










Serviços Personalizados
artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Compartilhar

 Permalink
PermalinkPsicologia USP
versão On-line ISSN 1678-5177
Psicol. USP v.6 n.1 São Paulo 1995
ARTIGOS ORIGINAIS
Organização do trabalho em colônias de saúvas: relações com locais e objetos das atividades1
The organization of labour in ant colonies: relations with places and objects of activities
Ana Maria Almeida Carvalho
Instituto de Psicologia - USP
RESUMO
Este trabalho relata os resultados principais de um projeto de pesquisa sobre divisão de trabalho em colônias artificiais da saúva. Atividades desempenhadas dentro e fora da colônia foram analisadas em termos de sua distribuição entre classes de obreiras, definidas pelo tamanho e cor do tegumento. Os resultados mostram uma divisão quantitativa de trabalho entre obreiras de tamanhos diferentes e sugerem a divisão da casta de obreiras em quatro sub-castas definidas pelo objeto predominante de atividade. Quanto à cor, indicador de idade, a distribuição mostrou-se mais afetada pelo local de desempenho. Uma manipulação experimental do acesso das obreiras a áreas externas não modificou a distribuição de atividades relativas a corte e transporte de vegetais entre classes de tamanho. São discutidas as implicações desses resultados para a compreensão dos mecanismos de organização do trabalho em colônias de formigas, e sugeridas direções para a investigação experimental desse tema.
Descritores: Divisão de trabalho animal. Ambiente animal. Formigas. Diferenças de idade. Tamanho do corpo. Insetos. Comportamento social animal.
ABSTRACT
This paper reports the main results of a research project about the division of labour in laboratory colonies of the fungus-growing ant Atta sexdens rubropilosa. Activities performed inside and outside the colonies were cross-analyzed in terms of its distribution according to size and age classes of workers. The results show a quantitative division of labour among workers of different sizes, and suggest a division of the workers’ caste into four size-related sub-castes according to the prevailing object of their activities. As to age classes, place appears as the main factor in the distribution of activities, younger workers performing mostly internal tasks. The results of an experimental manipulation of the workers access to external areas did not affect the distribition of leaf cutting and leaf transportation among workers of different size classes. Implications of these results to the understanding of the mechanisms of labour organization in ant colonies are discussed, and directions for further experimental investigations on the topic are suggested.
Index terms: Animal division of labor. Animal environments. Ants. Age differences. Body size. Insects. Animal social behavior.
A sobrevivência de um grupo social, tal como a de um animal tomado individualmente, requer uma variedade de tarefas para garantir sua alimentação, sua reprodução, sua defesa. Quando o grupo é tomado como unidade de análise, uma pergunta inevitável é: que mecanismos subjazem à organização e distribuição dessas tarefas, de forma a garantir que todas sejam realizadas da maneira e com a freqüência necessárias para a sobrevivência do grupo? Esta pergunta é especialmente intrigante quando se trata de grupos sociais com organização complexa, mas situados nos degraus inferiores da escala evolutiva, como é o caso das sociedades de insetos.
Sob os nomes de divisão de trabalho, polietismo ou diferenças individuais de comportamento, a distribuição de tarefas em colônias de insetos tem sido estudada ou mencionada por muitos autores. A literatura concorda sobre a existência de divisão de trabalho nas sociedades de insetos mais evoluídas, e já se fez mesmo a caracterização dessas sociedades com base nesse fenômeno: "Os insetos estritamente sociais, isto é, aqueles que exibem divisão de trabalho (...)" (Allee et al., 1965, p.420, grifo meu).
Em formigas, a divisão de trabalho é em geral considerada menos clara , menos rígida e menos fisiologicamente determinada do que em abelhas (Lindauer, 1952; Bernard, 1968; Goetsch, 1957). Há no entanto uma grande variedade de referências ao fenômeno na família Formicidae, em geral relacionando-o a características estruturais (polimorfismo, especialmente quanto ao tamanho das obreiras) ou ontogenéticas (idade, diversidade de experiências e conseqüentes diferenças individuais).
Diferentemente do que ocorre nas abelhas, em muitas espécies de formigas o polimorfismo entendido como existência de mais de uma forma na mesma espécie não se limita a uma diferenciação entre castas férteis (rainhas e machos) e estéreis (operárias), mas ocorre também entre estas últimas, sob duas formas: dimorfismo - ocorrência de duas sub-castas de obreiras claramente diferenciadas pelo tamanho - "mínimas" e "máximas", como em Pheidole; e polimorfismo propriamente dito - em que o tamanho das obreiras difere ao longo de um contínuo, sem limites precisos entre as sub-castas, como nos gêneros Camponotus, Messor e Atta (Wheeler, 1910).
A seqüência provável de evolução do polimorfismo, segundo Wilson (1953), foi uma diferenciação inicial, a partir da rainha, ao longo de um gradiente de tamanhos, e posteriormente a segregação de dois grupos de tamanhos bem diferentes, com a eliminação de tamanhos intermediários, resultando em espécies dimórficas. As espécies propriamente polimórficas representariam portanto uma fase de transição nessa seqüência evolutiva.
Supõe-se ainda que o valor adaptativo do polimorfismo estaria na especialização morfológica dele decorrente, que por sua vez favoreceria a especialização de funções.
Todos os autores concordam em atribuir o polimorfismo a uma divisão fisiológica do trabalho entre organismos originalmente semelhantes. Isso equivale a supor que a diferenciação filogenética das castas surgiu na esfera da função antes de se manifestar em peculiaridades estruturais (Wheeler, 1910, p.118).
Desta suposição decorre que seria esperada pouca ou nenhuma especialização em espécies monomórficas primitivas (por exemplo, Ponerinae) , e uma acentuada divisão de funções em espécies dimórficas, ficando as espécies mais claramente polimórficas em um estágio intermediário: uma especialização menos rígida ou mais quantitativa do que qualitativa (Allee et al., 1965).
Em muitas formigas em que há subdivisões mais claras da casta das obreiras (por exemplo, soldados), há provavelmente divisão de trabalho mais clara (Michener & Michener, 1951, p.124).
Estas expectativas são até certo ponto confirmadas quando se examina a extensa literatura específica sobre a organização do trabalho em colônias de formigas - além das referências quase obrigatórias a esse tópico em tratados e revisões mais gerais (por exemplo, Wheeler, 1910; Goetsch, 1957; Michener & Michener, 1951; Morley, 1958; Sudd, 1970).
Em 1911, Buckingham cita diversos autores do século XIX que se ocuparam dessa questão, e realiza ela própria um extenso estudo observacional e experimental sobre as relações entre tamanho, idade e função em Camponotus americanus, Camponotus herculeanus pictus, Pheidole pilifera, Pheidole vinelandica e Pheidole dentata. Nos dois gêneros estudados, não foi identificada uma divisão estrita de tarefas, e sim uma participação predominante de certas classes de obreiras em certas atividades; em Camponotus, onde há uma série polimórfica contínua de obreiras, as menores e intermediárias ocupam-se da maioria das tarefas (cuidados com a ninhada, construção, forrageamento) e as maiores são menos ativas, destacando-se apenas na defesa. Em Pheidole, onde não há obreiras de tamanho intermediário, são desempenhadas pelas obreiras pequenas as tarefas que no outro gênero cabem às intermediárias e às menores.
