










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.4 n.1-2 São Paulo 1993
ARTIGOS ORIGINAIS
A modularidade da memória e o sistema nervoso1
Modularity of memory and the nervous system
Gilberto Fernando Xavier
Instituto de Biociências - Departamento de Fisiologia - Universidade de São Paulo
RESUMO
A memória parece não ser uma entidade única, mas compreender conjuntos de habilidades mediadas por diferentes módulos do sistema nervoso. Não obstante a profunda divisão de tarefas e de processamentos, a cooperação entre os diferentes módulos proporciona uma sensação unificada. Cada um dos módulos possui conexões diferentes em relação aos outros e, também, seu nível de independência parece variar. Lesões em módulos independentes eliminam as funções por eles desempenhadas, enquanto lesões em módulos que cooperam resultam numa alteração de desempenho dos módulos remanescentes de modo a minimizar as deficiências. Esse funcionamento independente porém cooperativo entre os diferentes módulos permite explicar não só como determinados tipos de lesões cerebrais podem levar a uma profunda amnésia acompanhada da integridade de outras funções de memória, mas também a sensação de uma experiência de memória única em indivíduos normais. Dados sobre a identificação de circuitos do sistema nervoso relacionados aos diferentes módulos de memória são analisados.
Descritores: Memória. Cérebro. Distúrbios cerebrais. Amnésia. Aprendizagem.
ABSTRACT
Memory seems not to be a single entity but a pool of several abilities mediated by distinct brain systems. Examination of available evidence allows distinction between short-term memory, working memory and long-term memory. Besides the existing task division into modules, which involves several processing units working in parallel, the final result is a unitary memory experience. Each module of memory maintains extensive connections with the others, and variable levels of independence between them are maintained. Lesions of independent modules break up their function, while lesions in modules which cooperate intimately produce an alteration of functioning in the unaffected ones in order to minimize deficiencies. This independent but cooperative functioning of modules can explain experimental dissociations after lesioning, and also the unitary memory experience in normal subjects. Data on lesioning of neural circuits that underly some modules of memory are analysed.
Index Terms: Memory. Brain. Brain disorders. Amnesia. Learning.
Funções dissociáveis representam significativa evidência sobre a organização do sistema nervoso, proporcionando informações para a pesquisa básica em neurociências relacionada aos substratos neurais e arquitetura deste sistema.
A memória, capacidade de alterar o comportamento em função de experiências anteriores, depende de estruturas localizadas em diferentes regiões do sistema nervoso; o processamento de informações nessas estruturas neurais resulta em alterações nestas e/ou em outras regiões, levando a alterações no funcionamento do sistema.
Até recentemente, havia certo consenso entre investigadores da área de neuropsicologia (e.g., Mesulam, 1990), psicologia cognitiva (e.g., Baddeley, 1992) e aprendizagem animal (e.g., Collier, Quirk & Routtenberg, 1987) de que é útil, e mesmo necessário, postular a existência de diversos sistemas de memória.
Olton, um dos maiores defensores dessa idéia, expressou:
Memórias são como chocolates. Elas possuem diferentes formas e características, e contêm diferentes tipos de informação. Assim, a tarefa do neurocientista não é investigar o substrato neural singular de uma memória monolítica, mas identificar cada um dos diferentes sistemas neurais subjacentes a cada um dos tipos de memória (Olton, 1983, p.335).
Todavia, o próprio Olton (1989) questionou a existência de diversos sistemas de memória em artigo posterior:
A tese deste ensaio é simples. Ela advoga o exame da memória em termos de diferentes dimensões, ao invés de diferentes sistemas (p.l).
Nesse artigo, Olton (1989) questiona a lógica que levou à proposta de diferentes sistemas de memória e propõe uma análise dimensional envolvendo contingências temporais e procedimentos de treinamento que teria, segundo ele próprio, maior valor heurístico para a análise dos processos e mecanismos responsáveis pela memória.
A. Múltiplos sistemas de memória vs sistema único
Admite-se que as distinções conceituais sobre sistemas de memória refiram-se a unidades funcionais individuais, cada qual com características e processos diferentes da outra. A distinção inicialmente feita entre memória e hábito, por exemplo, teve por base diferenças nas taxas de aquisição de diferentes comportamentos, como aprender a datilografar e lembrar-se de um quadro após uma única exposição ao mesmo. Todavia, a maioria das evidências atuais que favorecem distinções entre tipos de memória advém de dissociações em indivíduos com disfunções neurais.
1. Evidências de estudos de lesões
Lesões mediais no lobo temporal ou em estruturas diencefálicas mediais produzem uma síndrome amnésica.
O caso de síndrome amnésica melhor conhecido é o de H.M., um paciente que sofria de epilepsia severa, e que teve removidos o giro parahipocampal, córtex entorrinal, amígdala e dois terços anteriores do hipocampo bilateralmente, numa tentativa de controlar os ataques associados com a epilepsia (Scoville & Milner, 1957). O resultado desse tratamento foi uma profunda amnésia anterógrada (também observada, posteriormente, em animais com lesões equivalentes), isto é, H.M. era incapaz de formar novas memórias para eventos ocorridos depois da cirurgia, muito embora suas funções intelectuais e de linguagem estivessem intactas. Por exemplo, se uma pessoa que estivesse desenvolvendo uma conversa com ele saísse da sala por alguns minutos, ao retornar verificaria que H.M. havia esquecido o conteúdo da conversa e mesmo do fato de estarem conversando. Todavia, era capaz de reconhecer por fotografia pessoas que ficaram famosas até 3 anos antes de cirurgia, bem como de lembrar episódios autobiográficos de sua adolescência.
Embora a lesão produzida no paciente H.M. tenha envolvido diversas estruturas, os efeitos amnésicos parecem estar associados à lesão hipocampal: pacientes com lesões restritas ao campo CA1 do hipocampo em decorrência de isquemia cerebral transitória (Zola-Morgan, Squire & Amaral, 1986), e pacientes com anormalidades hipocampais decorrentes de encefalite viral (Press, Amaral & Squire, 1989) exibem uma síndrome amnésica similar.
A amnésia em pacientes com síndrome de Korsakoff (portadores de lesões diencefálicas bilaterais nos núcleos talâmicos mediais, corpos mamilares e cerebelo, além de atrofia cortical) parece diferir, ao menos em parte, daquela exibida pelos pacientes hipocampais. Em ambos os casos, os pacientes conservam suas capacidades intelectuais e têm profunda amnésia; diferentemente dos pacientes hipocampais, os pacientes com síndrome de Korsakoff contudo exibem uma amnésia retrógrada que estende-se a décadas. Não está claro, todavia, se essa amnésia "retrógrada" decorre de amnésia anterógrada devido ao lento desenvolvimento da síndrome, uma vez que a severidade do distúrbio é progressivamente menor quanto mais remotos os eventos a serem lembrados.
É interessante notar que esses pacientes amnésicos são capazes de adquirir determinados tipos de informação da mesma forma que indivíduos normais, sugerindo que algumas funções de memória independem das regiões cerebrais afetadas nestas síndromes amnésicas.
A memória de curto prazo, por exemplo, parece intacta nestes pacientes, indicando que eles são capazes de reter pequenas quantidades de informação por curtíssimos períodos de tempo, mas têm dificuldades em mantê-las ou evocá-las depois que elas deixam de estar presente na experiência consciente.
Há, no entanto, funções de memória de longo prazo que estão preservadas em pacientes amnésicos. Por exemplo, H.M. e outros pacientes amnésicos exibem considerável retenção de habilidades motoras, perceptuais e cognitivas (para revisão ver Cohen, 1984). Mesmo o condicionamento clássico aversivo pode ser adquirido por pacientes portadores da síndrome de Korsakoff, sem que os mesmos estejam conscientes do fato (Daum, Channon & Canavan, 1989). A pré-ativação (priming) (facilitação inconsciente ou viés no desempenho em decorrência de material apresentado recentemente) também está preservada em pacientes amnésicos; por exemplo, exibem pré-ativação para completar palavras e em testes de associação livre, o que não ocorre com pacientes portadores de Alzheimer. Os efeitos de pré-ativação talvez dependam de áreas corticais comprometidas na síndrome de Alzheimer.
Esses tipos de dissociações, i.e., severos prejuízos em um tipo de tarefa em contraposição a um desempenho normal em outras tarefas, são apontados como evidência da existência de diferentes sistemas de memória. Cabe, no entanto, analisar como essas dissociações seriam possíveis no caso da existência de um único sistema de memória com diferentes dimensões mnêmicas, como proposto por Olton (1989).
Por um lado, pode-se imaginar que a memória em indivíduos lesados é fraca devido a falhas na codificação, deficiências no armazenamento ou, ainda, evocação ineficiente, de modo que apenas testes mais sensíveis (que avaliam aspectos particulares da memória), poderiam evidenciar efeitos de experiência prévia nestes indivíduos. "Diferentes processamentos" de memória poderiam levar a dissociações simplesmente tornando a memória fraca. Por exemplo, Meudell e Mayes (1981) testaram indivíduos normais muito tempo após a aprendizagem. Através dessa manipulação, eles reproduziram uma dissociação entre o desempenho facilitado e o reconhecimento impedido, normalmente observada em pacientes amnésicos testados imediatamente após a aprendizagem. Assim, a dissociação entre facilitação de desempenho e memória de reconhecimento impedida, estaria, nesse caso, relacionada à sensibilidade diferencial das medidas utilizadas aos diferentes aspectos da memória -ela será observada sempre que a memória esteja fraca, seja no paciente amnésico testado imediatamente após a aquisição, ou no indivíduo normal testado depois de um longo período da aquisição.
Por outro lado, é possível encontrar exemplos de ausência de prejuízos de memória em indivíduos lesados em situações cuja demanda é baixa, e subseqüente aparecimento de prejuízo com aumento da demanda. Por exemplo, numa situação de discriminação condicional com retardo (ver abaixo), a demanda da tarefa pode ser variada (1) pelo aumento do intervalo de tempo entre o momento de aquisição da informação e o momento de teste (que corresponde a verificar que o indivíduo retém a informação), ou (2) pelo aumento no número de informações apresentadas antes que se passe à fase de teste. Macacos com lesões em estruturas do lobo temporal exibem desempenho similar ao dos animais-controle quando a demanda deste tipo de tarefa é pequena (intervalo curto -segundos - associado à apresentação de uma única informação antes do teste); todavia, o aumento da demanda da tarefa (intervalo longo - minutos - associado a apresentação de 10 informações antes do primeiro teste) resulta em severos prejuízos no grupo lesado em relação ao controle (Mishkin, 1978; Murray & Mishkin, 1986). Resultados similares foram obtidos em outras situações comportamentais (e.g., Horel, Voytko & Salsbury, 1984; Zola-Morgan & Squire, 1985).
Este tipo de resultado sugere que a importância dos parâmetros da tarefa é relativa. Ao invés de diferentes sistemas de memória, com prejuízo em um deles, pode-se falar em problemas no sistema de codificação do contexto temporal da informação a ser lembrada. Algumas informações, por estarem fortemente associadas a esse contexto temporal, seriam mais passíveis de interferência, enquanto outras, menos associadas ao contexto temporal, seriam menos passíveis de interferência. Ou seja, fala-se num sistema único de memória com diferentes dimensões, sendo a temporal uma delas.
O mesmo raciocínio não se aplica, no entanto, quando os resultados experimentais apontam para a existência de uma dupla dissociação. Isto é, quando lesões em determinados sistemas neurais resultam em prejuízos no desempenho de uma dada tarefa "A", deixando intacto o desempenho numa segunda tarefa "B", enquanto um segundo tipo de lesão produz o resultado inverso, ou seja, prejuízo na tarefa "B" e desempenho normal na tarefa "A".
Neste contexto, é interessante notar que lesões combinadas do hipocampo e amígdala prejudicam o desempenho em tarefas de discriminação condicional com retardo, mas não a aprendizagem em tarefas de discriminação visual, enquanto lesões do tronco encefálico não interferem na discriminação condicional, mas impedem a discriminação visual (Zola-Morgan, Squire & Mishkin, 1982). Uma vez que os processos cognitivos envolvidos no desempenho destas tarefas são diferentes, essa dupla dissociação favorece fortemente a postulação de sistemas neurais distintos subjacentes a estes processos.
2. O argumento evolutivo
Sherry e Schacter (1987) desenvolveram uma argumentação baseada na idéia de incompatibilidade funcional entre as demandas ambientais e as propriedades dos sistemas de memória: aprendizagem e memória são especializações adaptativas que lidam com problemas específicos do animal no seu ambiente; adaptações que servem a uma função não podem, pela sua natureza especializada, servir de forma efetiva a uma outra função. Em outras palavras, as propriedades específicas que fazem um sistema ser efetivo para a solução de um dado problema também fazem com que ele seja incompatível com as demandas de outros problemas. Isto, no entanto, não significa que este sistema não seja capaz de lidar com uma grande variedade de problemas.
Neste contexto, pode-se admitir que diferentes sistemas de memória podem ser caracterizados por regras de operação fundamentalmente diferentes e, talvez, pelo tipo de informação que manipulam. Essa idéia parte do pressuposto de que a organização cognitiva se dá de forma modular, representada por sistemas paralelos de processamento de informações . Cada módulo atuaria em seu domínio específico com regras particulares, de forma independente e paralela em relação a outros módulos.
B. Distinções dicotômicas
Várias distinções dicotômicas entre tipos de memória foram feitas. Por exemplo, memória de curto e de longo prazo (Baddeley & Warrington, 1970), memória de referência e operacional (Honig, 1978), memória disposicional e representacional (Thomas, 1984), memória semântica e episódica (Tulving, 1972, 1983), memória semântica e cognitiva (Warrington & Weiskrantz, 1982), sistemas para memória e hábitos (Hirsh, 1974; Mishkin & Petri, 1984), memória procedimental e declarativa (Cohen & Squire, 1980; Cohen, 1984), "saber como" e "saber que" (Ryle, 1949), memória recente e remota (Schacter & Moscovitch, 1984), memória implícita e explícita, entre outras. Discutiremos aqui apenas as distinções que têm maior apoio experimental.
1. Memórias de curto e de longo prazo
A mais antiga de todas as dicotomias distingue entre memória de curto e de longo prazo.
