 también resultaron coherentes con la evidencia experimental. Por un lado, la cantidad de estímulos recuperados en memoria explícita, evaluada a partir de la tarea de recuerdo libre, resultó menor al número total de estímulos presentados en el experimento. Asimismo, el predominio del recuerdo libre para estímulos estudiados por sobre el recuerdo libre para estímulos nuevos resultó no significativo. En la memoria explícita, los estímulos estudiados y los nuevos no se diferenciaron en el rendimiento de esta tarea. Si bien los resultados mostraron que los estímulos estudiados tuvieron una mejor recuperación media en memoria explícita (m = 2,73; DE = 0,94) en comparación con los estímulos nuevos (m = 2,37; DE = 1,15), esta diferencia se debió al azar.
De este modo, el paso del tiempo entre la adquisición y la recuperación tiene una incidencia desigual en la memoria explícita e implícita (Froufe, 1997). En las disposiciones experimentales en las que se manejó el intervalo de retención, el decaimiento en la recuperación de la memoria explícita resultó acelerado, mientras que el efecto de priming de repetición pudo persistir con poca pérdida durante horas, días (Cave & Squire, 1992) y hasta meses (Tulving & Schacter, 1990).
Estos resultados recomiendan investigar si se produce efecto de priming de repetición del gusto según el reconocimiento y el tiempo de reacción en condición de codificación profunda y comparar los resultados con la memoria explícita en recuerdo libre.
Referencias
Baddeley, A. (1999). Memoria Humana. Teoría y práctica. Barcelona: McGraw-Hill. [ Links ]
Ballesteros, S. & García Rodríguez, P. (1998). Procesos Psicológicos Básicos. Madrid: Universitas. [ Links ]
Ballesteros, S., Reales, J. & Manga, D. (1999). Memoria implícita y memoria explícita intramodal e intermodal: influencia de las modalidades elegidas y del tipo de estímulos. Psicothema, 11, 831-851. [ Links ]
Buckner, R. L., Petersen, S. E., Ojemann, J. G., Meizin, F. M., Squire, L. R. & Raichle, M. E. (1995). Functional anatomical studies of explicit and implicit memory retrieval tasks. Journal of Neuroscience, 15, 12-29. [ Links ]
Buckner, R. L., Goodman, J., Burock, M., Rotte, M., Koutstaal, W., Schacter, D. L., Rosen, B. & Dale, A. M. (1998). Functional-anatomic correlates of object priming in humans revealed by rapid presentation event-related fMRI. Neuron, 20, 285–296. [ Links ]
Cave, C. R. & Squire, R. L. (1992). Intact and longlasting repetition priming in amnesia. Journal of Experimental Psychology: Learning, Memory and Cognition, 18, 509-520. [ Links ]
Craik, F. I. M. & Lockhart, R. S. (1972). Levels of processing: A framework for memory reseach. Journal of Verbal Learning and Verbal Behavior, 11, 671- 684. [ Links ]
Craik, F. I. M. & Tulving, E. (1975). Depth of processing and the retention of words in episodic memory. Journal of Experimental Psychology, 104, 268-294. [ Links ]
Düzel, E., Vargha-Khadem, F., Heinze, H. J. & Mishkin, F. (2001). Brain activity evidence for recognition without recollection after early hippocampal damage. Proceedings of the National Academy of Sciences, 98(14), 8101-8106. [ Links ]
Ellis, A. & Young, A. (1992). Neuropsicología cognitiva humana. Barcelona: Masson. [ Links ]
Froufe, M. (1997). El inconsciente cognitivo. Madrid: Biblioteca Nueva. [ Links ]
Graf, P. & Schacter, D. L. (1985). Implicit and explicit memory for new associations in normal and amnesic subjects. Journal of Experimental Psychology, 11, 501-518. [ Links ]
Henson, R. N. (2003). Neuroimaging studies of priming. Neurobiology, 70, 53-81. [ Links ]
Koenig, O., Bourron, G. & Royet, J.-P. (2000). Evidence for separate perceptive and semantic memories for odors: a priming experiment. Chemical Senses, 25, 703-708. [ Links ]