O tema foi retomado em muitos estudos posteriores. Goetsch (1929/30) e Ehrhardt (1930/31), utilizando marcação de obreiras e intervenções experimentais, relatam divisão quantitativa de trabalho (predomínio de desempenho de algumas tarefas por certas obreiras) no gênero polimórfico Messor. Em Myrmica rubra, Ehrhardt (1930/31) relaciona a especialização em tarefas com a idade e com diferenças individuais. Ainda com o procedimento de marcação de obreiras, Ökland (1930/31) verificou constância dos mesmos indivíduos em tarefas externas ao ninho em Formica rufa. Várias espécies de Camponotus foram estudadas por Kiil (1934), Lee (1938/39) e Vowles (1953), com resultados gerais semelhantes aos de Buckingham (1911). Weir (1958) encontrou, em duas espécies de Myrmica, modificações na preferência por tarefas ao longo da idade, sendo as tarefas internas ao ninho características de obreiras mais jovens. Dobrzánska (1959) realizou com diversas espécies de Formica um dos poucos estudos que focalizaram tanto atividades externas ao ninho (coleta de secreções de afídios, forrageamento) como internas (cuidados com a ninhada), e que analisaram simultaneamente relações com tamanho, com idade e com características individuais como excitabilidade e motilidade. Verificou que obreiras de diferentes tamanhos e com características individuais diferentes diferem na proporção de desempenho das várias tarefas, e que obreiras jovens tendem a permanecer no ninho cuidando da ninhada.
Em praticamente todos esses estudos, os autores relatam uma divisão quantitativa, antes que qualitativa, de tarefas: as obreiras desempenham eventualmente todas as tarefas, mas exibem preferência por algumas. Mesmo no caso dos "soldados" de espécies dimórficas não há concordância sobre o desempenho exclusivo da função de defesa do ninho (Goetsch, 1957; Gregg, 1942). Comparações mais detalhadas entre os resultados dos vários estudos são dificultadas pelas diferenças de procedimentos de coleta e de análise. Diversos estudos focalizam apenas tarefas externas ao ninho (e.g. Ökland, 1930/31; Kiil, 1934). As variáveis focalizadas na análise - tamanho, idade, características individuais - diferem e raramente são consideradas em conjunto, o que deixa em aberto a questão da determinação da divisão de trabalho por fatores morfológicos, fisiológicos ou de experiência.
Apesar dessas restrições, alguns aspectos gerais podem ser destacados no quadro oferecido pela literatura. Uma relação estrita entre tamanho da obreira e tarefas desempenhadas não é encontrada. Verifica-se basicamente uma tendência ao desempenho proporcionalmente maior de algumas atividades pelas obreiras de tamanhos diferentes. Na maioria dos estudos, o gradiente de tamanhos, ainda que contínuo, é subdividido em três categorias. As obreiras mínimas são geralmente associadas a tarefas internas ao ninho, como cuidar da ninhada, da limpeza ou do fungo, nas espécies que o cultivam (Allee et al., 1965; Autuori, 1949, 1957; Mariconi, 1970; Michener & Michener, 1951; Goetsch, 1957; Bernard, 1968). "Soldados", as obreiras maiores, são observados em locomoção ou inatividade em áreas externas, sendo associados principalmente às tarefas de defesa do ninho (Allee et al. , 1965; Hollingsworth, 1960). Obreiras de tamanho intermediário, nas espécies em que ocorrem, são mencionadas como construtoras e coletoras de vegetal (Autuori, 1957) ou como "generalistas" que desempenham diversos tipos de tarefas (Michener & Michener, 1951). Supondo-se, com Wheeler (1910), que a diferenciação filogenética das castas tenha surgido na esfera da função antes de se manifestar em características estruturais, essa menor especialização das obreiras de tamanhos intermediários poderia estar relacionada à tendência evolucionariamente mais recente ao dimorfismo, com desaparecimento dos tamanhos intermediários. Essa interpretação é compatível com o fato de que se encontra alguma divisão de trabalho, sem relação claramente estabelecida com tamanho ou idade, mesmo em grupos monomórficos primitivos como Ponerinae ( Bonavita & Poveda, 1970; Haskins & Haskins, 1950). Este quadro sugere que seja interessante analisar a divisão de trabalho em espécies claramente polimórficas utilizando uma categorização mais detalhada de tamanho das obreiras intermediárias, que poderia oferecer pistas sobre a relação entre especialização de funções e evolução do dimorfismo.
Quanto à idade, constata-se consistentemente que obreiras jovens tendem a permanecer no ninho, desempenhando indiferentemente as várias tarefas internas. Neste caso, a divisão de trabalho consistiria apenas na diferença de preferência por locais (Sudd, 1970). As diferenças relacionadas com idade poderiam também refletir diferenças individuais decorrentes de experiência e de interação com outros fatores: por exemplo, Dobrzánska (1959) sustenta que as diferenças que encontrou entre obreiras de tamanhos diferentes não estariam ligadas diretamente ao tamanho, e sim a características individuais (motilidade, atividade, excitabilidade) originadas de histórias de desenvolvimento diferentes para obreiras de tamanhos diferentes. Uma interpretação semelhante é apresentada por Chen (1938), a respeito de participação diferencial de obreiras de Camponotus japonicus na construção do ninho. Outro efeito de experiência sugerido por vários autores decorreria de uma tendência à persistência na tarefa: realizar uma tarefa aumentaria a probabilidade de voltar a realizá-la, criando assim algum tipo de divisão de trabalho entre indivíduos (Goetsch, 1957; Morley, 1958; Vowles, 1955).
Esta revisão da literatura a respeito da organização do trabalho em colônias de formigas sugeriu, à época de sua realização2, que as questões em aberto a respeito desse tema justificavam sua retomada. Esse projeto pareceu especialmente oportuno considerando que, dada a história de nosso laboratório3, tínhamos acesso a uma espécie pouco pesquisada sob este aspecto até então. O gênero Atta, possivelmente devido à sua distribuição, tinha sido pouco focalizado pela literatura; as referências a respeito da divisão de trabalho nesse gênero, e na espécie sexdens em particular, não provinham de trabalhos específicos sobre o tema (Autuori, 1957; Weber, 1956).
Este estudo visou portanto explorar a questão da organização do trabalho na sociedade de saúvas. O planejamento da coleta de dados foi orientado pela hipótese de que a coordenação de atividades em uma colônia de insetos poderia ser obtida através de especialização comportamental dos indivíduos, e pelas suposições de que essa especialização, no caso de ser verificada, poderia relacionar-se a especialização estrutural, a especialização ontogenética ou a ambas. Dadas as indicações da literatura, considerou-se relevante investigar a organização do trabalho levando em conta tanto a natureza da tarefa quanto o seu local de realização.