Memória de curto prazo refere-se à capacidade de armazenar pequena quantidade de informações por período de tempo limitado (Baddeley & Warrington, 1970); nesse período, a informação é mantida por repetição no sistema de memória. Pacientes com danos nos giros supramarginal e angular do hemisfério esquerdo exibem uma memória verbal auditiva de curto prazo limitada, mas uma memória de longo prazo quase normal. A duração de sua memória para dígitos, palavras ou letras apresentadas auditivamente é muito reduzida; em tarefas que envolvem lembrança intencional, usualmente recordam-se apenas do primeiro item de uma lista de dez itens. Desempenham-se, contudo, normalmente em tarefas que envolvem associação entre pares de palavras (nos quais uma palavra é usada como "dica" para a recordação da outra) e estórias. Esse tipo de deficiência refletiria a perda específica de um armazenamento articulador temporário que constitui um dos componentes do sistema de armazenamento de curto prazo.
A passagem do tempo é crítica, pois leva ao decaimento da retenção da informação. É um processo que depende intensamente de atenção.
A memória de longo prazo representa a capacidade de armazenar grande quantidade de informações por período indefinido de tempo; a atenção do indivíduo pode ser desviada da informação crítica sem prejuízo da memória. Acredita-se que as informações que são repetidas na memória de curto prazo poderiam resultar em memórias de longo prazo, num processo denominado consolidação da memória. Todavia, alguns pacientes com distúrbios da memória de curto prazo são capazes de formar memórias de longo prazo, sugerindo uma independência entre esses sistemas.
Há considerável controvérsia sobre os limites temporais entre esses tipos de memória: o que é considerado como definição temporal de memória de curto prazo por alguns autores é, para outros, o critério para memória de longo prazo. Este quadro levou pesquisadores a considerar essa distinção artificial e a propor a existência de níveis sucessivos (contínuos) de codificação da informação (e.g., Craik & Lockhart, 1972); todavia, as observações com pacientes amnésicos, como H.M., que têm uma queda abrupta na memória quando sua atenção é desviada do material a ser memorizado, constituem argumentos a favor da distinção (e.g., Squire, 1987).
2. Memória recente e remota
Esta distinção aplica-se à dimensão temporal da memória. Em humanos, questões usualmente utilizadas por pesquisadores para avaliar a memória recente são: Qual foi seu café da manhã hoje? Onde você estacionou seu carro antes de chegar aqui? Que programa de TV você assistiu ontem à noite? A avaliação da memória remota é realizada através de questões sobre a infância, ou através da capacidade do indivíduo de lembrar antigos programas de TV. Em animais, tarefas de discriminação condicional com retardo (ver abaixo) são utilizadas para avaliar a memória recente. O gradiente temporal de interferência sugere mecanismos de processamento da informação distintos. Também aqui não há consenso entre os pesquisadores quanto à demarcação temporal entre os tipos de memória.
3. Memória operacional e de referência
Memória operacional, como inicialmente definida por Honig (1978) e Olton, Becker e Handelman (1979), refere-se a um tipo de memória que codifica o contexto temporal específico da informação, e que pode ser " apagada" depois de ter sido utilizada.
As tarefas normalmente utilizadas para testar memória operacional envolvem algum tipo de discriminação condicional com ou sem retardo.
Por exemplo, um estímulo crítico é apresentado e então removido (fase de informação). Depois de um intervalo de tempo, duas ou mais alternativas são apresentadas (fase de escolha). A opção pela resposta correta depende dos (ou é condicional aos) eventos críticos apresentados na fase de informação. Diversos pares de fases (uma de informação e uma de escolha) são apresentados ao longo do treinamento do animal, sendo que o estímulo apresentado na tentativa de informação varia. Assim, o evento apresentado na tentativa de informação é específico àquele par de tentativas, de modo que a única forma de identificar a resposta correta na tentativa de escolha é reter alguma representação dessa informação. Este procedimento é usualmente referido como delay-matching-to-sample -DMTS ou delay-nonmatching-to-sample - DNMTS (no caso do estímulo correto na tentativa de escolha ser igual ou diferente, respectivamente, em relação ao estímulo correto na tentativa de informação). Este procedimento está sujeito a interferência pró-ativa, isto é, como o estímulo correto difere nos vários pares de tentativas, pode haver competição entre traços de memória sobre qual é o estímulo correto (Grant, 1975). O fenômeno de interferência pró-ativa torna-se particularmente evidente quando o estímulo errado num dado par de tentativas corresponde ao estímulo correto do par de tentativas anterior. Macacos cometem uma quantidade de erros significativamente maior nestes casos (Worsham, 1975). Ademais, o treinamento dos animais com um pequeno número de estímulos resulta numa maior percentagem de erros em relação a situações em que se utiliza uma grande quantidade de estímulos; neste último caso o número de vezes em que cada estímulo é reapresentado como incorreto, tendo sido correto no par de tentativas anterior, é proporcionalmente menor, resultando numa menor interferência.
O labirinto radial de 8 braços (Olton & Samuelson, 1976) representa outra tarefa que torna necessário o uso da memória operacional. Ratos são colocados numa plataforma central octogonal da qual irradiam 8 braços, um de cada lado do octógono, sendo colocado alimento na extremidade de cada braço no início de cada sessão de treino. O desempenho ótimo da tarefa consiste em escolher braços sempre diferentes numa dada sessão, evitando revisitar um braço do qual o alimento já foi obtido. Depois de alguns dias de treino os ratos escolhem em média 7,6 braços diferentes nas primeiras 8 escolhas. Os autores concluem que os ratos mantêm uma " lista" atualizada dos braços já visitados naquela sessão na memória operacional. Diversas sessões de treino podem ser realizadas sucessivamente no mesmo dia, com pequenos intervalos de tempo entre elas; durante esses intervalos, os animais são retidos na plataforma central do labirinto e alimento é recolocado ao final de cada braço. A quantidade de erros cometidos neste esquema de treinamento é similar à de animais submetidos a apenas uma sessão de treino por dia, indicando que as informações sobre a sessão anterior contidas na memória operacional não interferem nas da sessão em curso. Essa ausência de interferência é interpretada por Olton como indicativa de que os animais " apagam" o conteúdo da memória operacional entre as sessões, de modo a não confundir a memória sobre a lista de braços visitados na sessão anterior com a da presente sessão. Supõe-se que os animais devam aprender algo acerca do prazo de utilidade das informações, sendo este o fator determinante para sua manutenção na memória operacional; quando deixam de ser úteis podem ser apagadas. O tempo de manutenção de uma informação na memória operacional depende portanto da relevância dessa informação; a simples passagem do tempo não determina inexoravelmente o decaimento da memória, como parece ocorrer no caso da memória recente.
A memória de referência independe do contexto específico da informação; nela, armazenam-se informações aplicáveis a diversas situações (no caso do labirinto radial de 8 braços, a todas as tentativas) (Honig, 1978). Assim, seriam informações "inativas" (ou latentes), até que sejam ativadas pela apresentação de estímulos apropriados, o que corresponde à evocação ou lembrança (Honig, 1978). Tarefas que envolvem discriminação simultânea são normalmente utilizadas para testar memória de referência. Por exemplo, dois ou mais estímulos são apresentados. A resposta a um dos estímulos é sempre correta, independentemente da tentativa; sendo constante o estímulo crítico, não há necessidade de atualizar a informação a cada tentativa, nem possibilidade de interferência.
A diferença crítica entre esses dois tipos de memória é que no caso da memória operacional estímulos diferentes podem governar o critério de resposta em diferentes tentativas, sendo pequena a quantidade de itens de informações passíveis de serem preservados, enquanto no caso da memória de referência o estímulo crítico é constante nas diferentes tentativas e praticamente ilimitada a quantidade de itens de informação passíveis de serem preservados.
4. Memória episódica e semântica
Memória episódica, como originalmente descrita, refere-se à memória autobiográfica de eventos específicos temporal e espacialmente localizados. Este sistema contém informações sobre o contexto em que um evento ocorreu (Tulving, 1972; Schacter & Tulving, 1982), sendo, portanto, similar à memória operacional. Todavia, por referir-se à evocação de eventos já ocorridos (ou episódios), difere da memória operacional que se refere a um evento em curso (Honig, 1978). Em pacientes amnésicos, o conhecimento episódico adquirido no início da vida usualmente está preservado (Squire & Cohen, 1984; Squire, 1987). Trata-se de um tipo de memória autobiográfica pelo qual o indivíduo lembra o evento ou informação, bem como quando e onde ele ocorreu.
A memória semântica independe do contexto e contém informações sobre relações lógicas entre os eventos do ambiente, como, por exemplo, conceitos gerais, linguagem, fatos e regras de funcionamento do mundo, muitos dos quais estão explicitamente acessíveis. A maior parte do conhecimento semântico é adquirida nas primeiras décadas de. vida, sendo particularmente resistente à desorganização, não estando usualmente prejudicada em pacientes amnésicos (Squire & Cohen, 1984; Squire, 1987). A memória semântica é geralmente entendida como a responsável pelo desempenho em tarefas que envolvem completar palavras, decisões léxicas e identificação de palavras, nas quais o sujeito faz uso de um conhecimento pré-existente sobre palavras e conceitos. Assemelha-se, nestes casos, à memória implícita, mas difere da memória de procedimento.
5. Memória de procedimento e memória declarativa
A memória de procedimento refere-se à informação sobre regras e procedimentos que são aplicáveis a uma variedade de circunstâncias diferentes (Cohen, 1984). Este sistema de memória não permitiria o acesso explícito ao conteúdo de conhecimento, e se expressaria apenas através do desempenho; isto é, através da ativação das estruturas de processamento ou procedimentos envolvidos nas tarefas de aprendizagem. A aquisição de habilidades motoras envolve esse tipo de aprendizagem (daí o nome "de procedimento"). Todavia, há aprendizagens não-motoras que envolvem um processamento similar de informações, como é o caso do desenvolvimento de habilidades perceptuais (e.g., leitura em espelho) e de habilidades cognitivas (e.g., desempenho na torre de Hanói). Por esta razão, o sistema seria melhor denominado como sendo não-declarativo. A aquisição e retenção deste tipo de informação decorreria da plasticidade inerente às estruturas de processamento que se modificam em cada ocasião em que a informação é processada, o que requer múltiplas tentativas de aprendizagem e permite pouca elaboração sobre o que foi aprendido.
Embora pacientes amnésicos possam adquirir essas habilidades e exibam pré-ativação, eles não lembram onde, quando e sob que condições as informações foram adquiridas.
A aprendizagem de tarefas repetitivas por animais, como a discriminação visual que envolveria associações estímulo-resposta automáticas ou hábitos, é tida como análoga a tarefas que envolvem aprendizagem de procedimento em humanos (e.g., Mishkin, Malamut & Bachevalier, 1984).
A memória declarativa refere-se, por sua vez, a um sistema de conhecimento em que a informação específica e factual é armazenada de uma forma explicitamente acessível para uso posterior, sendo evocável em função da demanda (Cohen, 1984). Este sistema adquiriria e manteria uma representação dos produtos específicos das operações realizadas pelas estruturas de processamento ativadas durante as tarefas de aprendizagem, envolvendo a criação de novas estruturas de dados para representar explicitamente os resultados das experiências. De acordo com esta descrição, o conteúdo da memória declarativa constitui o objeto da maioria das investigações sobre memória2
6. Memória implícita e explícita
Memória explícita refere-se à lembrança consciente de experiências prévias, usualmente avaliada nos testes de evocação livre e de reconhecimento, que requer evocação intencional da informação armazenada (Graf & Schacter, 1985; Schacter, 1987). É observada em tarefas típicas de laboratório quando uma pessoa é instruída para aprender uma lista de palavras. É um tipo de memória da qual a pessoa está consciente, tem a intenção de evocar a informação, e pode processá-la conscientemente.
Memória implícita refere-se à melhora no desempenho de dadas tarefas, sem referência explícita às experiências prévias que facultam essa melhora, ou seja, sem necessidade de que seja consciente. A existência de memória implícita é inferida através da facilitação do desempenho, uma vez que está ausente o acesso consciente ou intencional à informação. É grande a quantidade de tarefas nas quais a memória implícita é evidenciada (para uma revisão, ver Schacter, 1987). Por exemplo, (1) em tarefas envolvendo reaprendizagem de material previamente estudado, o desempenho pode estar facilitado sem que o sujeito possa referir-se explicitamente ao episódio de aprendizagem prévio (Slamecka, 1985), ou mesmo ao material estudado (esse material não é lembrado ou mesmo reconhecido pelos sujeitos) (Nelson, 1978); (2) em situações sobre efeitos de informações codificadas subliminarmente, mostra-se que estímulos não percebidos conscientemente (pela sua exposição por intervalos de tempo da ordem de 5 ms) influenciam o comportamento subseqüente; (3) em testes de condicionamento, sujeitos normais podem adquirir respostas classicamente condicionadas sem consciência das contingências de condicionamento; (4) em situações de pré-ativação, há uma facilitação no processamento de um dado estímulo em função de sua apresentação prévia, sem lembrança explícita da informação. Num teste típico de identificação de palavras, sujeitos são expostos por 30 ms a um estímulo e, então, tentam identificá-lo. A pré-ativação é evidenciada, por exemplo, pelo aumento da precisão na identificação de palavras previamente apresentadas em relação a palavras não apresentadas previamente, ou pelo decréscimo no tempo de exposição necessário à identificação de itens prévia e recentemente apresentados. A duração do efeito de pré-ativação pode persistir por dias ou semanas (no caso de testes para completar palavras) ou por minutos ou horas (no caso de decisões léxicas); em qualquer caso, não é suscetível às interferências pró-ativa e retroativa que se manifestam no caso da memória explícita. Por outro lado, em testes de associação e de lembrança por dicas, Schacter e McGlynn (1987) citados por Schacter (1987), mostraram que a memória implícita e explícita para associações adquiridas recentemente dependem do processamento elaborativo realizado durante o período de estudo, sugerindo que, ao menos em parte, memória implícita e explícita partilham características comuns.
A memória implícita cobre uma ampla gama de fenômenos. Alguns destes fenômenos podem ser agrupados em função da quantidade de repetições necessária para que a memória implícita se manifeste. Por exemplo, no caso da aquisição de habilidades motoras, perceptuais e cognitivas e de condicionamento, o número de tentativas necessárias para a estabilização da memória é grande. Por outro lado, em situações de retenção durante reaprendizagem, estímulos codificados subliminarmente e pré-ativação, o fenômeno pode manifestar-se depois de apenas uma ou duas exposições ao material do teste.