Kolers, P. A. (1976). Specificity of operations in sentence recognition. Cognitive Psychology, 1, 289-306. [ Links ]
Kosslyn, S. & Rosenberg, R. (2004). Psychology. The brain, the person, the world. Needlam Heights, MA: Allyn & Bacon. [ Links ]
Lockhart, R. S. & Craik, F. I. M. (1990). Levels of processing: A retrospective commentary on a framework of research. Canadian Journal of Psychology, 44, 87-112. [ Links ]
Ruiz-Vargas, J. M. (2000). La organización neurocognitiva de la memoria. Barcelona: Anthropos. [ Links ]
Schacter, D. L. (1987). Implicit memory: History and current status. Journal of Experimental Psychology, Learning, Memory and Cognition, 13, 501-518. [ Links ]
Schacter, D. L. (1992). Priming and multiple memory systems: perceptual mechanisms of implicit memory. Journal of Cognitive Neurosciences, 4, 244- 256. [ Links ]
Schacter, D. L. (1999). En busca de la memoria. El cerebro, la mente y el pasado. Barcelona: SineQuaNon. [ Links ]
Schacter, D. L., Alapert, N. M., Savage, C. R., Rauch, S. L. & Albert, M. S. (1996). Conscious recollection and the human hippocampal formation: Evidence from positron emission tomography. Proceedings of the National Academy of Sciences, 93, 321-325. [ Links ]
Schott, B., Richardson-Klavehn, A., Heinze, H. J. & Düzel, E. (2002). Perceptual priming versus explicit memory: Dissociable neural correlates at encoding. Journal of Cognitive Neuroscience, 14, 578-592. [ Links ]
Schott, B., Henson, R. N., Richardson-Klavehn, A., Becker C., Thoma, V., Heinze, H. J. & Düzel, E. (2004). Redefining implicit and explicit memory: The functional neuroanatomy of priming, remembering, and control of retrieval. Proceedings of the National Academy of Sciences, 102, 1257-1262. [ Links ]
Schott, B., Richarson-Klavehn, A., Henson, R. N. A., Becker, C., Heinze, H. J. & Düzel, E. (2006). Neuroanatomical dissociation of encoding processes related to priming and explicit memory. The Journal of Neuroscience, 26, 792-800. [ Links ]
Sloman, S. A., Hayman, C. A. G., Ohta, N., Law, J. & Tulving, E. (1988). Forgetting in primed fragment completion. Journal of Experimental Psychology, 14, 223-239. [ Links ]
Squire, L. R. (1987). Memory and brain. New York: Oxford University Press. [ Links ]
Squire, L. R., Ojemann, J. G., Meizin, F. M., Petersen, S. E., Videen, T. O. & Raichle, M. E. (1992). Activation of the hippocampus in normal humans: A functional anatomical study of memory. Proceedings of the National Academy of Sciences, 89, 1837-1841. [ Links ]
Tulving, E. & Schacter, D. L. (1990). Priming and human memory systems. Science, 247, 301-306. [ Links ]
Tulving, E., Kapur, S., Markowitsch, H. J., Craik, F. I. M., Habib, R. & Houle, S. (1994). Neuroanatomical correlates of retrieval in episodic memory: Auditory sentence recognition. Proceedings of the National Academy of Sciences, 91, 2012-2015. [ Links ]
Wagner, A. D., Maril, A. & Schacter, D. L. (2000). Interactions between forms of memory: when priming hinders new episodic learning. Journal of Cognitive Neuroscience, 12, 52-60. [ Links ]
Wig, G. S., Grafton, S. T., Demos, K. E. & Kelley, W. M. (2005). Reductions in neural activity underlie behavioral components of repetition priming. Nature Neuroscience, 8, 1228-1233. [ Links ]
Recibido: mayo 23 de 2007
Revisado: abril 10 de 2008
Aceptado: abril 20 de 2008
*Artículo de investigación financiado por el Consejo Nacional de Investigaciones Técnicas y Científicas (CONICET) de Argentina.
**Los autores pertenecen al Instituto de Investigaciones Psicológicas de la Universidad del Salvador. Marcelo T. de Alvear 1314 (C1058AAV), Ciudad Autónoma de Buenos Aires, Argentina. Correos electrónicos: eugeniaraz@fibertel.com.ar, guimacbeth@hotmail.com, alalonso@ciudad,com,ar