Método
O procedimento de coleta de dados consistiu no registro das atividades desempenhadas pelas obreiras, que eram simultaneamente classificadas quanto ao tamanho e à idade.
Na espécie em questão - a formiga "agricultora" Atta sexdens rubropilosa Forel 1908 - a especialização estrutural não é muito acentuada: há polimorfismo de obreiras, mas não há grupos claramente definidos com características estruturais diferentes. O que se encontra é uma grande e contínua variação de tamanho das obreiras, que se poderia supor estar relacionada a especialização comportamental. Visando preservar ao máximo a proximidade a esse contínuo, foi desenvolvida uma escala de tamanhos com dez pontos ou classes de tamanho (Fig. 1), com os quais as obreiras eram comparadas a cada registro para a classificação (Carvalho, 1971).

A existência de diferenças comportamentais ao longo da ontogênese seria outro mecanismo possível de especialização comportamental, a exemplo do que se conhece a respeito de divisão de trabalho em abelhas melíferas (Lindauer, 1952). Conforme observações de vários autores (Wheeler, 1910; Morley, 1958; Dobrzánski, 1971; Carvalho, 1971), há relação entre a coloração do tegumento e a idade de obreiras de formiga, podendo a cor do tegumento ser utilizada como índice de idade. Em vista disto, a variação de cores das obreiras foi também arbitrariamente subdividida em cinco classes de cor, variando de amarelo a marrom escuro.4
As observações foram realizadas em 28 colônias artificiais, com idade de 6 a 32 meses (Fig. 2), obtidas segundo o procedimento desenvolvido por Cunha (1968). Os recipientes de vidro tinham 13 cm de altura e 9 ou 11 cm de diâmetro, com o fundo ajustado em placas removíveis de acrílico; eram mantidos sobre bandejas de alumínio de 40 por 30 cm , nas quais eram colocadas as folhas para o corte, e onde as obreiras formavam os depósitos de lixo. Foram obtidos 24793 registros, em 110 sessões de observação de duração variável. As observações nas áreas externas da colônia (bandeja e superfície) foram feitas a olho nu; nas áreas internas (câmara e canais) foi utilizado um estereomicroscópio.

INSTALAÇÃO DAS COLÔNIAS
As categorias utilizadas para a classificação das atividades combinaram três variáveis: (a) o local em que eram desempenhadas (fora do frasco, na superfície, nos canais ou na câmara); (b) o objeto da atividade (fungo, ninhada, lixo, vegetais, outra obreira etc); e (c) o tipo de atividade (lamber, transportar, explorar, cortar etc). A combinação dessas variáveis produziu 99 categorias diferentes (por exemplo, "lamber folha fora", "transportar ninhada na câmara" etc), que foram posteriormente sub-agrupadas no decorrer da análise.
A análise visou responder às seguintes questões:
a) que proporção de cada atividade é desempenhada pelas várias classes de obreiras (segundo tamanho e idade)
b) quais atividades são típicas de cada classe de obreiras
c) qual ou quais de duas variáveis consideradas na categorização das atividades - local e objeto - estão mais relacionadas à distribuição de atividades encontrada. A variável "tipo de atividade" foi descartada da análise após uma primeira inspeção dos resultados, que evidenciou ausência de regularidades passíveis de serem atribuídas a ela; as variáveis "local" e "objeto" foram tratadas separadamente.
Para as análises de distribuição de atividades entre as classes, os registros relativos a outras obreiras e à içá foram tratados em conjunto, devido ao pequeno número destes últimos
A incidência de cada classe de tamanho e de cor no total de registros apresentou-se bastante variável, como é indicado na Tabela 1. Desta forma, seria de esperar que, se não houvesse especialização comportamental relacionada às classes, a participação de cada classe em cada atividade fosse proporcional à sua incidência na população. A proporção de cada classe na população foi portanto considerada como expressão quantitativa da hipótese de inexistência de especialização, e a análise estatística utilizou esses valores para a comparação com as proporções efetivamente verificadas de participação de cada classe em cada atividade. O mesmo raciocínio foi aplicado à incidência das atividades na amostra (Tabela 2), que foi tomada como valor da hipótese nula na análise do ítem (b).

Resultados
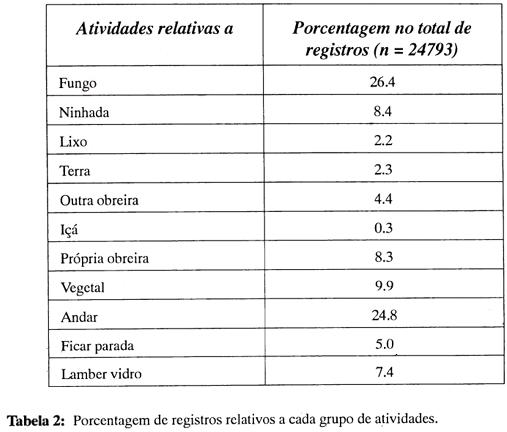
A Figura 3a, b apresenta os resultados da análise relativa à questão (a): que proporção de cada atividade, definida de acordo com seu objeto, é desempenhada pelas várias classes de obreiras, ou qual é a participação de cada classe no desempenho das atividades relativas a cada objeto? No eixo horizontal são representadas as classes de obreiras por tamanho (Fig. 3a) e idade (Fig. 3b), e no eixo vertical as diferenças entre as porcentagens observadas de desempenho de cada classe e as porcentagens representadas pelas classes na amostra (expressão da hipótese nula de inexistência de especialização). As diferenças assinaladas são estatisticamente significativas a 5%, pela Tábua de Diferenças entre Porcentagens (Geigy, 1965). Diferenças positivas significativas representam uma participação elevada da classe em questão na atividade considerada; diferenças negativas significativas representam uma participação muito reduzida; e diferenças não-significativas correspondem à hipótese nula (ausência de especialização).