A similaridade entre memória implícita/explícita e memória de procedimento/declarativa, respectivamente, tem levado pesquisadores a considerar essas distinções como sinônimas (e.g., Delis, 1989). Todavia, essa colocação parece apenas parcialmente verdadeira, e depende da classe de fenômenos considerada. Por um lado, a memória de procedimento, como a memória implícita, inclui a aprendizagem de habilidades motoras, perceptuais e cognitivas, isto é, processos que dependem de treinamento repetitivo e pré-ativação (Cohen, 1984). Os fenômenos de retenção durante a reaprendizagem e a percepção subliminar não são contudo considerados como parte do sistema de procedimento: a memória de procedimento poderia ser então classificada como sub-item da memória implícita. Congruentemente, pacientes amnésicos exibem memória implícita em situações não categorizadas como procedimentais, como, por exemplo, a aquisição de conhecimentos sobre novos fatos e vocabulário, mesmo sem memória explícita da aquisição dessa informação (Glisky, Schacter & Tulving, 1986; Schacter, Harbluk & McLachlan, 1984). Por outro lado, torna-se cada vez mais claro que processos diferentes subjazem aos diversos fenômenos que permitem evidenciar a memória implícita. Por exemplo, a aprendizagem de habilidades depende de treinamento repetitivo, diferindo, assim, da pré-ativação; ademais, essas duas classes de fenômenos podem ser dissociadas experimentalmente (Butters, 1987). Além disso, a retenção durante a reaprendizagem depende de aquisição intencional da informação, i.e., da atenção devotada ao processamento da informação no momento em que ela é apresentada, ao passo que a pré-ativação e a percepção subliminar independem da aquisição intencional. Além da necessidade de repetição, nos casos de memória de procedimento parece haver necessidade de atenção, pelo menos durante a fase inicial de aquisição, finda a qual passa a independer da atenção. Essa posterior independência da memória de procedimento em relação a processos de atenção parece estar associada ao fato de que sua aquisição depende de repetição da mesma tarefa, o que permite automatização. Nesse sentido, seu desempenho depende apenas da ativação das estruturas de processamento nela envolvidas, sem o envolvimento de intencionalidade em todas as etapas da execução.
Em conclusão, algumas dessas distinções certamente referem-se a processos comuns. Memória explícita e memória declarativa referem-se às mesmas classes de fenômenos. Por outro lado, memória implícita refere-se a classes diversas de fenômenos (Schacter, 1987), algumas das quais correspondem à memória de procedimento (Cohen, 1984) e outras não.
A memória episódica e parte do processamento caracterizado como memória semântica e memória de referência podem ser caracterizados como conhecimento declarativo (ou explícito). Todavia, parte do conhecimento semântico e de referência pode ser considerado como similar a fenômenos de memória implícita. Dentre as distinções que enfatizam o curso temporal do processamento de informações, a memória de longo prazo e a memória remota assemelham-se entre si, e, dependendo da natureza do processamento, podem ainda ser caracterizadas como declarativas (ou explícitas), ou implícitas; por outro lado, memória operacional difere da memória de curto prazo, já que nesta última o processamento da informação decai com a passagem do tempo e requer " repetição mental", enquanto a primeira depende de um contexto temporal específico, sendo mantida sob processamento em função de sua relevância.
Esse quadro, apresentado de forma resumida, enfatiza a necessidade de uma classificação que permita, por um lado, evitar sobreposições de conceitos e, por outro, estabelecer relações entre esses tipos postulados de processos.
C. A classificação de Squire e Zola-Morgan (1991)
Squire e Zola-Morgan (1991) e Squire (1992) propuseram uma classificação elegante dos sistemas de memória, baseados no critério de acesso consciente ao conteúdo da informação. A memória é subdividida em declarativa (ou explícita), no caso de haver acesso consciente ao conteúdo da informação, e não-declarativa (ou implícita), nos casos em que o conteúdo da informação não está acessível à consciência, sendo apenas evidenciável através do desempenho. A memória declarativa é subdividida em memória para fatos e para eventos, e a memória não-declarativa em habilidades e hábitos, pré-ativação, condicionamento clássico simples e aprendizagem não-associativa, incluídas aqui habituação e sensibilização (ver Figura 1).
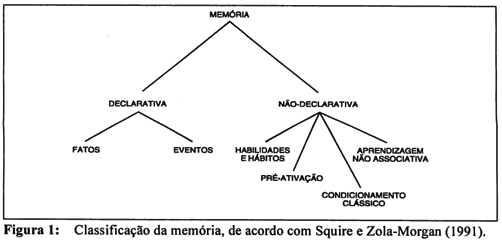
Observe-se que embora memória não-declarativa e memória implícita tenham sido tratadas como sinônimos, a classe de fenômenos reunidos sob essa denominação na classificação de Squire e Zola-Morgan (1991) não corresponde integralmente aos fenômenos caracterizados como memória implícita por Schacter (1987).
O sistema declarativo seria apto a manifestar-se através de múltiplos sistemas de resposta, enquanto o sistema não-declarativo seria inflexível, isto é, apresentaria acesso limitado a sistemas não envolvidos na aprendizagem original. Além disso, a aquisição de informações não-declarativas é referida como dependente de mudanças cumulativas nos sistemas perceptuo-motores em cada ocasião em que esses sistemas são acionados.
Do ponto de vista neural, o sistema declarativo é tratado como dependente da integridade do lobo temporal medial, enquanto o sistema não-declarativo seria independente das estruturas que compõem o lobo temporal (Squire & Zola-Morgan, 1991). Essa dependência do sistema declarativo em relação ao lobo temporal medial é crucial durante o estabelecimento de memórias declarativas de longo-prazo. O armazenamento destas dar-se-ia no neocórtex, sob prolongado controle das estruturas do lobo temporal medial; findo esse processo, no entanto, essas memórias tornam-se independentes das estruturas do lobo temporal.
O modelo incorpora algumas das principais características dos sistemas de memória descritos, mas exclui outras (memória de curto prazo e memória operacional). Além disso, há questões controversas que merecem mais discussão. (1) Há necessidade de repetição para a aquisição do conhecimento não-declarativo? (2) Há necessidade de repetição para a aquisição de conhecimento declarativo factual? (3) A aquisição de conhecimento não-declarativo depende de controle pelo sistema declarativo? (4) Como deve ser classificada a capacidade presente nos pacientes portadores de lesões no lobo temporal medial para processar informações declarativas por curtos intervalos de tempo, em contraposição à sua incapacidade para formar um registro permanente para esse tipo de informação? (5) O limite entre os sistemas declarativo e não-declarativo pode ser estabelecido precisamente? (4) Como interpretar dados de pacientes com lesões fora do lobo temporal medial que também exibem prejuízos no processamento de informações de natureza declarativa e integridade do processamento não-declarativo?
As discussões que se seguem tratarão dos aspectos controversos do modelo de Squire e Zola-Morgan (1991) e, paralelamente, permitirão lançar as bases de um modelo alternativo que incorpora algumas das proposições apresentadas por esses autores.
1. Aquisição do conhecimento não-declarativo
Se a aquisição de informações pelo sistema não-declarativo depende de mudanças cumulativas que ocorrem em cada ocasião que o sistema é acionado, como proposto por Squire e Zola-Morgan (1991), isto implica que o sistema possui algumas características adicionais, como por exemplo, (1) a necessidade de treino repetitivo para a aquisição do comportamento, (2) o caráter gradual da aprendizagem, (3) a detecção e preservação de elementos invariantes nas situações comportamentais, (4) a pouca flexibilidade na utilização deste sistema para a solução de problemas com elementos invariantes diferentes, e (5) a independência em relação à atenção. Estas características, no entanto, não se aplicam a todas as classes de fenômenos consideradas como parte do sistema não-declarativo. A pré-ativação, por exemplo, não é considerada como um fenômeno que requer treino repetitivo e depende de aquisição gradual. Por outro lado, a aquisição de habilidades e hábitos, e o condicionamento clássico, parecem depender de atenção e do controle declarativo (inclusive nos pacientes amnésicos temporais), pelo menos nas fases iniciais da aquisição.
Essas fontes de controvérsia parecem decorrer do fato de, no tratamento geralmente dado aos fenômenos não-declarativos, não se considerar o processo de aquisição de informações ao longo da história individual, nem a interferência desse processo, em conjunto com o sistema declarativo, na aquisição de novos conhecimentos não-declarativos.
A tese a ser defendida aqui é que a organização do processamento de informações, incluindo a escolha e execução de ações apropriadas, depende em parte do sistema declarativo e/ou da memória operacional: a organização é originalmente consciente, intencional, com certo grau de monitoramento consciente. Todavia, uma vez que uma tarefa foi aprendida, pode-se prescindir da consciência que ainda é necessária, assim mesmo, em vários momentos. Quando as tarefas são inicialmente adquiridas, sua organização precisa ser consciente; uma vez automatizadas as habilidades, regras e estratégias, o sistema passa a prescindir do funcionamento do sistema declarativo e/ou operacional; a intenção de desempenhar uma tarefa não-habitual ou não-dirigida por estímulos mobiliza novamente a consciência.
Nas fases iniciais de desenvolvimento de um indivíduo, a aquisição de uma habilidade motora envolve atentar para a seqüência precisa de movimentos a ser seguida, e o controle consciente desses movimentos, bem como a avaliação também consciente de suas conseqüências, através do sistema declarativo e/ou operacional. À medida em que essa habilidade torna-se, através da prática, automatizada, sua dependência em relação ao comando explícito diminui.
Sub-rotinas distintas, adquiridas de forma similar, controlam diversos movimentos " básicos". Movimentos complexos podem ser produzidos através da ação conjunta de diversas dessas sub-rotinas. Novamente, nas fases iniciais da aquisição, agora de um movimento complexo, haverá necessidade de controle pelo sistema explícito. Também através da execução repetitiva desse movimento, haverá a criação de uma nova sub-rotina. Esta, no entanto, pode valer-se das sub-rotinas pré-existentes e responsáveis pelos subcomponentes do movimento complexo. Assim, este segundo tipo de sub-rotina teria a função de acionar, de forma orquestrada, sub-rotinas pré-existentes. Nesse sentido, podem ser vistas como ocupando um nível hierárquico diferente. A criação de uma sub-rotina para trocar a marcha de um automóvel (sub-rotina de nível "n") constitui um bom exemplo para esse processo. Os movimentos de mãos, braços, pernas e tronco necessários para o ato de trocar a marcha são controlados por sub-rotinas já estabelecidas anteriormente (digamos, sub-rotinas de nível "n-1") capazes de acionar outras sub-rotinas previamente adquiridas (sub-rotinas de nível "n-2"). O processo de aprendizagem corresponde, neste caso, a " criar", através do treinamento repetitivo, a sub-rotina de nível "n" capaz de acionar, de forma orquestrada, cada uma das sub-rotinas de nível "n-1" previamente adquiridas. Esse processo de criação da nova sub-rotina é inicialmente controlado passo-a-passo pelo sistema explícito e/ou operacional cuja atividade representa objetivos e planos gerados por instruções verbais ou observação e imitação; a atenção é necessária. À medida que a habilidade é adquirida, ela passa a prescindir do controle imediato daqueles sistemas. Então, a simples atividade, nesses sistemas, correspondente a objetivos e planos, desencadeia, automaticamente, a completa execução da habilidade correspondente.
A aquisição de habilidades perceptuais e cognitivas dar-se-ia de forma similar.
Por serem processos que se tornaram automáticos, pouca ou nenhuma atenção é necessária. Sabe-se que a capacidade de prestar atenção, por ser limitada, constitui a principal limitação do processamento de informação. Assim, um processamento automático por consumir pouca ou nenhuma atenção torna-se adaptativo quando intensamente utilizado: pode ser realizado ao mesmo tempo em que a atenção é direcionada para outras informações . Embora o processamento automático traga grandes vantagens adaptativas, sua principal desvantagem é que o indivíduo tem pouco controle explícito sobre as informações processadas.
Esse conjunto de alterações cumulativas no sistema de processamento não-declarativo será, posteriormente, responsável por uma variedade de fenômenos descritos sob o título de memória implícita (ver Schacter, 1987), incluída aqui a pré-ativação. É interessante notar, neste contexto, que pacientes severamente amnésicos mostram pré-ativação normal para itens com representações pré-existentes na memória (e.g., palavras familiares e expressões idiomáticas), mas não exibem o fenômeno de pré-ativação para não-palavras ou associações entre pares de palavras não-relacionadas (Schacter, 1985; Schacter & Graf, 1986).
As alterações cumulativas do sistema não-declarativo não decorrem apenas da estimulação repetitiva por elementos concretos, mas também de categorias, regras e padrões invariantes presentes nos eventos repetitivos. Isto permite explicar, por exemplo, porque a apresentação prévia de uma palavra com conotações negativas pode induzir o indivíduo a interpretar uma segunda palavra, em princípio neutra, de forma negativa, sem que perceba conscientemente essa interferência (Chaves, comunicação pessoal). Na ausência da primeira palavra, tanto os aspectos positivos como os negativos associados à palavra neutra seriam ativados; entretanto, a prévia ativação de aspectos negativos pelo "pré-ativador" (primeira palavra) ativa os atributos negativos para a palavra neutra.
Informações relevantes por intervalos definidos de tempo são processadas de forma " explicitável" principalmente durante o intervalo relevante, findo o qual passa a haver decaimento no processamento. Neste caso, embora o conteúdo da informação se relacione com o sistema explícito, é o sistema implícito que determina o intervalo de tempo durante o qual ela deve ser processada, o que depende da experiência prévia repetitiva em situações similares. Um bom exemplo desse fenômeno é o processamento da informação realizado por motoristas sobre faróis em cruzamentos. Esta informação é relevante apenas por curtos períodos de tempo, até que se atravesse o cruzamento. São comuns relatos de motoristas que têm dúvidas sobre se o farol estava aberto ou fechado no cruzamento atravessado segundos antes; certamente essa informação foi processada adequadamente, enquanto foi útil; todavia, prolongar sua retenção no sistema poderia sobrecarregá-lo desnecessariamente. Assim, o sistema não-declarativo, em função da automatização ocorrida previamente, estabelece que o tempo de processamento da informação deve ser curto.
Provavelmente, a aquisição de conhecimento declarativo para fatos (memória semântica, na descrição de Tulving, 1972) também depende de estimulação repetitiva, de modo a que o sistema nervoso seja exposto aos aspectos invariantes da informação. Por exemplo, a formação do conceito de " árvore" depende de o indivíduo, em diversas ocasiões, ser exposto ao estímulo juntamente com o signo. Apesar de as árvores serem diferentes entre si, elas possuem características comuns que definirão o conceito. As informações sobre os estímulos apresentados nas diferentes exposições às árvores devem ser retidas no sistema nervoso para posterior comparação e identificação dos aspectos invariantes dos mesmos, de forma declarativa.
Assim, uma grande quantidade de processamentos ocorre fora do domínio declarativo desde que habilidades, regras e estratégias necessárias a essas atividades tenham sido automatizadas.
2. Memória declarativa de curto prazo em pacientes temporais
Uma das descrições mais impressionantes sobre a preservação de memória declarativa de curto prazo em pacientes portadores de lesões temporais, foi realizada por Karl Pribram (1986) sobre o paradigmático paciente H.M.
Pribram (1986) escreveu:
Lembro-me claramente de minha primeira entrevista com H.M.. Bill Scoville pediu-me para avaliar H.M. pois ele parecia peculiar. Eu estava ávido para examiná-lo visando investigar como suas respostas emocionais diferiam das de indivíduos não operados: tínhamos acabado de obter resultados em experimentos de esquiva condicionada (...) que correspondiam à primeira descrição de deficiência experimental quantitativa após lesão do hipocampo. Para minha surpresa, portanto, H.M. respondeu normalmente em todos os aspectos. Ele discutiu sua relação com sua família; tivemos uma interessante conversa sobre um assunto de interesse comum, envolvendo uma possível viagem da Cidade do Cabo para o Cairo, durante a qual H.M. ficou bastante animado. Testei sua lembrança para listas, como números de telefones, e novamente constatei seu desempenho normal. Neste momento da entrevista, fui chamado ao telefone. Quando retornei, tentei, sem sucesso, lembrar onde tínhamos parado e então pedi ajuda para H.M.: falávamos sobre números de telefones? Ou fazíamos subtrações ? H.M. fitou-me por algum tempo e perguntou com uma expressão confusa: 'Doutor, já o vi antes?'
A distração causada pela interrupção, aparentemente tinha produzido uma amnésia completa. H.M. tinha me parecido tão completamente normal que cheguei a duvidar que Scoville tivesse tido sucesso na cirurgia. Eu não tinha observado qualquer mudança em suas emoções ou memória (a viagem pela África era um interesse pré-operatório). Agora a deficiência de memória era marcante (p.332-3).
A memória de curto prazo, bem como a capacidade para evocar e utilizar informações declarativas adquiridas pré-cirurgicamente, estava normal. Isto enfatiza a independência da memória de curto prazo em relação ao sistema declarativo de longo prazo, e dissocia a evocação de informações do sistema declarativo das estruturas temporais. Além disso, a conversação coerente evidencia que a memória operacional não estava afetada, exceto quando do desvio da atenção. Depreende-se, portanto, que as estruturas que compõem o lobo temporal medial são relevantes para o armazenamento, mas não para a evocação de informações de natureza declarativa e para a manutenção de objetivos ativos, bem como para a manutenção de memórias de curto prazo.
3. Limites entre os sistema declarativo e não-declarativo
Não está claro se uma memória implícita pode, em determinadas condições, ser lembrada explicitamente.
Depreende-se das colocações de Squire e Zola-Morgan (1991) que a memória declarativa e não-declarativa atuam de forma totalmente independente. O código de processamento utilizado pelo sistema não-declarativo seria incompatível com o do sistema declarativo, impossibilitando a interação entre sistemas. Posição semelhante foi defendida por Cohen (1984) para a distinção memória declarativa/memória de procedimento.
Um exemplo elegante que apóia essa interpretação advém dos estudos de Winnick e Daniel (1970). Esses autores realizaram testes de identificação de palavras em três condições diferentes: (1) leitura de palavras familiares a partir da apresentação visual das mesmas, (2) verbalização de palavras a partir da apresentação de figuras correspondentes, e (3) verbalização de palavras a partir da suas definições . Houve um significativo efeito de pré-ativação na condição (1) e ausência de pré-ativação nas condições (2) e (3). Entretanto, em testes de lembrança intencional, as palavras utilizadas nas condições (2) e (3) foram mais lembradas do que aquelas utilizadas na condição (1).
Num outro estudo, realizado por Jacoby e Dallas (1981), sujeitos foram expostos a listas de palavras familiares em duas condições de processamento diferentes; na primeira, tinham de refletir sobre o significado das palavras apresentadas e, na segunda, tinham de dizer se uma determinada letra constava da palavra. Num teste de reconhecimento realizado posteriormente, os autores verificaram que a memória declarativa para a lista de palavras da primeira condição era melhor em relação à segunda condição. A memória não-declarativa para as listas de palavras apresentadas nas duas condições de estudo era contudo similar, como evidenciado por um teste posterior de identificação de palavras. Nesse mesmo estudo, os autores mostraram que uma mudança de modalidade de auditiva (na fase de exposição ao material) para visual (na fase de teste com o material) interferiu nos efeitos de pré-ativação (memória não-declarativa), tendo pouca ou nenhuma interferência em testes de reconhecimento (memória declarativa).
A observação de que a memória implícita não é afetada pelas variáveis experimentais que geralmente surtem efeitos marcados na memória explícita e que o desempenho nos testes de memória implícita independe do desempenho nos testes de memória explícita é consistente com o modelo de que esses sistemas de memória exibem funcionamento independente.
Por outro lado, diversas linhas de evidência mostram que, pelo menos em determinadas condições, esses sistemas partilham de características e elementos comuns.
Na amnésia pós-hipnótica sugestionada pode-se induzir o indivíduo a lembrar-se de informações obtidas durante a sessão de hipnose pela simples apresentação de um sinal arbitrário definido também durante a sessão. Isso mostra que a amnésia decorre de dificuldades na evocação e não de uma falha de aquisição ou armazenamento da informação. A evocação, no entanto, pode ser mais evidente dentro de certas modalidades. Isto é, indivíduos amnésicos para listas de palavras memorizadas durante uma sessão de hipnose terão uma forte tendência, posteriormente, a utilizar as informações sobre estas listas para associações entre palavras, categorização e tarefas de completar palavras (Kihlstrom, 1980). Assim, parece ter havido uma codificação adequada da informação que simplesmente não está acessível ao sistema declarativo; todavia, interfere no desempenho em tarefas típicas de memória não-declarativa.
Embora haja uma semelhança entre sugestão pós-hipnótica e memória implícita, pois ambas podem afetar o desempenho numa tarefa sem que o indivíduo tenha acesso consciente ao porquê, há algumas diferenças óbvias: sugestões pós-hipnóticas não correspondem a associações do tipo estímulo-resposta ou estímulo-estímulo, e não tiveram a oportunidade de tornarem-se automatizadas através da prática. No entanto, a alteração de desempenho por seu intermédio depende de atenção. Por exemplo, voluntários tinham que localizar dois dígitos diferentes simultaneamente, em listas de dígitos que apareciam numa tela de computador. A tarefa de localização de um dos dígitos foi apresentada aos voluntários durante uma sessão de hipnose com sugestão de amnésia, enquanto a tarefa de localização do segundo dígito foi apresentada sem hipnose e, portanto, sem amnésia sugerida. Observou-se que mesmo em situações em que as duas tarefas não eram conflitantes houve um comprometimento do desempenho, isto é, uma tarefa interferiu no desempenho da outra tarefa. Esses dados indicam que mesmo fora do domínio consciente (declarativo), houve necessidade de atenção para a execução da tarefa.
Assim, se por um lado existem situações em que o processamento implícito é claramente dissociável em relação ao sistema explícito, existem situações caracterizáveis como envolvendo conhecimento implícito nas quais o conhecimento parece estar acessível à consciência fenomenológica, podendo ser obtido diretamente através da introspecção; o acesso a este tipo de conhecimento parece ser matéria de ativação, de modo que o mesmo poderia ser caracterizado como " pré-declarativo". Se esse tipo de conhecimento é ativado acima de um certo limiar, torna-se consciente; se não, é implícito, pois interfere no desempenho sem que o indivíduo esteja ciente do mesmo.
O fenômeno da hipermnésia, em que a evocação de informações pode ser aumentada ao longo do tempo em decorrência de um esforço consciente de evocação (Haber & Erdelyi, 1967), permite especificar mais o funcionamento desse sistema. Erdelyi e Kleinbard (1978) expuseram um indivíduo a uma lista de 40 figuras simples, por 5 segundos cada.
Em seguida, o sujeito tinha que denominá-las, por lembrança. Inicialmente, cerca de 18 figuras foram lembradas. Então, o sujeito foi estimulado a tentar "chutar" respostas, até completar 40 itens. Imediatamente após essa primeira tentativa de evocação, pediu-se que o indivíduo tentasse evocá-los novamente. Esse procedimento foi repetido várias vezes e pediu-se, então, que ele continuasse o processo em casa, por uma semana. Cerca de 70 horas depois da exposição às figuras, o sujeito foi capaz de lembrar-se de 33 itens.
Esses resultados contrastam com o decréscimo de retenção em função da passagem do tempo, descrito por Ebbinghaus (1964).
Deve-se enfatizar que há uma diferença fundamental entre a apresentação de uma certa quantidade de itens e sua posterior evocação em função da passagem do tempo (o que geralmente produz curvas de decréscimo da quantidade de itens evocados, como descrito por Ebbinghaus, 1964) e a posterior evocação pelo esforço consciente (Ederlyi & Kleinbard, 1978), que resulta num aumento do número de itens evocados com a passagem do tempo. A diferença crítica parece residir no esforço consciente de evocar os itens no caso da hipermnésia, em contraposição à retirada intencional da consciência dos estímulos, na situação de evocação simples (como se o indivíduo esperasse evocar os itens mais prontamente pelo relaxamento consciente). Segundo Erdelyi (1988), o desdobramento voluntário da consciência em direção ao mesmo complexo de memória de forma crônica e de longo prazo determina a acessibilidade da memória à experiência consciente.
Parece haver duas funções de memória, uma de melhora e outra de decaimento, que dependem do tipo de tratamento consciente que recebem. Uma vez que o esforço consciente de evocação pode alterar seletivamente, e a posteriori, a extensão com que essas funções de melhora e decaimento operam para um determinado complexo de memória, segue-se que podemos intencionalmente determinar o acesso à memória de eventos passados o que, por sua vez, depende de motivações, medos, desejos, etc, constituindo um mecanismo cognitivo de controle sobre o que permanece passível de evocação.
Mesmo quando não imediatamente acessíveis, as informações permanecem no sistema nervoso por certo período de tempo, e podem ser evocadas mediante esforço consciente. Além disso, informações com grande conteúdo emocional negativo podem ser mantidas de forma mais imediatamente acessível, enquanto informações com conteúdo positivo estariam menos acessíveis. Mais que isso, o contexto de apresentação de uma dada palavra poderia interferir em sua " evocabilidade"; por exemplo, a palavra "dinheiro" precedida da palavra "assalto" ou "férias", poderia, implicitamente, assumir significado negativo e positivo, respectivamente, resultando numa maior ou menor evocação. Diferentemente do conhecimento implícito automatizado, este tipo de conhecimento pré-declarativo estaria acessível à consciência.
4. Ausência de processamento declarativo em pacientes com o lobo temporal íntegro.
Diversas fontes de evidências indicam que há prejuízo de processamento declarativo em indivíduos com o lobo temporal íntegro. Mais que isso, a capacidade para processar declarativamente uma dada informação parece estar relacionada com o tipo de resposta requerida do paciente, ou mesmo com sua capacidade para emitir uma determinada resposta.
Pacientes com lesões nas áreas de projeção visual primária, mais especificamente, com danos neurológicos nas projeções da retina para o córtex occipital esquerdo, que não possuem experiência perceptual consciente dentro do campo afetado (blindsight ou "visão às cegas"), desempenham acima dos níveis do acaso em testes de discriminação forçada relativos a localização, orientação e outras dimensões do estímulo visual. Por exemplo, quando forçados a pegarem um objeto localizado nesse " campo cego", os ajustamentos preparatórios do pulso, dedos e braço são adequados à forma, orientação, tamanho, localização e distância do objeto, apesar da insistência do paciente em afirmar que não existe qualquer objeto naquela região do espaço. Assim, o principal problema deste paciente parece ser a experiência fenomenológica (consciente) de qualidades que foram adequadamente analisadas. Além disso, variações na intensidade do estímulo aparentemente interferem no desempenho dos indivíduos, o que sugere que o limiar para sua detecção consciente pode estar alterado (Weiskrantz, 1988).
Bisiach (1988) mostrou que, num paciente com lesões cerebrais no hemisfério direito, a detecção de estímulos no campo visual contralateral ao hemisfério lesado varia marcadamente em função da resposta requerida (verbal ou pressão de uma tecla). Numa situação em que LEDs (diodos emissores de luz) acendiam-se no lado direito ou no lado esquerdo do ponto de fixação visual, pacientes deveriam (1) relatar verbalmente a quantidade de LEDs que acenderam (um, dois ou nenhum) ou (2) pressionar uma tecla quando um LED acendeu, evitando pressioná-la no caso de dois ou nenhum LEDs terem acendido. Em ambos os casos, o paciente exibiu prejuízo de desempenho em relação ao hemicampo visual contralateral à lesão cerebral: a detecção de estímulos visuais foi normal no hemicampo visual direito e prejudicada no hemicampo visual esquerdo; i.e., em muitas tentativas com um único estímulo no lado esquerdo o paciente respondeu "nenhum" na condição verbal e não pressionou a tecla na condição motora. Além disso, em situações de estimulação bilateral, os pacientes comportaram-se como se apenas um estímulo tivesse sido apresentado (o do lado direito). Entretanto, a detecção do estímulo apresentado no hemicampo visual esquerdo foi significantemente inferior quando a resposta a ser emitida envolvia a pressão à tecla, tanto nos casos de estimulação simples como nos de estimulação dupla.