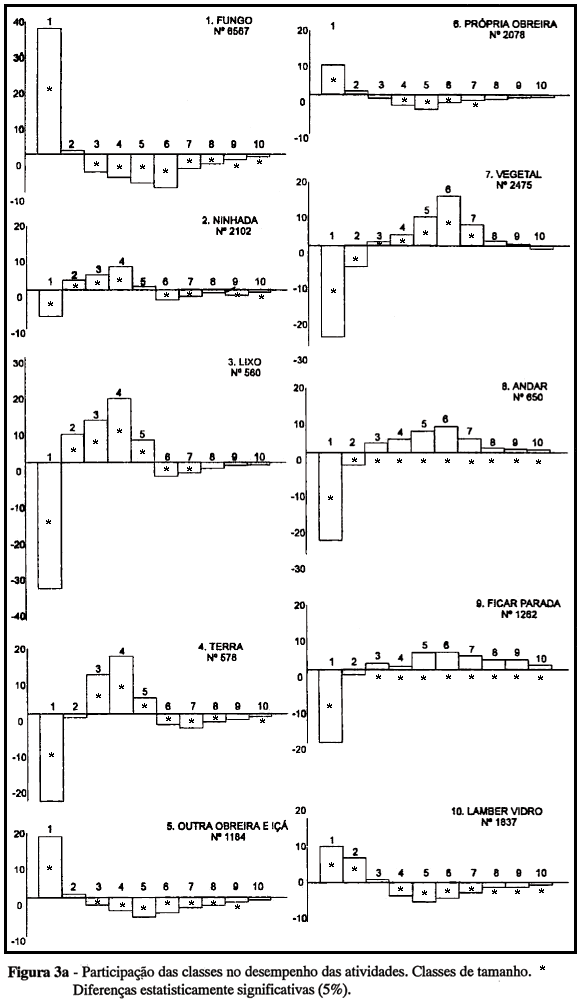
Na Fig. 3a verificam-se duas condições de rejeição da hipótese nula com relação a diferenças estruturais: a participação das classes de tamanho difere de um objeto de atividade para outro e, em cada caso, difere também dos valores esperados pelo acaso. A comparação entre os resultados relativos aos vários objetos de atividade permite a identificação de diferentes tipos de distribuição. Na primeira, há alta participação das obreiras menores (classe 1) e participação baixa ou não-significativa das demais classes (gráficos 1, 5, 6 e 10, referentes a Fungo, Outra obreira/içá, Própria obreira e Lamber vidro). No segundo tipo, há participação elevada de obreiras um pouco maiores (classes 2, 3 e 4) nas atividades relativas à Ninhada, Lixo e Terra (gráficos 2, 3 e 4). No terceiro tipo, onde se encontram as atividades relativas a Vegetal, a participação maior é das classes 5, 6 e 7 (gráfico 7). Nas categorias Andar e Ficar parada todas as classes, exceto 1 e 2, apresentam tendência a participação alta. A categoria Lamber vidro, decorrente do tipo de instalação utilizado para as colônias, apresenta uma distribuição semelhante à das atividades relativas a fungo e a outras obreiras/içá, nos dois casos atividades internas.
As diferenças relativas a classes de idade são menos definidas (Fig. 3b). Agrupando-se as classes mais jovens (A e B) e as mais velhas (C,D), as diferenças parecem estar mais relacionadas ao local da atividade do que ao seu objeto: as atividades realizadas no interior no ninho (fungo, ninhada, outras obreiras, a própria obreira e lamber vidro) apresentam participação alta das obreiras mais jovens, enquanto as classes mais velhas são predominantes em atividades externas (lixo, terra). Atividades que são realizadas tanto dentro quanto fora do ninho (relativas a vegetal, andar e imobilidade) não apresentam uma distribuição clara em relação à idade.
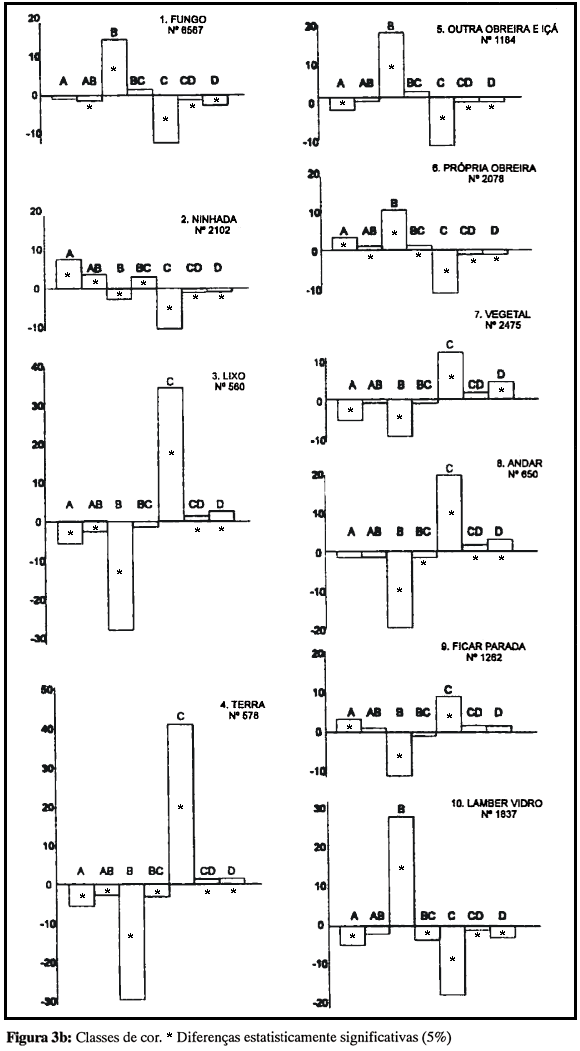
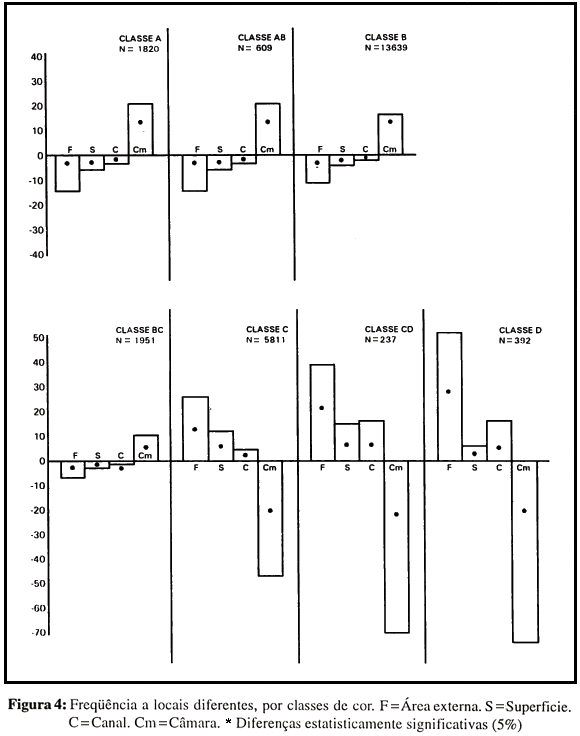
Este resultado sugere que o mecanismo de distribuição das atividades, no caso da variável idade, esteja mais relacionado ao local do que ao objeto; esta possibilidade harmoniza-se com outros dados da literatura, que apontam a tendência de obreiras jovens a permanecerem no interior do ninho. De fato, quando é analisada a freqüência das várias classes de idade a quatro locais da colônia - câmara, canais, superfície e área externa - identificam-se duas tendências bem nítidas (Fig. 4): freqüentar a câmara e freqüentar outros locais. Todas as diferenças neste caso são significativas, e a segunda tendência substitui a primeira em um contínuo perfeito: até a classe C a primeira tendência decresce regularmente ao mesmo tempo que cresce a segunda; a partir da classe C, as duas tendências, já invertidas, diferenciam-se cada vez mais. Este resultado indica que a mudança de comportamento com a idade é controlada pelo local mais do que pela natureza da atividade; ou seja, a mudança ontogenética não se caracterizaria pelo surgimento de novos comportamentos, ou de reatividade a novos objetos de atividade, mas sim pelo acesso e freqüência a novos locais da colônia.
FREQÜÊNCIA DE OBREIRAS AOS LOCAIS DA COLÔNIA, POR CLASSES DE IDADE
Para explorar um pouco mais essa possibilidade, foram comparadas as distribuições das mesmas atividades quando realizadas em locais diferentes. A figura 5 (a,b,c) apresenta três exemplos dessas comparações, respectivamente para "Imobilidade", "Andar" e "Transportar vegetal". Nos dois primeiros casos, as distribuições na câmara e fora do ninho aparecem claramente invertidas; no caso de transporte de vegetais também se verifica essa tendência, mas nota-se que as obreiras mais jovens (classe A) participam pouco dessa atividade, mesmo quando realizada na câmara, o que indica alguma diferenciação controlada pelo objeto da atividade, o que não se observa nas demais classes de idade. A ausência de diferenciação atribuível ao tipo de atividade é ilustrada pelo fato de que obreiras da classe A apresentam comportamentos de transporte e de lambimento em relação à ninhada, mas não em relação a fungo e vegetais. Desta forma, a mudança ontogenética também não implica o surgimento de novos padrões de comportamento, e sim a extensão de um padrão já existente a um objeto novo.
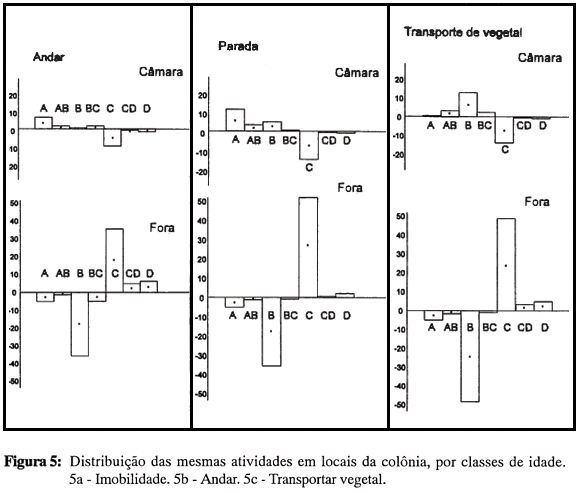
No conjunto, estes resultados indicam que, pelo menos a partir de uma certa idade (classe B), é inadequado falar em especialização ontogenética em relação às atividades, já que as distribuições refletem principalmente uma propensão diferencial de freqüentar os vários locais da colônia.
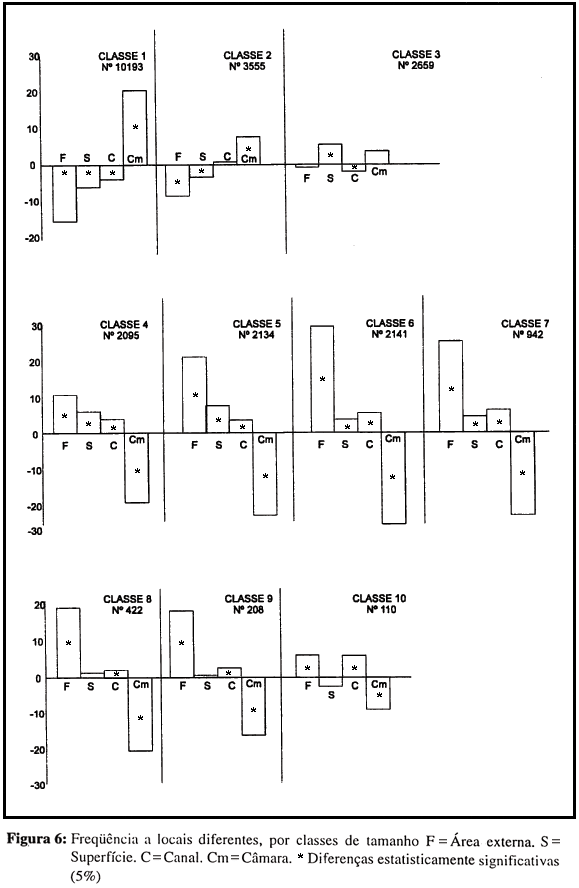
A Figura 6 apresenta a freqüência diferencial dos locais por obreiras de tamanhos diferentes. Podem ser identificadas as mesmas tendências verificadas quanto à idade (permanecer na câmara ou freqüentar outros locais), mas de forma muito mais dispersa e gradativa: na classe 1 prevalece a primeira tendência; nas classes 2 e 3 parece ocorrer uma transição; da classe 4 à 6 prevalece a segunda tendência, que se dispersa gradualmente nas classes maiores. Quando se comparam distribuições relativas aos mesmos objetos de atividade, mas em diferentes locais, verifica-se um ligeiro deslocamento da distribuição no sentido de maior participação das classes menores em atividades internas (é o caso de atividades relativas a vegetais); por outro lado, em atividades relativas a lixo, que são predominantemente externas, há participação alta das classes 2 e 3, que se ocupam pouco de atividades externas relativas a outros objetos: é como se estas obreiras pequenas saíssem do ninho "para trabalhar com o lixo", e não o inverso. O papel do local da atividade na distribuição mostra-se portanto muito mais reduzido no caso de diferenças estruturais do que no de diferenças ontogenéticas.
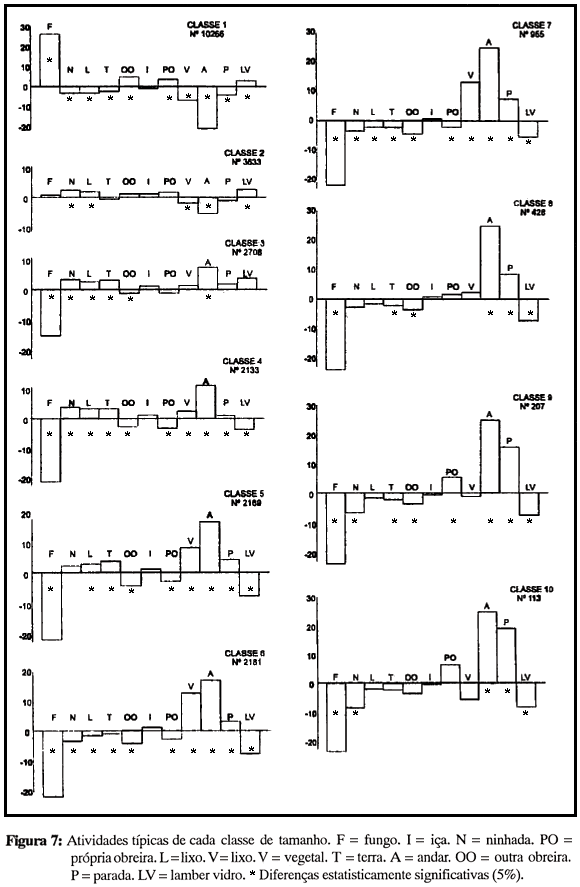
Dada a influência aparentemente reduzida do tipo de atividades sobre a sua distribuição em termos de idade das obreiras, a questão seguinte - quais são as atividades típicas de cada classe - foi focalizada apenas em termos de diferenças estruturais. Para esta análise, a porcentagem de cada classe de tamanho que se ocupou de cada atividade (agrupadas por objeto às quais eram dirigidas) foi comparada com a porcentagem representada pelas atividades no conjunto do registro. Na Figura 7 observa-se que as mudanças ocorrem gradualmente ao longo do contínuo de tamanhos: a distribuição típica de cada classe é semelhante à das classes vizinhas, o que significa que a divisão de atividades não é qualitativa, e sim quantitativa: alguns grupos de obreiras são predominantes, mas não exclusivos, no desempenho das atividades relativas a cada objeto.