Em outro grupo de estudos, envolvendo pacientes com lesões frontais direitas que negligenciavam o hemiespaço esquerdo, uma luz verde ou vermelha era apresentada por 200 ms à direita ou à esquerda do ponto de fixação visual. Duas teclas de resposta (uma verde e a outra vermelha) foram colocadas uma em cada lado do espaço. A disposição das lâmpadas e teclas foi tal que gerou as 4 combinações possíveis (duas cruzadas e duas não-cruzadas). O paciente tinha que pressionar a tecla de mesma cor em relação ao estímulo apresentado. Nesta situação um paciente identificado como FS foi capaz de relatar verbalmente a cor da lâmpada apresentada no seu hemicampo visual direito em todas as tentativas de uma sessão sem tentativas com teclas. Ademais, exibiu respostas precisas a todas as tentativas com o estímulo no lado direito quando uma resposta do lado direito era requerida. Entretanto, a apresentação do estímulo ainda do lado direito, mas com a tecla para identificá-lo localizada agora no lado esquerdo, levava à ausência de qualquer reação em metade das tentativas; além disso, em algumas dessas ocasiões o paciente declarou espontaneamente que nenhum estímulo havia aparecido (Bisiach, Berti & Vallar, 1985). Ou seja, uma falha no sistema de ação relevante parece prevenir o processamento adicional daquela representação que teria sido (no caso da integridade do sistema de ação) a base para um relato ou resposta sobre ela.
Parece que o processamento consciente depende do direcionamento da atenção, que a intencionalidade é fator fundamental para o desempenho de uma ação consciente complexa e que o ambiente para o qual se desenvolve a ação faz parte da intencionalidade. Todavia, experimentos com pacientes portadores de secção do corpo caloso mostram a capacidade de desempenhar ações altamente direcionadas, sem percepção consciente.
JW, um paciente com secção completa do corpo caloso, cujo hemisfério direito é capaz de entender apenas palavras individuais (Gazzaniga, Smylie, Baynes, Hirst & McCleary, 1984) foi treinado a dizer que número, " 1" ou "2", havia sido apresentado por 150 ms a 6º de seu ponto de fixação (Gazzaniga, Holtzman & Smylie, 1987). JW era capaz de dizer precisamente que número havia aparecido independentemente do hemicampo visual em que o estímulo havia sido apresentado. Num segundo experimento, o número "2" foi substituído pelo número "9", apenas quando apresentado para o hemicampo visual esquerdo; ao hemicampo visual direito o número "2" continuou a ser apresentado. Isto é, " 1" ou "2" foram apresentados no hemicampo visual direito, enquanto " 1" ou "9" no hemicampo visual esquerdo. Considerando o viés previamente estabelecido, do conhecimento do hemisfério esquerdo de que a resposta envolvia " 1" ou " 2", JW respondeu à apresentação do número " 9" como se fosse " 2" de forma altamente consistente. Esses dados indicam que JW é capaz de discriminar números (" 1" de "diferente de "1") com o hemisfério direito e mesmo transferir o resultado dessa discriminação para o hemisfério esquerdo para que o mesmo expresse a discriminação verbalmente, em função de seu viés prévio. Todavia, num experimento do tipo DMTS em que se apresenta o estímulo " 1" ou "2" no hemicampo visual esquerdo e, posteriormente, um segundo estímulo " 1" ou "2" no hemicampo visual direito, o paciente é incapaz de comparar as informações apresentadas e dizer se são iguais ou diferentes. Ou seja, JW é incapaz de fazer comparações de informações " entre campos".
Estes resultados mostram que o hemisfério esquerdo do JW não foi capaz de apreender conscientemente uma informação presente no hemisfério direito e disponível para o sistema de resposta verbal do hemisfério esquerdo. Parece que a informação transmitida para o hemisfério esquerdo gera uma prontidão de resposta para uma de duas respostas possíveis. Essa prontidão é estabelecida fora do escopo da consciência.
Não está claro como a informação do hemisfério direito é transferida para o hemisfério esquerdo. Acredita-se que isso ocorra da mesma forma como estados emocionais podem ser transferidos entre os dois hemisférios desconectados (i.e., subcorticalmente).
Em estudos subseqüentes de apresentação de palavras em ambos os campo visuais verificou-se que quando os estímulos foram apresentados no campo visual direito a precisão e confiança na resposta foram altos. Quando os estímulos foram apresentados no campo visual esquerdo, a precisão da resposta foi alta, mas a confiança na resposta foi baixa (Gazzaniga, 1988).
Além disso, JW é capaz de "categorizar" palavras (Gazzaniga, 1988). Por exemplo, se a palavra " fruta" é apresentada ao seu hemisfério direito, a mão esquerda aponta para a palavra " maçã" dentre uma lista de palavras. Todavia, o hemisfério direito parece não ter idéia do que está ocorrendo. Parece que a capacidade de relacionar palavras não significa que o sistema de resposta saiba porque ele o está fazendo (Gazzaniga et al., 1984). Mais que isso, se projetar a figura de um cavalo no hemisfério direito, o hemisfério esquerdo prontamente dirá que nada viu. Se o experimentador disser algo do tipo " - não desenhe o que é, mas o que se coloca em cima dele", poderá ouvir a resposta "- do que você está falando? Eu não vi coisa alguma !!" Se um lápis estiver disponível para a mão esquerda, ela desenhará uma sela. JW disse não saber o que havia desenhado. Pediu-se então para que ele desenhasse o que foi projetado. A mão esquerda desenhou um cavalo e, após ter completado a figura, disse sobre seu primeiro desenho: "- aquilo deve ser uma sela".
A maioria desses estudos pressupõe que cada hemisfério cerebral constitui uma unidade funcional, um sistema independente que opera de forma similar, esteja conectado ou não com o outro lado. Todavia, isto não é assim. Por exemplo, o subteste dos "cubos" do WAIS (Wechsler Adult Intelligence Scale) requer que a pessoa organize de 4 a 9 cubos cujas faces são pintadas de branco, vermelho, e branco e vermelho, de modo que o padrão visual resultante se iguale ao de uma figura apresentada. Antes da secção do corpo caloso, pacientes desempenhavam essa tarefa tão bem quanto indivíduos normais, quando usavam a mão direita. Após a secção do corpo caloso, apareceram grandes dificuldades no desempenho da tarefa, tanto com a mão direita como com a esquerda. O tempo utilizado para resolver os padrões mais simples duplicou e a execução dos padrões mais complexos tornou-se impossível (Gazzaniga, 1988). Assim, como os resultados pós-operatórios, tanto para o hemisfério direito como para o esquerdo, estavam baixos, pode-se dizer que de alguma forma o hemisfério esquerdo beneficiava-se dos processos localizados no hemisfério direito aos quais ele tinha acesso através do corpo caloso.
Parece, portanto, haver fatores dissociáveis em atividades aparentemente unificadas, como se controladores executivos manipulassem dados de sistemas de processamento especializado. Esses controladores parecem localizar-se no hemisfério esquerdo, e, quando o hemisfério direito torna-se isolado de sua influência, as funções específicas do hemisfério direito tornam-se de difícil teste.
Os pacientes com secção do corpo caloso categorizam suas respostas controladas pelo hemisfério direito como emanadas de sua vontade e, como resultado, incorporam estes comportamentos em teorias que expliquem porque eles se comportam dessa forma. Por exemplo, JW teve a palavra "cor-de-rosa" projetada no hemisfério direito e a palavra "garrafa" no hemisfério esquerdo. Quando uma figura com 10 garrafas de formas e cores diferentes foi apresentada, ele imediatamente apontou, com a mão direita, para a garrafa cor-de-rosa. Quando perguntado porque havia feito isso ele respondeu: " - cor-de-rosa é uma cor legal". Outro exemplo: se a palavra "rir" for apresentada para o hemisfério direito, o paciente ri e, se perguntado porque o fez, responde: " - Vocês me vêm com esse monte de testes todo o mês. Que jeito estranho de viver(...)" (Gazzaniga, 1988, p.234). Isto é, embora capaz de ler e de fazer o indivíduo comportar-se de acordo com o significado da palavra lida, o hemisfério direito é incapaz de fazer o indivíduo expressar-se verbal e conscientemente sobre o que foi lido.
Resultados semelhantes aparecem em situações de pré-ativação ou nos pacientes amnésicos que são incapazes de lembrar de onde partiu a idéia de fazer algo ou porque são melhores na execução de alguma tarefa, e também nas explicações oferecidas por sujeitos submetidos a sugestão hipnótica para explicar a origem de seus comportamentos.
D. Uma classificação alternativa
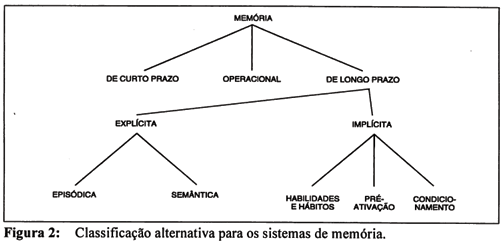
Quer parecer que elementos relevantes dos sistemas de memória não foram considerados na classificação de Squire e Zola-Morgan (1991). Por exemplo, a classificação refere-se apenas aos sistemas de memória de longo prazo, sem considerar o sistema de processamento de curto prazo. A distinção entre sistemas de curto e longo prazos parece natural. Além disso, a memória operacional não é incluída no modelo e deve, no nosso entender, constituir uma categoria a parte; além de receber informações dos sistemas de curto e longo prazos, ela contém os objetivos de processamento e ação em curso, possuindo, portanto, uma função integradora. Assim, na classificação proposta no presente trabalho (Figura 2), a memória pode ser subdividida em:
1. memória de curto prazo, correspondente a um sistema que armazena informações por curto intervalo de tempo, cuja capacidade é limitada, sendo a informação mantida por processos de atenção e ensaio;
2. memória operacional, um tipo de memória transitória que pode manter informações por períodos variáveis de tempo, em função da utilidade da informação. Este sistema tem acesso aos objetivos de processamento, planos de ação e é capaz de obter informações armazenadas nos sistemas de curto e longo prazos;
3. A memória de longo prazo pode ser adicionalmente subdividida em:
3.1. memória explícita, usualmente caracterizada pelo acesso consciente ao conteúdo da informação (embora este não seja sempre o caso), que compreende:
3.1.1. a memória episódica, para eventos e fatos experienciados em contexto espacial e temporal específicos, envolvendo informação autobiográfica e
3.1.2. memória semântica, para conhecimentos que independem do contexto, como conhecimentos gerais aritméticos, geográficos e históricos, e o significado de palavras e conceitos; neste último caso, depende da detecção dos elementos invariantes da situação, o que requer certa quantidade de repetições para a formação da síntese que expressa o conceito.
3.2. memória implícita, usualmente evidenciada através do desempenho e que corresponderia a alterações nos sistemas de processamento em função de sua utilização repetitiva. Esse processamento repetitivo resulta na automatização, tornando o posterior processamento independente de atenção. Depois de tornada automática, não há acesso consciente ao conteúdo da informação. A memória implícita compreende:
3.2.1. habilidades e hábitos, motores, perceptuais e cognitivos,
3.2.2. pré-ativação, viés ou facilitação do desempenho em função da apresentação prévia do material de teste, evidenciado através da capacidade para completar uma palavra apresentada anteriormente, ou de reconhecer mais prontamente palavras e figuras anteriormente apresentadas por apenas alguns milissegundos, e
3.2.3. condicionamento, expresso pela associação entre dois estímulos arbitrários em decorrência de sua apresentação pareada, fazendo com que a resposta inicialmente eliciada por apenas um dos estímulos passe a ser eliciada pelo outro.
Informações recebidas pelos órgãos sensoriais são processadas pelas estruturas nervosas a eles conectadas e aí retidas por um breve período de tempo, até que a informação seja adicionalmente codificada. Processos de atenção atuam neste estágio para manter a informação na memória de curto prazo. Em seguida, o material é codificado e mantido sob a forma de um armazenamento transitório, em função da utilidade da informação (na memória operacional); este sistema também possui acesso a objetivos de processamento, planos de ação e ao conteúdo já armazenado na memória explícita; o tempo de processamento da informação neste sistema é variável: depende, entre outras coisas, da natureza e relevância da informação processada, de experiências anteriores em relação a essa informação e de tentativas intencionais de sua evocação. Na eventual formação de memória explícita, o material é codificado de acordo com sua relevância, importância, saliência e familiaridade, e comparações com informações anteriormente coligidas são realizadas. Se a informação é considerada relevante, segue-se uma fase de consolidação, em que há uma reconstrução da experiência real para criar um registro permanente. Nesta fase, o material é revisado e modificado de acordo com outros conhecimentos e com base em experiências novas. Algumas das estruturas do lobo temporal estariam relacionadas com estas últimas etapas do processo, a partir da fase de consolidação; sua função, em associação com outras estruturas nervosas, seria a de manter o nível de atividade local nas estruturas de processamento inicial da informação, para desencadear os processos de alteração na eficiência sináptica até sua mudança permanente. Essa é a razão pela qual essas estruturas são necessárias a aprendizagens do tipo "treino único" ou à formação de memórias de eventos específicos. A independência de alguns dos sistemas de memória implícita em relação às estruturas do lobo temporal estaria associada ao fato de que, estabelecidos os objetivos de processamento e ação, a repetição de um dado processamento manteria, per se, o nível de atividade necessário, nas estruturas de processamento, ao desencadeamento dos processos responsáveis pela alteração da eficiência sináptica desses sistemas.
E. Um modelo neural para os módulos de memória
A arquitetura cerebral parece estar organizada sob a forma de módulos funcionais capazes de trabalhar cooperativa e independentemente. Esses módulos desempenham suas funções em paralelo. Através do processamento paralelo distribuído é possível que um grande número de unidades de processamento ativadas influencie outras em qualquer momento no tempo, de modo que as informações possam ser analisadas rapidamente.
A noção de processamento paralelo distribuído e de sistemas de ação (ver Morris, 1989 e Nadei et al., 1990) incorpora a idéia de que um sistema, por exemplo, o visual, processa informações separadamente das informações processadas pelo sistema auditivo. No próprio sistema visual, uma área (V1, p.ex.) processa informações separadamente de outra (V2) e mesmo dentro de uma determinada área, uma minicoluna pode ser considerada como um sistema funcional que processa informações separadamente das colunas ou minicolunas vizinhas. Isto não significa que os diferentes sistemas funcionais não cooperem em todos os níveis para produzir cognição e ação; apenas enfatiza que a enorme especificidade anatômica do sistema nervoso apóia múltiplos níveis de processamento de informação.