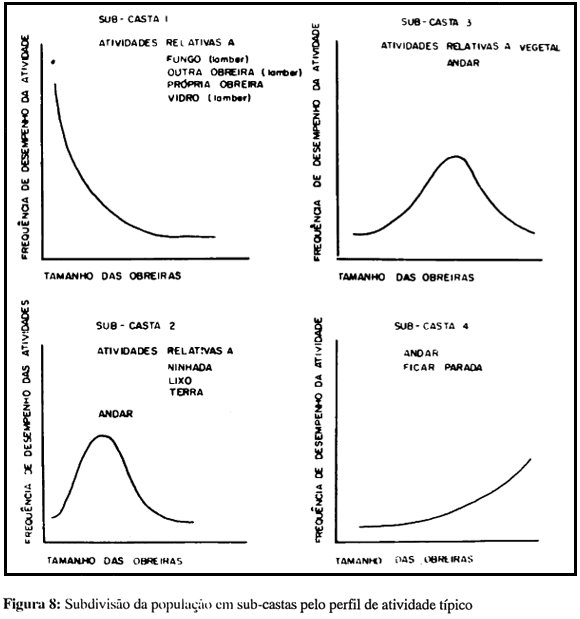
No conjunto, no entanto, as distribuições sugerem uma distribuição da população de obreiras, com base em critérios comportamentais - atividades predominantes - não em dez, mas em quatro subgrupos ou subcastas, ilustrados na figura 8.
A análise seguinte retomou a questão do papel relativo do local e do objeto da atividade com um tratamento experimental. Foram feitas novas observações sobre dois tipos de atividade - corte e transporte de vegetais - em colônias com dois tipos de alterações na organização espacial. No primeiro tipo, a colônia foi alojada em uma caixa de acrílico completamente fechada, com três compartimentos internos (Fig. 9): no maior deles foi colocada a esponja de fungo; o segundo compartimento, menor, era periodicamente abastecido com folhas; e o terceiro foi mantido vazio, com a expectativa de que fosse utilizado para o depósito de lixo, o que realmente ocorreu. No segundo tipo, o arranjo era idêntico, com a exceção de que a caixa permanecia aberta, permitindo às obreiras o acesso ao exterior; neste caso, o depósito de lixo foi formado fora da caixa . Esperava-se que o registro de atividades que são tipicamente desempenhadas fora da colônia, quando realizado nessas condições alteradas, oferecesse pistas sobre o papel do local na determinação da distribuição de atividades entre as classes de tamanho.

Não se verificaram diferenças nas distribuições relativas a corte e transporte de vegetais nas duas condições, que foram por isso comparadas em conjunto com os dados anteriormente disponíveis. Para esta análise, os dados das classes de obreiras que tinham participação relativa baixa, alta e não-significativa nos primeiros registros foram agrupados e comparados com os dados das classes correspondentes no segundo registro. As diferenças não alcançaram significância em nenhuma das duas atividades analisadas, indicando alguma rigidez na distribuição, pelo menos no que se refere às alterações espaciais examinadas.
Discussão
Os resultados principais deste estudo confirmam e estendem para a espécie Atta sexdens os achados sobre organização do trabalho na colônia e suas relações com diferenças estruturais (tamanho) e idade em outras espécies polimórficas5. A distribuição de atividades entre as classes de tamanho apresentou-se com natureza quantitativa antes que qualitativa, e relacionada principalmente ao objeto de atividade. Quanto à idade, a distribuição é mediada pela preferência por locais de trabalho - dentro ou fora da colônia.
Algumas questões podem ser colocadas a partir desses resultados. Em primeiro lugar, o fato de se verificar uma divisão de atividades, ainda que quantitativa, entre obreiras com características morfológicas diferentes, permite perguntar sobre a extensão das especializações relacionadas ao polimorfismo, e sobre as relações funcionais entre essas especializações e a natureza das tarefas preferencialmente desempenhadas. O polimorfismo não se reduz a uma diferença linear de tamanho corporal (aumento proporcional de todas as partes do corpo), mas envolve também outros aspectos propriamente morfológicos. Hollingsworth (1960) analisou detalhadamente diferenças estruturais em sub-castas de obreiras de Anomma, mostrando que a mudança de tamanho em uma série polimórfica contínua é acompanhada por mudanças na forma, particularmente notáveis na cabeça: a cabeça da obreira maior é proporcionalmente mais alta e mais larga do que a da obreira menor; as antenas das obreiras maiores apresentam maior número de segmentos no funículo, e os segmentos são mais compridos do que largos; nas obreiras menores pode haver inversão (largura maior do que o comprimento) e o número de segmentos é menor. Nos indivíduos menores o dente basal da mandíbula é quase indistinto, enquanto se apresenta bem nítido nas demais; por outro lado, quanto menor a obreira, maior o número de dentes (a obreira maior apresenta dois). É razoável supor que diferenças desse tipo estivessem correlacionadas com as tarefas desempenhadas pelas obreiras, ou pelo menos com a eficiência com que as desempenham. De fato, embora todas as obreiras fossem observadas defendendo o ninho, sua eficiência decrescia com o tamanho, e "quanto menor a mandíbula, mais eficiente parece ser como instrumento de corte" (Hollingsworth, 1960, p.22).
Bernard (1968) oferece outra informação interessante para este ângulo de análise: em Camponotus e em Pheidole, há diferenças na morfologia cerebral de obreiras de tamanhos diferentes, as obreiras menores apresentando corpos pedunculados duas a três vezes maiores do que as obreiras máximas; e em Lasius as obreiras têm corpos pedunculados maiores e mais complexos do que os da rainha, que no entanto tem um volume cefálico doze vezes maior.