Esse mesmo raciocínio aplica-se aos sistemas de memória. A cooperação entre os diferentes sistemas proporciona a sensação de uma experiência de memória única, não evidenciando a profunda divisão de tarefas e de processamentos que ocorre para produzir a sensação unificada. Essa integração da informação requer que cada nível de uma vasta rede de conexões anatômicas contribua para a sensação final. Essa organização modular permite explicar dissociações experimentalmente observadas em indivíduos normais ou com lesões cerebrais ou desconexões. Em módulos que funcionam independentemente, lesões devem eliminar as funções por eles desempenhadas. Em módulos que cooperam, lesões devem alterar o desempenho dos módulos remanescentes, eventualmente minimizando as deficiências.
1. Funcionamento dos módulos e sua interação
Em função da existência de diversos sistemas de processamento e ação com conexões diferentes, o processamento em cada um destes sistemas certamente envolve tipos de informações distintas. Devido às diferenças entre a arquitetura destes sistemas, muito provavelmente eles manipulam a informação de forma diferente. Ademais, como a quantidade de sistemas,de processamento paralelo é grande, o acesso e controle voluntário a todos os aspectos envolvidos nesse processamento é, em alguns casos, dificultada, ocorrendo apenas em alguns módulos, em especial, nos quais não houve automatização no processamento da informação.
Todavia, podem existir aspectos comuns ao funcionamento desses diferentes sistemas. Mais especificamente, haveria, no sistema nervoso, uma grande quantidade de unidades dedicadas de processamento inerentemente plásticas, cada qual devotada a uma tarefa específica mas simples. Essas unidades são capazes de alterar seu funcionamento ontogeneticamente, ou por terem realizado processamentos anteriores, ou em função do resultado de processamentos realizados por outras unidades. Quando ativadas, essas unidades excitam e inibem outras ao longo de uma rica rede de conexões . Algumas acabam por formar ligações associativas cuja força pode ser alterada em função de diferentes fatores. A quantidade de neurotransmissores liberada com a chegada de cada potencial de ação pode ser alterada pela quantidade de cálcio que entra no terminal pré-sináptico, a probabilidade de ativação do receptor pós-sináptico, bem como a escala temporal dessa ativação, além do número de receptores funcionais e o próprio número e localização das sinapses podem ser modificados. Essas mudanças permitem explicar, entre outras coisas, a expansão das áreas corticais somatosensoriais relacionadas ao processamento de informações provenientes de três dedos intensamente estimulados associada à redução das áreas correspondentes aos dois dedos não estimulados, em macacos (Merzenich & Jenkins, 1993; Merzenich & Sameshima, 1993). Neste caso, o treinamento repetitivo foi capaz de induzir modificações nas estruturas de processamento em função de sua própria atividade. Algumas dessas alterações podem eventualmente requerer alterações permanentes na expressão de genes.
Nessa rede, conjuntos de nós podem representar informações da memória de longo prazo e a malha de ligações associativas das relações entre os nós. No caso da memória explícita, os nós podem ser ativados por processos sensoriais, codificando representações de eventos externos ou internos, ou ainda por objetivos e ações em curso. A ativação de um nó pode espalhar-se, pela ligação associativa, ativando outros nós da rede. Uma dada população de nós disparando, possivelmente com níveis de atividade diferentes em diferentes regiões corticais, pode representar uma determinada informação, cada região representando aspectos daquela informação (Figura 3A). Assim, um dado nó pode estar envolvido em representações diferentes, inclusive com atividades similares, já que a informação é representada pelo conjunto de disparos dos nós que se relacionam àquela informação e não por um nó individual (Figura 3B). Essa idéia é particularmente interessante, pois permite explicar a manutenção de representações, mesmo em casos de perda celular, como ocorre com o passar da idade: a permanência de sub-conjuntos de células constituintes das populações de nós garante que o processamento prossiga (Figura 3C). Ademais, permite explicar como uma informação constituída de componentes modalmente diferentes pode se constituir numa experiência única. No caso da memória implícita, o funcionamento seria similar, sendo que a ativação de um dos nós da rede desencadearia uma cascata de eventos que resultaria na ativação de nós cujas malhas tornaram-se fortalecidas pela ativação repetitiva. A ativação desse sistema dar-se-ia pela estimulação sensorial ou por objetivos de processamento ou ação; neste caso, o envolvimento de processamento explícito e/ou operacional durante as fases iniciais de aquisição do conhecimento implícito resultaria em ligações associativas entre os nós de ambos os sistemas; isto permite que objetivos e condições estabelecidos no sistema operacional acionem diretamente as estruturas de processamento do sistema implícito.
Por exemplo, durante uma conversa o ouvinte processa explicitamente e/ou operacionalmente o significado das palavras utilizadas pelo locutor mas processa implicitamente os princípios lingüísticos e fonológicos pelos quais o significado da prosa é decodificado. Durante a percepção de um ambiente, um indivíduo processa explicitamente dois objetos nele presentes, mas implicitamente os "cálculos" realizados para estabelecer que um está mais próximo ou é maior que o outro. Embora se tenha acesso explícito aos produtos do processamento paralelo realizado pelo sistema implícito, sendo possível referir-se a eles, os procedimentos e operações utilizados no processamento não são acessíveis ao sistema explícito por terem sido automatizados.
2. Sistemas de memória e o sistema nervoso
O nível de especificidade da relação entre os sistemas de memória e os circuitos do sistema nervoso está sendo atualmente estabelecido. Em alguns casos essa relação é surpreendente.
a. Memória de curto prazo
Pacientes com lesões nos giros supramarginal e angular do hemisfério esquerdo exibem prejuízos da memória verbal auditiva de curto prazo, mas desempenham normalmente tarefas que envolvem associação entre pares de palavras, lembrança de estórias e que envolvem memória implícita.
Esse tipo de resultado, em associação com dados sobre a formação de memória explícita (ver abaixo), sugere que as estruturas de processamento específico de um dado tipo de informação são as responsáveis pela sua manutenção por curtos períodos de tempo.
Assim, as regiões nervosas responsáveis pelo processamento inicial de informações, por exemplo, o córtex visual, auditivo e somatosensorial, manteriam essas informações por curtos períodos de tempo, até que fossem transferidas para outras estruturas que as processam adicionalmente.
b. Memória operacional
A memória operacional parece depender do córtex pré-frontal (e.g., Baddeley, 1992).
Lesões nesta região prejudicam o desempenho de macacos em tarefas que envolvem memória operacional (Goldman-Rakic, 1991), deixando intacta a capacidade para aprendizagem associativa.
Funahashi, Bruce e Goldman-Rakic (1989) mostraram que células do córtex pré-frontal de macacos apresentam um aumento na taxa de disparos para estímulos espacialmente localizados no campo visual durante o intervalo entre tentativas de uma tarefa de DMTS visual.
Esta região cortical é parte de uma complexa rede que envolve áreas sensoriais, límbicas (incluindo o hipocampo) e pré-motoras, aparentemente relacionada ao processamento de informações de natureza espacial. Goldman-Rakic (1992) sugeriu que o hipocampo teria o papel de contribuir no armazenamento de novas associações, enquanto o córtex pré-frontal teria a função de obter do armazenamento de longo prazo, situado em áreas de processamento modalmente específicas do sistema nervoso (ver abaixo), os resultados dessa aprendizagem associativa.
Thierry, Godbout, Mantz, Pirot e Glowinsk (1992) e Thierry, Mantz e Glowinski (1992), Berger, Nisenbaun, Strieker e Zigmond (1987), e Hökfelt, Skirboll, Reboket, Goldstein, Markey e Dann (1980), mostraram que o córtex pré-frontal de roedores possui grande quantidade de cateco-laminas. A queda dos níveis de dopamina no córtex pré-frontal resulta em prejuízos da memória operacional. Isto ajuda a explicar porque pacientes parkinsonianos têm dificuldades em tarefas que envolvem memória operacional; seu acesso à informação seria mais lenta em função da ausência de ativação dopaminérgica no córtex pré-frontal (ver abaixo, discussão sobre a relação entre memória implícita e operacional).
O cerebelo também parece estar envolvido no processamento da memória operacional. Pascual-Leone, Grafman, Clark, Stewart, Massaquoi, Low e Hallett (1993) propuseram que o cerebelo atua na catalogação e manutenção de eventos no tempo, atividades estas que seriam necessárias para manter listas de eventos na memória operacional, numa seqüência temporal adequada. Sobra ainda verificar se essa contribuição do cerebelo restringe-se apenas a tarefas que envolvem seqüências de movimentos (Inhoff, Diener, Rafai & Ivry, 1989; Ivry & Keele, 1988; Ivry & Keele, 1989) ou se aplica-se também a outras situações que requerem ordenamento temporal de informações (Ito, 1990; Bloedel, 1992).
c. Memória explícita
A mais popular das relações entre sistemas de memória e estruturas nervosas, como vimos, refere-se à necessidade da integridade de circuitos límbicos para o armazenamento de novo conhecimento explícito (e.g., Cohen, 1984; Squire & Zola-Morgan, 1991; Squire, 1992). Os elementos principais desse circuito estão representados na Figura 4.
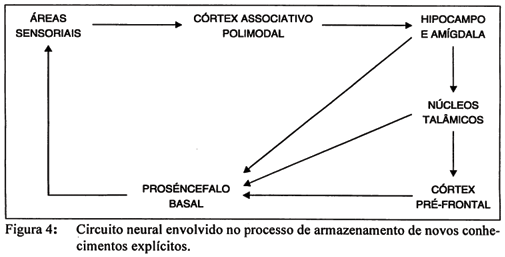
Resumidamente, eles incluem projeções de córtices de processamento polimodal (informações de natureza sensorial) para a amígdala e o hipocampo, deste para os núcleos talâmicos e hipotalâmicos, que por sua vez projetam-se para o córtex pré-frontal; esses últimos três conjuntos de estruturas projetam-se para o prosencéfalo basal (incluindo núcleos septais e núcleo basalis de Meynert) que projetam terminais colinérgicos para os tecidos corticiais relacionados ao processamento inicial e onde a informação é armazenada. Assim, haveria necessidade desse circuito para que a informação fosse armazenada. Depois de consolidada a informação, no entanto, o circuito límbico deixa de ser necessário para sua evocação.
O hipocampo, a amígdala e o córtex pré-frontal parecem centrais ao funcionamento desse circuito. Em humanos, o hipocampo direito parece relacionar-se com os aspectos espaciais da experiência de aprendizagem, enquanto o hipocampo esquerdo estaria associado a memória para material verbal (Smith & Milner, 1989). Por outro lado, a amígdala estaria relacionada com o estabelecimento de relações multimodais da experiência de aprendizagem e estados afetivos a elas relacionados (Kesner, Walser & Winzenried, 1989). Janowsky, Shinamura e Squire (1989) propuseram que o córtex pré-frontal seria importante na reconstrução da ordem temporal dos eventos passados (ver abaixo).
Lesões que desconectam estruturas de processamento sensoriais desse circuito límbico podem gerar amnésias modalmente específicas. Por exemplo, lesões bilaterais do córtex occipito-temporal interferem na transferência de informações visuais para o sistema límbico, dando origem a prejuízos de memória relacionados com a modalidade visual (Damasio, 1990). Aparentemente, esse resultado decorre da impossibilidade de o sistema límbico favorecer o armazenamento de informações nas regiões de processamento visual desconectadas pela lesão. Considerando-se que muitas das experiências sensoriais são polimodais, admite-se que os diferentes componentes modalmente específicos de uma dada experiência sejam armazenados, paralelamente, nos cortices sensoriais correspondentes ao seu processamento inicial; a evocação da experiência completa poderia ser iniciada pela ativação de qualquer dos seus componentes; evidentemente, quanto maior a quantidade de componentes ativados, maiores as chances de evocação, o que permite explicar efeitos de contexto, exaustivamente descritos na literatura sobre aprendizagem.
Depreende-se dessa proposta que lesões em regiões corticais específicas podem levar à perda de informações previamente adquiridas. Esse parece ser o caso. Por exemplo, lesões na área visual denominada " V4", responsável pelo processamento de cores, resultam em perda dessa faculdade. Além disso, os pacientes são incapazes de evocar da memória informações sobre cores experienciadas antes da lesão, embora sejam capazes de lembrar com precisão de formas, profundidade e movimento (Livingstone & Hubel, 1988).
Ainda sobre a relação entre estruturas nervosas e o sistema de memória explícita, alguns autores vêm tentando estabelecer correlações entre a atividade metabólica de regiões cerebrais, através de técnicas que permitem evidenciar a distribuição de sangue nas diferentes regiões, e o desempenho de tarefas que envolvem os diferentes sistemas de memória (e.g., Fazio, Perani, Gilardi, Colombo, Cappa & Vallar, 1992; Perani, Bressi, Cappa, Vallar, Alberoni, Grassi, Caltagirone, Cipolotti, Frances-chi, Lenzi & Fazio, 1993; Tulving, 1989). Por exemplo, Tulving (1989) mostrou que o fluxo sanguíneo no lobo frontal está aumentado durante a evocação de conhecimento episódico, mas não durante a evocação de conhecimento semântico. Fazio et al. (1992) mostraram que o hipocampo, o tálamo, o córtex cingular e o córtex frontal basal exibem aumento de atividade metabólica em tarefas que envolvem memória episódica. Já Petersen, Fox, Posner, Mintum e Raichle (1988) mostraram que o córtex associativo frontal esquerdo e a área cingular exibem aumento de atividade em tarefas que envolvem memória semântica, embora não esteja descartada a possibilidade de que esse aumento esteja relacionado com a seleção da resposta adequada para o desempenho da tarefa (Frith, Friston, Liddle & Frackowiak, 1991).
d. Memória implícita
A aquisição de conhecimento implícito está afetada em pacientes com disfunções cerebelares (Schmahmann, 1991; Lalonde & Botez, 1990; Bloedel, Bracha, Kelly et al., 1991), em pacientes com a doença de Huntington (Knopman & Nissen, 1991) e, em algumas tarefas que envolvem memória implícita, em pacientes com a doença de Parkinson (Harrington, Haaland, Yeo & Marder, 1990). Isto sugere que as estruturas nervosas afetadas nesses pacientes relacionam-se com os sistemas de memória implícita.