Não temos conhecimento de estudos morfológicos desse tipo em Atta sexdens, mas algumas observações e mensurações realizadas no processo de elaboração da escala de tamanhos são sugestivas: medidas da cabeça (comprimento e largura, comprimento da mandíbula, escapo, funículo) tendem a ser proporcionalmente maiores em relação ao comprimento total da obreira na classe 10 em comparação com a classe 1, com exceção da largura do funículo, onde essa proporção se inverte; a diferença mais notável de proporção é entre o comprimento da pata dianteira e o comprimento total (quase 80% nas obreiras maiores contra pouco menos de 50% nas mínimas). Considerando as especificidades e requisitos das várias tarefas envolvidas na sobrevivência de formigas altamente especializadas como as agricultoras - corte, transporte e tratamento dos vegetais, poda, transplante, umedecimento e proteção do fungo contra parasitas, remoção de resíduos etc - pode-se imaginar que a especialização comportamental fosse beneficiada por uma especialização estrutural, ou mesmo que a exigisse. Há um aspecto geral de coerência entre as tarefas e as características físicas que acompanham a diferenciação de tamanhos: por exemplo, parece razoável que as tarefas relativas ao fungo sejam desempenhadas por obreiras pequenas, que poderiam com mais facilidade do que as grandes penetrar nas locas formadas pela esponja; é possível que a estrutura diferenciada das antenas esteja relacionada a capacidades sensoriais importantes para certas tarefas; e ainda, é difícil imaginar uma obreira da classe 1 cortando talos de vegetal cuja espessura mal cabe entre suas mandíbulas, ou transportando pedaços de vegetal de grandes dimensões. Lutz (1929) encontrou relação entre o peso da obreira e o peso da carga em Atta cephalotes: obreiras pequenas não podiam transportar cargas muito grandes. Essas possíveis relações fariam esperar algum grau de rigidez no engajamento das classes de tamanho em tarefas para as quais estariam estruturalmente especializadas, como sugere Wilson (1983).
Esta suposição, no entanto, não se harmoniza bem com a constatação de que a divisão de trabalho é quantitativa, ou seja, de que obreiras de diversos tamanhos podem se engajar na mesma atividade, ainda que com freqüências diferenciais.
Duas direções de pesquisa podem ser sugeridas a partir dessas colocações: a análise de diferenças estruturais entre as sub-castas de obreiras e de suas relações funcionais com os vários tipos de atividades - que poderia ser guiada pela sub-divisão da população de obreiras em quatro sub-castas, sugerida neste trabalho; e a investigação sobre a flexibilidade da divisão de trabalho em condições alteradas de composição da população da colônia em termos de classes de tamanho6.
A sub-divisão da população de obreiras em quatro sub-castas é outra indicação que parece ser relevante comentar. Como foi visto na Introdução deste artigo, as obreiras de tamanho intermediário são freqüentemente consideradas como "generalistas" ou não-especializadas, enquanto nossos resultados sugeriram algum grau de especialização: classes 2, 3 e 4 em atividades relativas a ninhada, lixo e terra, e classes 5,6 e 7 em atividades relativas a vegetal. Caso as classes de tamanho intermediário tivessem sido agrupadas em uma única categoria ("médias", em oposição a mínimas - classe 1, e a máximas - classes 8-10), essas diferenças não se revelariam. Uma outra diferença que é interessante apontar refere-se ao engajamento de obreiras da classe 2 em atividades relativas ao lixo, que eram realizadas na área externa do ninho devido às condições peculiares de instalação de nossas colônias. Essa situação diverge da que se verifica na natureza, onde o lixo é interno, situado em panelas especiais (Autuori, 1942, 1947). Uma vez que a literatura indica que obreiras pequenas tendem a permanecer no interior do ninho, e já que as atividades relativas a lixo foram as únicas atividades externas em que se observou engajamento significativo da classe 2, esta diferença configura uma espécie de situação experimental que complementa a que foi ensaiada com as colônias fechadas e semi-fechadas neste trabalho, com o mesmo tipo de resultado: as obreiras pequenas desempenham as tarefas relativas ao lixo, ainda que sejam externas, sugerindo que os fatores de controle relacionem-se mais com a atividade do que com o local. Esta indicação não se confirma no caso das classes de cor: são as obreiras de classes C e mais velhas que desempenham as atividades relativas a lixo em nossas colônias.
Esta observação introduz uma outra questão de interesse que não foi contemplada neste trabalho devido ao número insuficiente de registros: a interação cor-tamanho. Rodrigues (1995) observa que quando a obreira de classe C pertence à classe 1 de tamanho, algum tipo de interação faz dessa obreira uma excelente jardineira (cuidadora de fungo, atividade privilegiada da classe 1).
Finalmente, fica em aberto a partir de nossos resultados a investigação de mudanças de comportamento com a idade. Pesquisas psicofisiológicas e psicofarmacológicas talvez possam investigar a relação entre a tendência de obreiras jovens a permanecerem no interior do ninho e fatores relacionados a desenvolvimento sensorial e motivacional (por exemplo, "timidez", Sudd, 1970). No mesmo contexto, outro aspecto sugestivo é a distribuição de tarefas internas entre obreiras de idade diferente: obreiras mais jovens (classes A e AB) só apresentaram participação significativa em atividades relativas à ninhada, a si próprias, e em inatividade; embora apresentem os comportamentos de lamber e transportar a ninhada, não apresentam com a mesma freqüência esses comportamentos em relação a fungo. O fato de um padrão motor já presente manifestar-se seletivamente em relação a seu objeto novamente sugere variáveis sensoriais e motivacionais a serem investigadas.
Embora já venham sendo estudadas há mais de um século, as sociedades de insetos continuam a apresentar desafios estimulantes para a compreensão de suas especificidades, da flexibilidade e das características definidoras dos fenômenos sociais na natureza.