A aprendizagem de habilidades motoras parece depender de aferências corticais de áreas sensoriais de associação para o estriado ou gânglios basais (Mishkin, Malamut & Bachavalier, 1984). O caudado e o putamen (estruturas-chave do estriado) recebem grandes projeções corticais e projetam-se para o globo pálido e estruturas do sistema extra-piramidal, podendo constituir um dos elos da aquisição de conhecimento implícito que envolva conexões entre estímulos e respostas. Congruentemente, Seitz, Roland, Bohm, Greitz et al. (1990) descreveram alterações no fluxo sanguíneo no cerebelo e gânglios basais durante a aprendizagem de tarefas que envolvem movimentos complexos dos dedos da mão. Inicialmente, houve aumento do fluxo sanguíneo no cerebelo e decréscimo de fluxo no estriado; no processo de aquisição da habilidade, houve aumento paulatino de fluxo no estriado. Não é surpreendente, portanto, que a aquisição desse tipo de habilidade motora esteja afetada em pacientes portadores da doença de Parkinson. Todavia, esses mesmos pacientes são capazes de adquirir habilidades perceptuais visuais (Harrington et al., 1990); essa dissociação sugere que as estruturas nervosas relacionadas com a aquisição de habilidades motoras diferem das envolvidas na aquisição de habilidades perceptuais. Estas últimas provavelmente relacionam-se com as estruturas de processamento específico da informação.
Além do envolvimento na aquisição da habilidade propriamente dita, parece haver um envolvimento do neo-estriado (Saint-Cyr, Taylor & Lang, 1988) e do cerebelo (Grafman, Litvan, Massaquoi, Steward, Sirigu & Hallett, 1992) na aquisição do planejamento cognitivo que envolve o desempenho da ação. Assim, estas estruturas poderiam constituir o elo entre os sistemas implícito, operacional e explícito, devido às suas ligações com elementos destes últimos, através do tálamo e dos lobos frontais (Schmahmann, 1991; Leiner, Leiner & Dow, 1991). Nesse sentido, Ja-hanshani, Brown e Marsden (1992) mostraram que as dificuldades dos pacientes parkinsonianos estão relacionadas com o início do movimento, o que poderia ser decorrência das dificuldades de induzir a excitação cortical que antecede o desempenho da ação. O mesmo se aplica à atividade do córtex pré-frontal; o decréscimo dos níveis de dopamina tornaria mais lento seu funcionamento, resultando em dificuldades na memória operacional.
F. Uma palavrinha sobre a relação entre memória e consciência
A habilidade para relacionar uma grande quantidade de informações, passadas, presentes e antecipadas, proporciona flexibilidade ao controle comportamental, parecendo ser esta a principal vantagem evolutiva para o aparecimento do que se costuma chamar de consciência; ela parece estar baseada em atividades concomitantes de (ou ter acesso a) módulos distribuídos nas redes neurais.
Muitas destas características, como vimos, estão relacionadas com as funções do córtex pré-frontal, mais especificamente do córtex pré-frontal granular. Esta estrutura sofreu um visível aumento de área ao longo da evolução filogenética, tendo se tornado um dos mais proeminentes componentes do cérebro humano. Mesulam (1986) sugeriu que sua função está associada à organização de processos cognitivos como julgamento, perspicácia, curiosidade, abstração e criatividade. Suas conexões com outras estruturas do sistema nervoso são muito diversificadas. Nesse sentido, esta estrutura não só recebe informações sobre uma ampla gama de processamentos que ocorrem nos diferentes módulos do sistema nervoso, como pode influenciar toda essa atividade, estimulando o funcionamento de alguns módulos e inibindo o funcionamento de outros.
Parece ter ficado claro nas discussões sobre sistemas de memória que, dada a natureza modular do processamento de informações no sistema nervoso, as informações sobre um determinado objeto ou evento são distribuídas continuamente através da rede de processamento, não sendo necessário que essa informação esteja plenamente (conscientemente) representada para que possa influenciar a experiência, pensamento ou ação em curso, embora, sob determinadas condições, ela possa tornar-se consciente.
Sob este prisma, as características e o substrato anatômico possivelmente relacionados com a consciência assemelham-se aos envolvidos na memória operacional. Isto tem levado alguns autores a proporem que a consciência e a memória operacional correspondem ao mesmo processo (e.g., Baddeley, 1992).
Alternativamente, Gazzaniga (1988), baseado em estudos de pacientes com secção do corpo caloso, propôs que o monitoramento de toda essa atividade, em humanos, é feito por um sistema, denominado interpretador, baseado no hemisfério esquerdo. Esse interpretador avaliaria os resultados de processamento dos módulos funcionais assim que ocorrem e imediatamente construiria uma hipótese sobre o porque de determinadas ações terem sido realizadas. O interpretador considera o comportamento como forte evidência dos resultados de processamento de vários módulos funcionais envolvidos com a atividade em curso e os interpreta em função de uma pré-concepção nele construída. Se o resultado de processamento de um módulo afeta um dado comportamento de forma dissonante em relação à pré-concepção estabelecida através de ações interpretativas prévias, o comportamento mudará a pré-concepção ou levará o interpretador a tentar encontrar "justificativas" para a discrepância. Em outras palavras, o interpretador tenta relacionar as atividades e dar-lhes um sentido lógico. Essa função interpretativa trabalharia sobre os produtos das atividades modulares para construir um esquema que possa explicar a lógica subjacente a toda essa atividade, que resulta num determinado comportamento. O comportamento pode, portanto, tornar-se um poderoso determinante daquilo que seu agente acredita ser verdadeiro.
Seria prematuro favorecer uma destas propostas. Todavia, o desafio de fazê-lo ou mesmo de propor uma hipótese alternativa deverá ser enfrentado.
Conclusão
O sistema nervoso está organizado de forma estritamente modular. Cada módulo mantém conexões diferentes com os outros e o nível de independência no funcionamento desses módulos parece variar. Lesões em módulos independentes eliminam as funções por eles desempenhadas, enquanto lesões em módulos que cooperam resultam numa alteração de desempenho dos módulos remanescentes de modo a minimizar as deficiências. O funcionamento independente porém cooperativo entre os diferentes módulos permite explicar não só dissociações experimentalmente observadas em indivíduos com lesões cerebrais, mas também a sensação de uma experiência de memória única em indivíduos normais.
REFERÊNCIAS BIBLIOGRÁFICAS
BADDELEY, A.D. Working memory. Science, v.255, n.5044, p.556-9, 1992. [ Links ]
BADDELEY, A.D.; WARRINGTON, E.K. Amnesia and the distinction between long-and short-term memory. Journal of Verbal Learning and Verbal Behavior, v.9, n.2, p. 176-89, 1970. [ Links ]
BERGER, T.W.; NISENBAUM, E.S.; STRICKER, E.M; ZIGMOND, M.J. Evidence for two functionally distinct subpopulations of neurons within the striatum and their differential sensitivity to dopamine. In: CHIODO, L.A; FREEMAN, A.S., eds. Neurophysiology of dopaminergic systems: current status and clinical perspectives. Grosse Point, Lakeshore Publishing, 1987. p.253-84. [ Links ]
BISIACH, E. The (haunted) brain and consciousness. In: MARCEL, A.J.; BISIACH, E., eds. Consciousness in contemporary science. Oxford, Clarendon Press, 1988. p.101-20. [ Links ]
BISIACH, E.; BERTI, A.; VALLAR, G. Analogical and logical disorders underlying unilateral neglect of space. In: POSNER, M.I.; MARIN, O.S.M., eds. Attention and performance XI. Hillsdale, Lawrence Erlbaum, 1985. p.239-49. [ Links ]
BLOEDEL, J.R. Functional heterogeneity with structural homogeneity: how does the cerebellum operate? Behavioural and Brain Sciences, v.15, n.4, p.666-78, 1992. [ Links ]
BLOEDEL, J.R.; BRACHA, V.; KELLY, T.M. et al. Substrates for motor learning. Does the cerebellum do it all? Annual New York Academic Science, v.627, p.305-18, 1991. [ Links ]
BUTTERS, N. Procedural learning in dementia: a double dissociation between Alzheimer and Huntington's disease patients on verbal priming and motor skill learning. In: MEETING OF THE INTERNATIONAL NEUROPSYCHOLOGICAL SOCIETY, Washington, 1987. Abstracts. Washington, 1987. p.305. [ Links ]
COHEN, N.J. Preserved learning capacity in amnesia: evidence for multiple memory systems. In: SQUIRE, L.R.; BUTTERS, N., eds. The neuropsychology of memory. New York, Guilford Press, 1984. p.83-103. [ Links ]
COHEN, N.J.; SQUIRE, L.R. Preserved learning and retention of pattern analyzing skill in amnesia: dissociation of knowing how and knowing that. Science, v.210, n.4466, p.207-10, 1980. [ Links ]
COLLIER, T.J.; QUIRK, G.J.; ROUTTENBERG, A. Separable roles of hippocampal granule cells in forgetting and pyramidal cells in remembering spatial information. Brain Research, v.409, n.2, p.316-28, 1987. [ Links ]
CRAIK, F.I.M.; LOCKHART, R.S. Levels of processing: a framework for memory research. Journal of Verbal Learning and Verbal Behavior, v.11, n.6, p.671-84,1972. [ Links ]
DAMASIO, A.R. Category-related recognition defects as a clue to the neural substrates of knowledge. Trends in Neurosciences, v.13, n.3, p.95-8, 1990. [ Links ]
DAUM, I.; CHANNON, S.; CANAVAN, A. Classical conditioning in patients with severe memory problems. Journal of Neurology and Neurosurgery Psychiatry, v.52, n.1,p.47-51, 1989. [ Links ]
DELIS, D.C. Neuropsychological assessment of learning and memory. In: SQUIRE, L.R., ed. Memory and its disorders. New York, Elsevier Science, 1989. p.3-33. [ Links ]
EBBINGHAUS, H. Memory. New York, Dover, 1964. [ Links ]
ERDELYI, M.H. Hypermnesia and insight. In: MARCEL, A.J.; BISIACH, E., eds. Consciousness in contemporary science. Oxford, Clarendon Press, 1988. p.200-17. [ Links ]
ERDELYI, M.H.; KLEINBARD, J. Has Ebbinghaus decayed with time? The growth of recall (hypermnesia) over days. Journal of Experimental Psychology: Human Learning and Memory, v.4, n.4, p.275-89, 1978. [ Links ]
FAZIO, F.; PERANI, D.; GILARDI, M.C.; COLOMBO, F.; CAPPA, S.F.; VALLAR, G. Metabolic impairment in human amnesia: a PET study of memory networks. Journal of Cerebral Blood Flow and Metabolism, v.12, p.353-8, 1992. [ Links ]
FRITH, C.D.; FRISTON, K.; LIDDLE, P.F.; FRACKOWIAk, R.S.J. Willed action and the prefrontal cortex in man: a study with PET. Proceedings of the Royal Society of London: Sect.B, v.244, p.241-6, 1991. [ Links ]
FUNAHASHI, S.; BRUCE, C.J.; GOLDMAN-RAKIC, P.S. Mnemonic coding of visual space in the monkey's dorsolateral prefrontal cortex. Journal of Neurophysiology, v.61,n.2,p.331-49, 1989. [ Links ]
GAZZANIGA, M.S. Brain modularity: towards a phylosophy of conscious experience. In: MARCEL, A.J.; BISIACH, E., eds. Consciousness in contemporary science. Oxford, Clarendon Press, 1988. p.218-38. [ Links ]
GAZZANIGA, M.S. Right hemisphere language: a twenty year perspective. The American Psychologist, v.38, n.5, p.525-37. 1983. [ Links ]
GAZZANIGA, M.S.; HOLTZMAN, J.D.; SMYLIE, C.S. Speech without conscious awareness. Neurology, v.35, n.4, p.682-5, 1987. [ Links ]
GAZZANIGA, M.S.; SMYLIE, C.S.; BAYNES, K.; HIRST, W.; McCLEARY, C. Profiles of right hemisphere language and speech following brain bisection. Brain and Language, v.22, n.2, p.206-20, 1984. [ Links ]
GLISKY, E.L.; SCHACTER, D.L.; TULVING, E. Learning and retention for computer-related vocabulary in memory-impaired patients: method of vanishing cues. Journal of Clinical and Experimental Neuropsychology, v.8, n.3, p.292-312, 1986. [ Links ]
GOLDMAN-RAKIC, P.S. La memoria funcional y la mente. Investigación y Ciencia, p.68-75,nov. 1992. [ Links ]
GOLDMAN-RAKIC, P.S. Prefrontal cortical dysfuncion in schizophrenia: the relevance of working memory. In: CARROLL, B.J; BARRETT, J.E., eds. Psychopathology and the brain, s.l., Raven Press, 1991. p.322-51. [ Links ]
GRAF, P.; SCHACTER, D.L. Implicit and explicit memory for new associations in normal and amnesic subjects. Journal of Experimental Psychology: Learning, Memory, and Cognition, v.11, n.3, p.501-18, 1985. [ Links ]
GRAFMAN, J.; LITVAN, I.; MASSAQUOI, S.; STEWARD, M.; SIRIGU, A.; HALLETT, M. Cognitive planning deficit in patients with cerebellar atrophy. Neurology, v.42, n.8, p. 1493-6, 1992. [ Links ]
GRANT, D.S. Proactive interference in pigeon short-term memory. Journal of Experimental Psychology: Animal Behavior Processes, v.104, n.3, p.207-20, 1975. [ Links ]
HABER, R.N.; ERDELYI, M.H. Emergence and recovery of initially unavailable perceptual material. Journal of Verbal Learning and Verbal Behavior, v.6, p.618-28, 1967. [ Links ]