REFERÊNCIAS BIBLIOGRÁFICAS
ALLEE,W.C.; PARK, O.; EMERSON, A.E.; PARK, T.; SCHMIDT, K.P. Principles of animal ecology. London, W.B. Saunders, 1965. [ Links ]
AUTUORI, M. Contribuição para o conhecimento da saúva. III: escavação de um sauveiro. Arquivos do Instituto Biológico, v.13, p.137-48, 1942. [ Links ]
AUTUORI, M. Contribuição para o conhecimento da saúva. IV: o sauveiro depois da primeira revoada. Arquivos do Instituto Biológico, v.18, p.39-70, 1947. [ Links ]
AUTUORI, M. Investigação sobre a biologia da saúva. Ciência e Cultura, v.1, n.1/2, p.4-15, 1949. [ Links ]
AUTUORI, M. A saúva e seu combate. Anhembi, v.25, n.74, p.3-20, 1957. [ Links ]
BERNARD, F. Les fourmis ( Hymenoptera-Formicidae) d'Europe Occidentale et Septentrionale. Paris, Masson, 1968. [ Links ]
BONAVITA, A.; POVEDA, A. Mise en évidence d'une division de travail chez une fourmi primitive. Comptes Rendus des Séances de l'Academie des Sciences, v.270, p.515-8, 1970. [ Links ]
BUCKINGHAM, E. N. Division of labour among ants. Proceedings of the American Academy of Arts and Sciences, v.46, p.426-507, 1911. [ Links ]
CARVALHO, A.M.A. Alguns dados sobre a divisão de trabalho entre obreiras de Atta sexdens rubropilosa Forel, 1908 (Hymenoptera - Formicidae), em colônias iniciais, mantidas em laboratório. São Paulo, 1972. 175p. Tese (Doutorado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
CARVALHO, A.M.A. Desenvolvimento de uma técnica para o estudo de aspectos da organização social em colônias de formigas. São Paulo, 1971. 96p. Dissertação (Mestrado) - Instituto de Psicologia, Universidade de São Paulo. [ Links ]
CHEN, S.C. Social modification of the activity of ants in nest-building. Physiological Zoology, v.10, p.420-37, 1938. [ Links ]
CUNHA, W.H.A. Algumas observações sobre o comportamento da fêmea de Atta sexdens rubropilosa Forel, 1908, durante a fundação da colônia. Ciência e Cultura, v.20, n.2, p.233-4, 1968. [ Links ]
DOBRZÁNSKA, J. Studies on the division of labour in ants of the genus Formica. Acta Biologiae Experimentalis, v.19, p.57-81, 1959. [ Links ]
DOBRZÁNSKI, J. Manipulatory learning in ants. Acta Neurobiologiae Experimentalis, v.31, p.111-40, 1971. [ Links ]
EHRHARDT, S. Über arbeitsteilung bei Myrmica- und Messor Arten. Zeitschrift fur Morphologie und Ökologie der Tiere, v.20, p.755-812, 1930/31. [ Links ]
GEIGY, J.R., ed. Tablas científicas. Brasilea, J.R.Geigy, 1965. [ Links ]
GOETSCH, W. The ants. Chicago, Chicago University Press, 1957. [ Links ]
GOETSCH, W. Beiträge zur Biologie körnersammelnder Ameisen. Zeitschrift fur Morphologie und Ökologie der Tiere, v.16, p.371-452, 1929/30. [ Links ]
GREGG, R.E. The origin of castes in ants with special reference to Pheidole morrisi Forel. Ecology, v.23, p.295-307, 1942. [ Links ]
HASKINS, C.P.; HASKINS, E.F. Notes on the biology and social behaviour of the archaic Ponerine ants of the genera Myrmecia and Promyrmecia. Annals Entomological Society of Amererica, v.43, p.461-91, 1950. [ Links ]
HOLLINGSWORTH, M.J. Studies on the polymorphic workers of the Army ant Dorillus (Anomma) nigricans Illiger. Insectes Sociaux, v.7, p.17-37, 1960. [ Links ]
KIIL, V. Untersuchungen über Arbeitsteilung bei Ameisen (Formica rufa L., Camponotus herculeanus L. und C. ligniperda Latr. ). Biologisches Zentralblatt, v.54, p.114-56, 1934. [ Links ]
LEE, J. Division of labor among the workers of the Asiatic Carpenter Ants (Camponotus japonicus var. atterrimus). Peking Natural History Bulletin, v.13, p.137-45, 1938/39. [ Links ]
LINDAUER, M. Ein Beitrag zur Frage der Arbeitsteilung im Bienenstaat. Zeitschrift fur Vergleichende Physiologie , v.34, p.299-345, 1952. [ Links ]
LUTZ, F.E. Observations on leaf-cutting ants. American Museum Novitates, v.388, p.1-21, 1929. [ Links ]
MARICONI, F.A.M. As saúvas. São Paulo, Ceres, 1970. [ Links ]
MICHENER, C.D.; MICHENER, M.H. American social insects. New York, Van Nostrand, 1951. [ Links ]
MORLEY, D.W. A evolução de uma sociedade de insetos. São Paulo, Nacional, 1958. [ Links ]
ÖKLAND, F. Studien über die Arbeitsteilung und die Teilung des Arbeitsgebietes bei der roten Waldameise (Formica rufa L.). Zeitschrift fur Morphologie und Ökologie der Tiere, v.20, p.63-131, 1930/31. [ Links ]
RODRIGUES, M.M.P Divisão de trabalho e especialização em obreiras de Atta sexdens. Psicologia USP, v.6, n.1, 1995. [ Links ]
SUDD, J.H. An introduction to the behaviour of ants. London, Arnold, 1970. [ Links ]
VOWLES, D.M. The foraging of ants. British Journal of Animal Behaviour, v.3, n.1, p.1-13, 1955 [ Links ]
VOWLES, D.M. Individual behaviour patterns in ants. Advancement of Science, v.10, p.18-21, 1953. [ Links ]
WEBER, N.A. Evolution in fungus-growing ants. Proceedings of the 18th International Congress of Entomology, v.2, p.459-74, 1956. [ Links ]
WEIR, J.S. Polyethism in workers of the ant Myrmica. Insectes Sociaux, v.5, p.97-128, 1958. [ Links ]
WHEELER, W.N. The ants: their structure, development and behavior. New York, Columbia University Press, 1910. [ Links ]
WILSON, E.O. Caste and division of labour in leaf-cutter ants (Hymenoptera-Formicidae: Atta). I. The overall pattern in Atta sexdens. Behavioral Ecology and Sociobiology, v.7, p.157-65, 1980. [ Links ]
WILSON, E.O. Caste and division of labour in leaf-cutter ants (Hymenoptera- Formicidae: Atta). III. Ergonomic resiliency in foraging by A. cephalotes. Behavioral Ecology and Sociobiology, v.14, p.47-54, 1983. [ Links ]
WILSON, E.O. The origin and evolution of polymorphism in ants. Quarterly Review of Biology, v.28, p.136-56, 1953. [ Links ]
1 Parte de tese de doutorado em Psicologia - IPUSP, 1973. Orientador: Walter Hugo de Andrade Cunha.
2 A revisão apresentada aqui é uma síntese do capítulo introdutório da tese de Doutorado da autora (1972), no qual os trabalhos citados são comentados com maiores detalhes. Desenvolvimentos posteriores do tema são focalizados na discussão deste artigo.
3 V. Apresentação desta revista.
4 Detalhes sobre a elaboração e teste das escalas encontram-se em Carvalho (1971) e Carvalho (1972). A escala de cores foi posteriormente padronizada através de avaliação por 18 juízes, estabelecendo-se sua correspondência com cores do Munsell Book of Color - Neighbouring Hues Edition, a saber: A = 5YR 5/8; B = 2.5YR 4/8; C = 2.5YR 4/6; D = 10R 3/4; E = 2.5YR 3/4. Após a tabulação dos dados, as classes mais velhas (D e E) foram tratadas em conjunto devido ao número reduzido de registros. Foram tabulados como intermediários os casos que envolviam dificuldade de classificação entre dois pontos vizinhos da escala, resultado uma escala de 7 pontos: A, AB, B, BC, C, CD, D.
5 Resultados semelhantes para a espécie Atta sexdens foram obtidos por Wilson (1980) em estudo publicado alguns anos após a conclusão da tese em que se baseia este trabalho.
6 A segunda direção foi explorada por M.M.P.Rodrigues em tese de Doutorado e no artigo apresentado neste mesmo número.