HARRINGTON, D.L.; HAALAND, K.Y.; YEO, R.A.; MARDER, E. Procedural memory in Parkinson's disease: impaired motor but not visuoperceptual learning. Journal of Clinical Experimental Neuropsychology, v.12, n.2, p.323-39, 1990. [ Links ]
HIRSH, R. The hippocampus and contextual retrieval of information from memory: a theory. Behavioral Biology, v.12, n.4, p.421-44, 1974. [ Links ]
HÖKFELT, T.; SKIRBOLL, L.; REBOKET, J.F.; GOLDSTEIN, M.; MARKEY, K.; DANN, O. A subpopulation of mesencephalic dopamine neurons projecting to limbic area contains a cholecystokinin-like peptide, evidence from immunohisto-chemistry combined with retrograde tracing. Neuroscience, v.5, p.2093-2124, 1980. [ Links ]
HONIG, W.K. Studies of working memory in the pigeon. In: HULSE, S.H.; FOWLER, H.; HONIG, W.K., eds. Cognitive processes in animal behavior. Hillsdale, Lawrence Erlbaum, 1978. p.211-48. [ Links ]
HOREL, J.A.; VOYTKO, M.L.; SALSBURY, K.G. Visual learning suppressed by cooling the temporal pole. Behavioral Neuroscience, v.98, n.2, p.310-24, 1984. [ Links ]
INHOFF, A.W.; DIENER, H.C.; RAFAL, R.D.; IVRY, R. The role of cerebellar structures in the execution of serial movements. Brain, v.112, p.565-81, 1989. [ Links ]
ITO, M. A new physiological concept on cerebellum. Revue Neurologique, v.146, p.564-9, 1990. [ Links ]
IVRY, R.; KEELE, S.W. Dissociation of the lateral and medial cerebellum in movement timing and movement execution. Experimental Brain Research, v.73, p. 167-80, 1988. [ Links ]
IVRY, R.; KEELE, S.W. Timing function of the cerebellum. Journal of Cognitive Neuroscience, v.1, p. 136-52, 1989. [ Links ]
JACOBY, L.L.; DALLAS, M. On the relationship between autobiographical memory and perceptual learning. Journal of Experimental Psychology: General, v.3, p.306-40, 1981. [ Links ]
JAHANSHANI, M.; BROWN, R.G.; MARSDEN, C.D. Simple and choice reaction time and the use of advance information for motor preparation in Parkinson's disease. Brain, v.115, p.539-64, 1992. [ Links ]
JANOWSKY, J.S.; SHINAMURA, A.P.; SQUIRE, L.R. Source memory impairment in patients with frontal lobe lesions. Neuropsychologia, v.27, n.8, p. 1043-56, 1989. [ Links ]
KESNER, R.P.; WALSER, R.D.; WINZENRIED, G. Central but not basolateral amygdala mediates memory for positive affective experiences. Behavioral Brain Research, v.33, n.2, p. 189-95, 1989. [ Links ]
KIHLSTROM, J.F. Posthypnotic amnesia for recently learned materials: interactions with "episodic" and "semantic" memory. Cognitive Psychology, v.12, n.2, p.227-51, 1980. [ Links ]
KNOPMAN, D.S.; NISSEN, M.J. Procedural learning is impaired in Huntington's disease: from the serial reaction time task. Neuropsychologia, v.29, n.3, p.245-54, 1991. [ Links ]
LALONDE, R.; BOTEZ, M.I. The cerebellum and learning processes in animals. Brain Research Reviews, v.15, n.3, p.325-32, 1990. [ Links ]
LEINER, H.C.; LEINER, A.L.; DOW, R.S. The human cerebo-cerebellar system: its computing, cognitive, and language skills. Behavioural Brain Research, v.44, n.2, p.113-28, 1991. [ Links ]
LIVINGSTONE, M.; HUBEL, D. Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science, v.240, n.4853, p.740-9, 1988. [ Links ]
MERZENICH, M.M.; JENKINS, W.M. Reorganization of cortical representations of the hand following alterations of skin inputs induced by nerve injury, skin island transfers, and experience. Journal of Hand Therapy, v.6, n.2, p.89-104, 1993. [ Links ]
MERZENICH, M.M; SAMESHIMA, K. Cortical plasticity and memory. Current Opinion in Neurobiology, v.3, n.2, p.187-96, 1993. [ Links ]
MESULAM, M. Frontal cortex and bahavior. Annals of Neurology, v.19, p.320-5, 1986. [ Links ]
MESULAM, M. Large-scale neurocognitive networks and distributed processing for attention, language, and memory. Annals of Neurology, v.28, n.5, p.597-613, 1990. [ Links ]
MEUDELL, P.; MAYES, A. The clapatede phenomenon: a further example in amnesics, a demonstration of a similar effect in normal people with attenuated memory, and a reinterpretation. Current Psychological Research, v.1, p.75-88, 1981. [ Links ]
MISHKIN, M. Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature, v.273, n.5660, p.297-8, 1978. [ Links ]
MISHKIN, M.; APPENZELLER, T. The anatomy of memory. Scientific American, v.256, n.6, p.80-9, 1987. [ Links ]
MISHKIN, M.; MALAMUT, B.; BACHEVALIER, J. Memories and habits: two neural systems. In: LYNCH, G.; McGAUGH, J.L.; WEINGARTEN, N.M., eds. Neurobiology of learning and memory. New York, Guilford Press, 1984. p.65-77. [ Links ]
MISHKIN, M.; PETRI, H.L. Memories and habits: some implications for the analysis of learning and retention. In: SQUIRE, L.R.; BUTTERS, N., eds. The neuropsychology of memory. New York, Guilford Press, 1984. p.287-96. [ Links ]
MORRIS, R.G.M., ed. Parallel distributed processing. Implications for psychology and neurobiology. Oxford, Clarendon Press, 1989. [ Links ]
MURRAY, E.A.; MISHKIN, M. Visual recognition in monkeys following rhinal cortical ablations combined with either amygdalectomy or hippocampectomy. Journal of Neuroscience, v.6, p. 1991-2003, 1986. [ Links ]
NADEL, L.; COOPER, L.A.; CULICOVER, P.; HARNISH, R.M., eds. Neural connections, mental computation. Cambridge, M.I.T. Press, 1990. [ Links ]
NELSON, T.O. Detecting small amounts of information in memory: saving for nonrecognized items. Journal of Experimental Psychology: Human, Learning, and Cognition, v.4, n.5, p.453-68, 1978. [ Links ]
OLTON, D.S. Dimensional mnemonics. The Psychology of Learning and Motivation, v.24,p.1-23, 1989. [ Links ]
OLTON, D.S. Memory functions and the hippocampus. In: SEIFERT, W., ed. Neurobiology of the hippocampus. New York, Academic Press, 1983. [ Links ]
OLTON, D.S.; BECKER, J.T.; HANDELMAN, G.E. Hippocampus, space and memory. Behavioral and Brain Sciences, v.2, n.3, p.315-65, 1979. [ Links ]
OLTON, D.S.; SAMUELSON, R.J. Remembrance of places passed: spatial memory in rats. Journal of Experimental Psychology: Animal Behavior Processes, v.2, n.2, p.97-116, 1976. [ Links ]
PASCUAL-LEONE, A.; GRAFMAN, J.; CLARK, K. STEWART, M.; MASSAQUOI, S.; LOW, J.S.; HALLETT, M. Procedural learning in Parkinson's disease and cerebellar degeneration. Annals of Neurology, v.34, p.594-602, 1993. [ Links ]
PERANI, D.; BRESSI, S.; CAPPA, S.F.; VALLAR, G.; ALBERONI, M.; GRASSI, F.; CALTAGIRONE, C.; CIPOLOTTI, L.; FRANCESCHI, M.; LENZI, G.L.; FAZIO, F. Evidence of multiple memory systems in the human brain. Brain, v.116, p.903-19, 1993. [ Links ]
PETERSEN, S.E.; FOX, P.T.; POSNER, M.I.; MINTUM, M.; RAICHLE, M.E. Positron emission tomographic studies of the cortical anatomy of single-word processing. Nature, v.331, p.585-9, 1988. [ Links ]
PRESS, G.A.; AMARAL, D.G.; SQUIRE, L.R. Hippocampal abnormalities in amnesic patients revealed by high-resolution magnetic resonance imaging. Nature, v.341, p.54-7, 1989. [ Links ]
PRIBRAM, K.H. The hippocampal system and recombinant processing In: ISAACSON, R.L.; PRIBRAM, K.H., eds. The hippocampus. New York, Plenum Press, 1986. v.4, p.329-70. [ Links ]
RYLE, G. The concept of mind. San Francisco, Hutchinson, 1949. [ Links ]
SAINT-CYR, J.; TAYLOR, A.E.; LANG, A.E. Procedural learning and neostriatal dysfunction in man. Brain, v.111, p.941-59, 1988. [ Links ]
SCHACTER, D.L. Implicit memory: history and current status. Journal of Experimental Psychology: Learning, Memory, and Cognition, v.13, n.3, p.501-18, 1987. [ Links ]
SCHACTER, D.L. Multiple forms of memory in humans and animals. In: WEINGARTEN, N.; LYNCH, G.; McGAUGH, J.L., eds. Memory systems of the brain: animal and human cognitive processes. New York, Guilford Press, 1985. p.351-79. [ Links ]
SCHACTER, D.L.; GRAF, P. Preserved learning in amnesic patients: perspectives from research on direct priming. Journal of Clinical and Experimental Psychology, v.8, n.6, p.727-43, 1986. [ Links ]
SCHACTER, D.L.; HARBLUK, J.L.; McLACHLAN, D.R. Retrieval without recollection: an experimental analysis of source amnesia. Journal of Verbal Learning and Verbal Behavior, v.23, n.5, p.593-611, 1984. [ Links ]
SCHACTER, D.L.; MOSCOVITCH, M. Infants, amnesics, and dissociable memory systems. In: MOSCOVITCH, M., ed. Infant memory. New York, Plenum Press, 1984. p.173-216. [ Links ]
SCHACTER, D.L.; TULVING, E. Amnesia and memory research. In: CERMARK, L.S., ed. Human memory and amnesia. Hillsdale, Erlbaum, 1982. [ Links ]
SCHMAHMANN, J.D. An emerging concept. The cerebellar contribution to higher function. Archives of Neurology, v.48, n.11, p. 1178-87, 1991. [ Links ]
SCOVILLE, W.B.; MILNER, B. Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery, and Psychiatry, v.20, p.11-21, 1957. [ Links ]
SEITZ, R.J.; ROLAND, E.; BOHM, C.; GREITZ, T. et al. Motor learning in man: a positrom emission tomographic study. Neuroreport: An International Journal for the Rapid Communication of Research in Neuroscience, v.1, n.1, p.57-60, 1990. [ Links ]
SHERRY, D.F.; SCHACTER, D.L. The evolution of multiple memory systems. Psychological Review, v.94, n.4, p.439-54, 1987. [ Links ]
SLAMECKA, N.J. Ebbinghaus: some associations. Journal of Experimental Psychology: Learning, Memory, and Cognition, v.11, n.3, p.414-35, 1985. [ Links ]
SMITH, M.L.; MILNER, B. Right hippocampal impairment in the recall of spatial location: encoding deficit or rapid forgetting? Neuropsychologia, v.27, n.1, p.71-81, 1989. [ Links ]
SQUIRE, L.R. Memory and brain. New York, Oxford University Press, 1987. [ Links ]
SQUIRE, L.R. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychological Review, v.99, n.3, p. 195-231, 1992. [ Links ]
SQUIRE, L.R.; COHEN, N.J. Human memory and amnesia. In: LYNCH, G.; McGAUGH, J.L.; WEINGARTEN, N.M., eds. Neurobiology of learning and memory. New York, Guilford Press, 1984. p.3-64. [ Links ]
SQUIRE, L.R.; ZOLA-MORGAN, S. The medial temporal lobe memory system. Science, v.253, n.5026, p. 1380-6, 1991. [ Links ]
THIERRY, A.M.; GODBOUT, R.; MANTZ, J.; PIROT, S.; GLOWINSKI, J. Differential influence of dopaminergic and noradrenergic afferents on their target cells in the rat prefrontal cortex. Clinical Neuropharmacology, v.15, p.139A-40A, 1992. Supplement 1. [ Links ]
THIERRY, A.M.; MANTZ, J.; GLOWINSKI, J. Influence of dopaminergic and noradrenergic afferents on their target cells in the rat medial prefrontal cortex. Advances Neurology, v.57, p.545-54, 1992. [ Links ]
THOMAS, K. Man and the natural world. Harmondsworth, Penguin Books, 1984. [ Links ]
TULVING, E. Elements of episodic memory. Oxford, Oxford University Press, 1983. [ Links ]
TULVING, E. Episodic and semantic memory. In: TULVING, E.; DOANLDSON, W., eds. Organization of memory. New York, Academic Press, 1972. p.381-403. [ Links ]
TULVING, E. Remembering and Knowing the past. American Scientist, v.77, p.361-7, 1989. [ Links ]
WARRINGTON, E.K.; WEISKRANTZ, L. Amnesia: a disconnection syndrom? Neuropsychologia, v.17, p. 187-94, 1982. [ Links ]
WEISKRANTZ, L. Some contributions of neuropsychology of vision and memory to the problem of consciousness. In: MARCEL, A.; BISIACH, E., eds. Consciousness and contemporary science. New York, Oxford University Press, 1988. p. 183-9. [ Links ]
WINNICK, W.A.; DANIEL, S.A. Two kinds of response priming in tachistoscopic recognition. Journal of Experimental Psychology, v.84, n.1, p.74-81, 1970. [ Links ]
WORSHAM, R.W. Temporal discrimination factors in the delayed-matching-to-sample task in monkeys. Animal Learning and Behavior, v.3, n.2, p.93-7, 1975. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R. Medial temporal lesions in monkeys impair memory in a variety of tasks sensitive to human amnesia. Behavioral Neuroscience, v.99, n.1,p.22-34, 1985. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R.; AMARAL, D.G. Human amnesia and the medial temporal region: enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. Journal of Neuroscience, v.6, n.10, p.2950-67, 1986. [ Links ]
ZOLA-MORGAN, S.; SQUIRE, L.R.; MISHKIN, M. The neuroanatomy of amnesia: amygdala-hippocampus versus temporal stem. Science, v.218, n.4579, p. 1337-9, 1982. [ Links ]
1 O autor agradece a Andréa Maria Garrido dos Santos, Maria Cristina Magila, Marina Milos e, em particular, ao Prof. Dr. César Ades, pelas críticas e sugestões à versão preliminar deste trabalho.
2 Mishkin et al. (1984) apresentaram uma distinção similar entre "memórias e hábitos" , enfatizando as mesmas características acima descritas para memória declarativa e de procedimento, respectivamente. De acordo com essa proposta, o sistema controlador do armazenamento de memória envolveria projeções do hipocampo e amígdala para estruturas diencefálicas, destas últimas para o córtex pré-frontal, e de todas estas estruturas para o prosencéfalo basal, que por sua vez projeta-se para o córtex sensorial ativado quando da entrada da informação, no qual ocorreria o armazenamento da informação, sob o controle daquele circuito (para detalhes, ver Mishkin & Appenzeller, 1987).
